人參中苯甲酸脅迫響應基因WRKY7的克隆與表達分析
2017-04-05 13:25:14王梓李勇王蓉丁萬隆
中國中藥雜志 2017年6期
王梓+李勇+王蓉+丁萬隆

[摘要]WRKY轉錄因子是高等植物中最重要的轉錄因子基因家族之一,其在植物形態建成、生長發育以及應對生物(病菌、害蟲等)、非生物(干旱、鹽、冷、熱等)脅迫過程中發揮重要作用。該研究根據人參化感自毒物質苯甲酸誘導的人參轉錄組數據,設計全長擴增引物,利用RTPCR方法獲得1條編碼WRKY轉錄因子的基因,暫命名為WRKY7,并對其進行TA克隆、測序和序列分析。該基因cDNA全長1 216 bp,其開放閱讀框(open reading frame, ORF)為1 014 bp,編碼337個氨基酸。序列分析及系統進化分析表明,該蛋白屬于WRKY家族第Ⅲ類成員,與西洋參種的WRKY6轉錄因子親緣關系最近,同源性為87%。實時熒光定量PCR分析發現,苯甲酸脅迫后WRKY7的表達量顯著升高,推測該基因可能參與了人參響應苯甲酸脅迫的過程,為進一步研究人參自毒脅迫響應的分子機制奠定了基礎。
[關鍵詞]人參; WRKY轉錄因子; 化感自毒作用; 克隆; 序列分析
[Abstract]WRKY transcription factor is one of the most important transcription factor families widely existing in higher plants, which playing critical role in plant morphogenesis, development, biotic (including phytopathogens, pests etc.) and abiotic (drought, salt, chilling, high temperature, etc.) stress. In the present work, primers used to amplify fulllength gene encoding WRKY transcription factor were designed based on the transcriptome data of P. ginseng that induced by benzoic acid, one of the most important autotoxins identified from root exudates and rhizosphere soil of P. ginseng. Then, a WRKY gene, temporarily named as WRKY7, was confirmed by RTRCR. Furthermore, sequencing and sequence analysis of WRKY7 was conducted. Results indicated that, the full length cDNA of WRKY7 was 1 216 bp, the open reading frame (ORF) of which was 1 014 bp, encodes 337 amino acids. Homologous analysis and phylogenetic tree showed that, WRKY7 belonged to the Ⅲ category of WRKY families, which showing 87% similarity to WRKY6 in P. quinquefolius. Realtime PCR results showed that the expression of WRKY7 in P. ginseng induced by benzoic acid was upregulated markedly than the control, so we speculated that WRKY7 was involved in the response to benzoic acid stress, which will be helpful for further research on the molecular mechanism of ginseng plant response to benzoic acid stress.
