廣西馬尾松第2代育種群體的組建*
2017-03-08 07:18:09馮源恒李火根楊章旗黃永利羅群鳳
林業科學 2017年1期
馮源恒 李火根 楊章旗 黃永利 羅群鳳 張 遠
(1. 廣西壯族自治區林業科學研究院 國家林業局馬尾松工程技術研究中心 廣西馬尾松工程技術研究中心 南寧 530002; 2. 南京林業大學林木遺傳與生物技術省部共建教育部重點實驗室 南京 210037; 3.南寧市林業科學研究所 南寧 530107)
廣西馬尾松第2代育種群體的組建*
馮源恒1,2李火根2楊章旗1黃永利3羅群鳳1張 遠2
(1. 廣西壯族自治區林業科學研究院 國家林業局馬尾松工程技術研究中心 廣西馬尾松工程技術研究中心 南寧 530002; 2. 南京林業大學林木遺傳與生物技術省部共建教育部重點實驗室 南京 210037; 3.南寧市林業科學研究所 南寧 530107)
【目的】 基于廣西馬尾松第1代育種群體的8片20年生及以上的子代測定林測定試驗,在綜合評價育種目標性狀與遺傳多樣性的基礎上,選擇建立馬尾松第2代育種群體,為馬尾松高世代育種研究提供重要材料。【方法】 采用SAS分析軟件依據線性模型對子代生長量數據進行統計分析,并據此進行第2代優樹選擇。采用SSR分子標記對第2代優樹進行遺傳多樣性分析、親本分析及遺傳距離估算,根據優樹間遺傳距離對第2代育種群體進行結構劃分。【結果】 在參試的子代測定林中,參試家系間在生長性狀上均達到極顯著差異; 多數子代林的材積指標家系遺傳力處于中等以上水平(h2≥0.2),適于開展優良家系選擇。基于此,采用配合選擇與單株選擇相結合的方法選擇出第2代育種群體材料163株,平均遺傳增益為21.95%。采用16對SSR引物對該第2代育種材料進行遺傳多樣性研究,16個位點共檢測到45個等位變異。每個位點平均觀察等位基因數(NA)為2.7,多態率為100%; 平均有效等位基因數(Ne)為1.54; Shannon多樣性指數(I)平均為0.49; 平均觀測雜合度(Ho)為0.32。采用Coancestry Version 1.0軟件計算出第2代育種群體的平均共祖系數為0.042,育種群體狀態數為11.9; 根據16對SSR引物的擴增結果,采用CERVUS2.0軟件對構建的第2代育種群體材料進行父本分析,發現在包含163個優樹的第2代群體中,有57個個體在置信度95%的情況下可以確定父本,另有102個個體在置信度80%的情況下能夠確定父本。為了有效避免高世代雜交育種過程中發生近交,以遺傳距離為指標對第2代育種群體材料進行聚類,根據距離聚類結果將163個體劃分為10個亞系,編號為桂GC2-A—桂GC2-J。在建立馬尾松第2代種子