[Key words]Panax ginseng; WRKY transcription factors; autotoxicity; cloning; bioinformatic analysis
轉錄因子是一類含有DNA結合域的蛋白質,能識別基因上游特異的核苷酸序列,形成轉錄復合體從而激活或抑制靶基因的表達,WRKY是植物十大轉錄因子家族之一。這類轉錄因子最早發現于甘薯[1],后相繼從20余種植物中分離出WRKY轉錄因子[2]。WRKY蛋白的共同特點是含有60個氨基酸組成的WRKY結構域,在N端有一段高度保守的WRKYGQK多肽序列,其是WRKY結構域的核心,如果上述氨基酸發生變異,將直接影響WRKY轉錄因子的DNA結合活性[3]。根據WRKY轉錄因子結構域的數量和鋅指結構,可將WRKY轉錄因子分為3類[4]:第Ⅰ類含有2個WRKY結構域,其鋅指結構為C2H2型;第Ⅱ類含有1個WRKY結構域,其鋅指結構為C2H2型;第Ⅲ類含有1個WRKY結構域,其鋅指結構為C2HC型。
WRKY轉錄因子與植物生長發育、衰老及環境脅迫響應密切相關。當植物遭受生物脅迫時,WRKY蛋白通過多通路、多層次調控基因表達,進而調節植物對脅迫信號的轉導途徑[5]。例如:AtWRKY33超表達可提高擬南芥對灰霉病和黑斑病的抗病性[6]。干旱、高溫、物理損傷等非生物脅迫同樣可以激發WRKY基因高表達,以幫助植物抵御外界不良環境[7]。例如:擬南芥AtWRKY18,AtWRKY33,AtWRKY40,AtWRKY46表達量增加能提高植株耐熱性[8];大豆GmWRKY21過量表達可增強擬南芥的抗寒能力[9]。WRKY轉錄因子在植物逆境信號傳遞過程中發揮重要作用,1個WRKY基因往往能響應多種脅迫信號,而多個WRKY基因之間則形成復雜的基因調控網絡。
人參是五加科多年生藥用植物,連作障礙嚴重影響人參產量和品質。已有研究表明,苯甲酸對人參具有明顯的自毒活性[10]。本課題組前期開展了苯甲酸脅迫下的人參轉錄組分析,發現了多個與苯甲酸脅迫相關的轉錄因子基因、過氧化物酶基因、活性氧及人參皂苷合成相關基因。本文在已有研究基礎上,利用RTPCR技術克隆獲得1條WRKY轉錄因子基因,暫命名為WRKY7。進而比較了該基因與其他WRKY家族基因的序列結構差異,通過系統進化關系分析了其與已知WRKY蛋白的關系。最終,通過Realtime PCR方法檢測了WRKY7在苯甲酸脅迫條件下的表達情況。本文為深入研究轉錄因子WRKY7的生物學功能奠定了理論基礎。
1材料與方法
11植物
取三年生人參幼苗,參考李勇等[11]的配方,用營養液和石英砂培養人參幼苗直至幼苗完全展開。人參苗脅迫處理參考Wu等[12]。分別于處理后第0,1,3,5,7,9天采集對照組和處理組人參根部樣品,每次取4株人參幼苗的毛根,混合保存于液氮中。
12試劑
苯甲酸(分析純,北京化學試劑有限公司);RNA Trizol提取試劑盒(Invitrogen公司);Dnase Ⅰ(RNaseFree)酶、RTase MMLV(RNase H)逆轉錄酶、Premix Taq酶、2×SYBR Premix Ex Taq酶、pMD18T載體均來自TaKaRa公司;大腸桿菌(Escherichia coli)DH5α感受態細胞(TIANGEN公司);DNA凝膠回收試劑盒、質粒DNA小量提取試劑盒為Axygen公司產品;瓊脂糖(Biowest公司);本實驗所用引物均由Sunbio公司合成。
13儀器
3K30臺式高速冷凍離心機(Sigma);紫外可見分光光度計(Nanodrop2000);基因擴增儀(BIORAD);熒光定量PCR儀(BIORAD);凝膠成像儀(BIORAD);DYY6C電泳儀(北京市六一儀器廠);臺式微量冷凍離心機(Beckman);移液器(Drangonlab);高壓滅菌鍋(華泰醫療設備有限公司);電熱恒溫培養箱(泰斯特儀器有限公司);超凈工作臺(亞泰科隆儀器技術有限公司)。
14RNA提取和cDNA合成
取人參根部樣品在液氮中研磨,用Trizol法提取人參根部總RNA,用Dnase I(RNaseFree)對產物進行處理,去除RNA中的DNA,并對其進行濃度測定。取上述提取的RNA模板1 μg,加入Random primer 1 μL,再加DEPC水至6 μL,將上述體系混勻后70 ℃孵育10 min,迅速冰浴2 min,加入5×RTase MMLV buffer 2 μL,dNTP(10 mmol·L-1)05 μL,RNase inhibitor 05 μL,RTase MMLV(RNase H)逆轉錄酶1 μL,25 ℃反應5 min,42 ℃反應60 min,70 ℃保溫15 min,即可得到cDNA,-20 ℃保存備用。
15WRKY7基因的擴增和克隆
根據RNAseq拼接數據返回的序列,利用Primer 3(http://primer3utee/)設計引物(表1),擴增基因的完整編碼區。以人參cDNA為模板,按照如下體系擴增人參WRKY7基因:cDNA模板1 μL,Premix Taq酶125 μL,10 μmol·L1上下游引物各1 μL,ddH2O 95 μL,終體積為25 μL。擴增程序:95 ℃預變性3 min;94 ℃變性30 s,56 ℃退火30 s,72 ℃延伸90 s,30個循環;最后72 ℃延伸5 min,4 ℃保溫。PCR產物用1%瓊脂糖凝膠電泳檢測,Axygen凝膠回收試劑盒用于目的片段的回收。將回收產物5 μL,Ligation solution Ⅰ 5 μL,pMD18T載體05 μL混勻置于4 ℃連接過夜。連接產物轉化大腸桿菌DH5α感受態細胞,然后在含有氨芐的LB平板上培養,挑取單克隆,經菌落PCR檢測后選擇陽性克隆送Sunbio公司測序。
16WRKY7基因的生物信息學分析
用NCBI網站ORF Finder(http://www.ncbi.nlm.nih.gov/gorf/orfig.cgi)尋找WRKY7基因的編碼框。
17實時熒光定量PCR
為了檢測人參WRKY7基因的表達情況,利用實時熒光定量PCR方法檢測該基因經苯甲酸脅迫誘導后在人參根部的表達量。根據WRKY7基因序列設計實時熒光定量PCR引物,將人參40S核糖體蛋白基因作為內參基因設計引物(表1)。以苯甲酸處理不同時間后所提取的人參RNA反轉錄cDNA為模板,熒光定量PCR檢測。反應體系20 μL,包括:2×SYBR Premix Ex Taq 10 μL,10 μmol·L-1上下游引物各1 μL,cDNA 2 μL,ddH2O 6 μL 。每個體系至少重復3次。擴增程序:95 ℃預變性3 min;95 ℃變性15 s,59 ℃退火15 s,72 ℃延伸15 s,40個循環;最后60 ℃延伸5 s,4 ℃保溫。
2結果與分析
21人參WRKY7基因的克隆
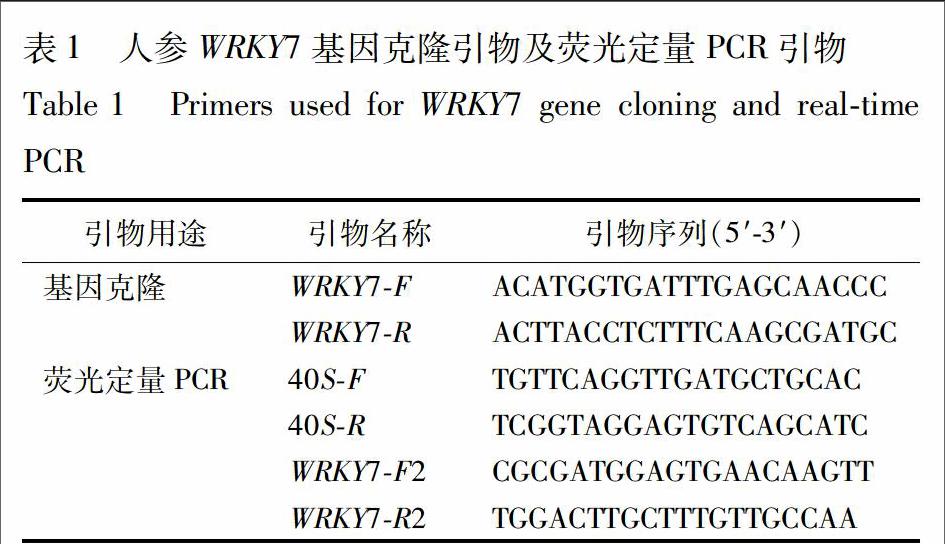
通過將處理組植物RNA送到公司建庫和生物信息學分析,從人參轉錄組數據中發現1條2 053 bp的轉錄本,功能注釋為WRKY轉錄因子,推測其與苯甲酸脅迫響應相關。分析發現,該轉錄本含有1個完整的開放閱讀框(open reading frame,ORF)。根據ORF序列設計基因全長擴增引物WRKY7F和WRKY7R,以苯甲酸脅迫不同時間的反轉錄cDNA為模板,經PCR擴增得到1條長1 216 bp條帶(圖1)。測序結果經ORF Finder分析,確定其開放閱讀框為1 014 bp,編碼337個氨基酸。為區別已知人參WRKY基因,將擴增出的基因暫命名為WRKY7(GenBank登錄號 KX255632)。
22人參WRKY7基因編碼蛋白質特性分析
221人參WRKY7編碼氨基酸序列分析通過NCBI開放閱讀框分析表明其ORF為1 014 bp,蛋白質預測其編碼337個氨基酸。其中第129~132位分別為色氨酸(W),精氨酸(R),賴氨酸(K),酪氨酸(Y),符合WRKY基因家族標志性結構,其下游有典型的“C2HC”型鋅指結構,說明該蛋白屬于WRKY家族第Ⅲ類。
222WRKY7編碼蛋白理化性質分析利用ExPASy Proteomics Server提供的在線軟件Protparam預測WRKY7編碼蛋白的理化性質。WRKY7編碼蛋白由337個氨基酸編碼,分子式為C1669H2619N463O539S14,總原子數為5 304。相對分子質量38243 7 kDa,理論等電點pI為563,帶正電殘基(Arg+Lys)為39,帶負電殘基(Asp+Glu)為46。該蛋白的不穩定系數為4798,表明WRKY7編碼蛋白不穩定。脂肪系數為6564,總平均親水性系數為-0741。
223WRKY7編碼蛋白二級結構分析及功能結構域分析利用SPOMA對WRKY7編碼蛋白進行二級結構分析。WRKY7編碼蛋白二級結構中α螺旋(alpha helix)占2255%、延伸鏈(extended strand)占2255%、β折疊(beta turn)占712%、無規卷曲(random coil)占4777%。InterProScan分析發現,WRKY7編碼蛋白含有1個保守的WRKY結構域(WRKY domain,IPR003657),位于第114~186位氨基酸殘基(圖2)。
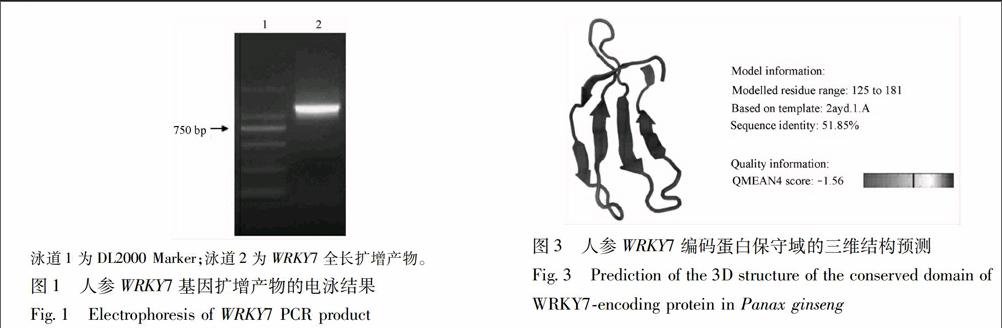
224WRKY7編碼蛋白三維建模用SWISSMODEL三維建模工具作圖,以擬南芥WRKY transcription factor 1為模板(PDB id:2ayd),對人參WRKY7編碼蛋白進行三維結構建模,預測其空間結構(圖3)。
225WRKY7編碼蛋白信號肽、亞細胞定位、跨膜預測分析利用SignalP40 Server預測WRKY7編碼蛋白不具有信號肽。利用在線工具WOLF PSORT預測該蛋白的亞細胞定位情況,結果顯示其細胞核的定位系數為13(nucl:13),表明該蛋白最可能定位于細胞核。TMHMM Server V20預測結果表明,WRKY7編碼蛋白無跨膜區。
226WRKY7基因序列相似性分析同源性分析結果顯示,發現該基因序列與人參Pginseng WRKY6(KT2775471)有87%的相似度;與西洋參P quinquefolius WRKY6(JF9271621),WRKY9(JF9271651)也有87%的相似度;與杭菊Chrysanthemum morifolium WRKY3(KC2922151),WRKY9(KC6153631)有75%的相似度;與大豆Glycine max WRKY60(NM_0012517481)有75%的相似度。WRKY7編碼的氨基酸序列在NCBI中分析發現,該蛋白與人參P ginseng相似性最高(82%,ALS204001),其次為西洋參P quinquefolius(81%,AEQ290191),龍眼Dimocarpus longan(51%,AEO314762),胡楊Populus euphratica(51%,XP_0110378711),葡萄Vitis aestivalis(51%,AAR924771)等。
227氨基酸多序列比對及系統進化樹分析從NCBI比對結果中選出9條與WRKY7編碼蛋白同源性較高的氨基酸序列,經多序列比對分析發現,其與人參、西洋參WRKY基因有著廣泛的相似度,其N端含有WRKY家族標志性片段WRKYGQK,C端鋅指結構為CX7CX23HX1C(圖4)。說明其屬于WRKY家族第Ⅲ類轉錄因子。上述基因的WRKY結構域高度保守,其他區域的可變性較高。
紅色為相對保守的氨基酸殘基;藍色為保守度低的氨基酸殘基;灰色為殘基數量無法完全匹配的序列;少于50%的序列無法匹配的部分用大寫字母表示,多于50%的序列無法匹配的部分用小寫字母表示。
為進一步了解人參WRKY7與其他WRKY家族成員的系統進化關系,從NCBI數據庫下載了其他19個物種23條WRKY家族成員的蛋白序列,Mega 70繪制的NeighborJoining系統進化樹顯示,人參WRKY7與西洋參PqWRKY6、PqWRKY9親緣關系最近(圖5)。這暗示著它們可能具有相似的生物學功能。
23苯甲酸脅迫下人參WRKY7基因的表達分析
為驗證人參WRKY7在苯甲酸脅迫下的表達情況,對該基因進行了誘導表達分析(圖6)。其中0 d樣品為混樣,對照組和處理組共用一個。結果顯示,苯甲酸脅迫誘導后WRKY7的表達水平明顯上調,在第5天和第9天時表達量最高。同時與對照相比,處理組的表達量在第3,5,9天時分別是對照組的
3討論
植物WRKY轉錄因子基因以家族的形式存在,作為一種重要的誘導型調節因子,參與多種植物抗逆反應及生長發育與生理生化代謝過程。目前,WRKY轉錄因子已被證明參與了植物響應生物脅迫[13]、非生物脅迫[14]、生長發育[15]、衰老[16]等生理過程。當植物受到外源物質脅迫時,植物細胞膜上某些受體會被識別,進而激活促分裂原激活蛋白激酶信號通路,通過磷酸化或去磷酸化將胞外信號傳導入細胞核內,抑制或激活WRKY轉錄因子的表達[17],進而與下游目標基因啟動機子區的Wbox進行特異性結合[18],從而調控目標基因的表達。
本研究通過對苯甲酸脅迫人參轉錄組的生物信息學分析,從中發現了一個WRKY轉錄因子基因WRKY7,其與人參WRKY6基因(KT2775471)相似度為87%,所編碼的氨基酸序列相似度為82%。氨基酸序列分析發現,WRKY7有一個高度保守的WRKY結構域,下端鋅指結構為CX7CX23HX1C,屬于′C2HC′型,為WRKY家族的第Ⅲ類。該基因與擬南芥AtWRKY53有較高的同源性,AtWRKY53被報道受到干旱[19]、機械損傷[20]等誘導,故而推測人參WRKY7基因可能也具有類似功能。研究發現,苯甲酸脅迫后人參WRKY7基因的表達水平顯著上調,暗示其在人參響應自毒物質脅迫過程中扮演重要角色。WRKY7有可能進一步啟動了下游抗逆基因的表達,從而提高了植物對苯甲酸脅迫的耐受性。
目前,針對人參化感自毒作用分子機制的研究尚處于起步階段,本課題組前期通過分析化感自毒物質誘導下人參轉錄組數據的差異,發現了大量與人參生長發育、抗病抗逆等密切相關的差異性表達基因,但對于這些轉錄組數據的挖掘還不夠深入,對于關鍵基因響應化感自毒物質的通路尚不明確。WRKY作為參與植物非生物脅迫響應的一類重要轉錄因子,其在響應自毒脅迫中的作用值得關注。本研究后續將構建過量表達載體,并將WRKY7基因轉入擬南芥,通過重組體篩選及脅迫測驗,進一步研究該基因的生物學功能。同時會繼續對已獲得的轉錄組數據進行深入分析,探究不同轉錄因子基因在苯甲酸脅迫下的生物調控網絡,為深入闡釋人參響應化感自毒物質的信號轉導途徑,以及自毒脅迫響應的分子機制提供理論依據。
[參考文獻]
[1]Ishiguro S, Nakamura K Characterization of a cDNA encoding a novel DNAbinding protein, SPF1, that recognizes SP8 sequences in the 5′ upstream regions of genes coding for sporamin and betaamylase from sweet potato[J] Mol Gen Genet, 1994, 244(6):563
[2]Wu K L, Guo Z J, Wang H H, et al The WRKY family of transcription factors in rice and Arabidopsis and their origins[J] DNA Res, 2005, 12(1):9
[3]Ross C A, Liu Y, Shen Q J, et al The WRKY gene family in rice (Oryza sativa)[J] J Integr Plant Biol, 2007, 220(6):827
[4]Eulgem T, Rushton P J, Robatzek S, et al The WRKY superfamily of plant transcription factor[J] Trends Plant Sci, 2000, 5(5):199
[5]Eulgem T Regulation of the Arabidopsis defense transcriptome[J] Trends Plant Sci, 2005, 10(2):71
[6]Zheng Z, Qamar S A, Chen Z, et al Arabidopsis WRKY33 transcription factor is required for resistance to necrotrophic fungal pathogens[J] Plant J Cell Mol Biol, 2006, 48(4):592
[7]lker B, Somssich I E WRKY transcription factors: from DNA binding towards biological function[J] Curr Opin Plant Biol, 2004, 7(5):4919
[8]Suzuki N, Shulaev V, Mittler R Enhanced tolerance to environmental stress in transgenic plants expressing the transcriptional coactivator multiprotein bridging factor 1c[J] Plant Physiol, 2005, 139(3):1313
[9]Zhou Q Y, Tian A G, Zou H F, et al Soybean WRKYtype transcription factor genes, GmWRKY13, GmWRKY21, and GmWRKY54, confer differential tolerance to abiotic stresses in transgenic Arabidopsis plants[J] Plant Biotechnol J, 2008, 6(5):486
[10]黃小芳 人參根系分泌物及其自毒活性研究[D] 北京:北京協和醫科大學, 2009
[11]李勇, 黃小芳, 丁萬隆 營養元素虧缺對人參根分泌物主成分的影響[J] 應用生態學報, 2008, 19(8):1688
[12]Wu B, Long Q L, Gao Y, et al Transcriptome response of Panax ginseng roots, stems and leaves exposed to benzoic acid revealed the molecular mechanism of autotoxicity[J] BMC Genomics, 2015, 16:1010
[13]Peng X X, Tang X K, Zhou P L, et al Isolation and expression patterns of rice WRKY82 transcription factor gene responsive to both biotic and abiotic stresses[J] Agr Sci China, 2011, 10(6):893
[14]冉昆, 王少敏, 魏樹偉, 等 植物非生物脅迫相關的WRKY轉錄因子研究進展[J] 青島農業大學學報, 2014, 31(3):217
[15]徐文靜 WRKY轉錄因子在水稻生長發育及與白葉枯病菌互作中的表達研究[D] 保定:河北農業大學, 2012
[16]Miao Y, Laun T, Zimmermann P, et al Targets of the WRKY53 transcription factor and its role during leaf senescence in Arabidopsis[J] Plant Mol Biol, 2004, 55(6):853
[17]Kim C Y, Zhang S Activation of a mitogenactivated protein kinase cascade induces WRKY family of transcription factors and defense genes in tobacco[J] Plant J, 2004, 38(1):142
[18]Laloi C, MestresOrtega D, Marco Y, et al The Arabidopsis cytosolic thioredoxin h5 gene induction by oxidative stress and its Wboxmediated response to pathogen elicitor[J] Plant Physiol, 2004, 134(3):1006
[19]Sun Y, Yu D Activated expression of AtWRKY53 negatively regulates drought tolerance by mediating stomatal movement[J] Plant Cell Rep, 2015, 34(8):1
[20]Yong H C, Sheng L Transcriptional profiling reveals novel interactions between wounding, pathogen, abiotic stress, and hormonal responses in Arabidopsis[J] Plant Physiol, 2002, 129(2):66
[責任編輯呂冬梅]
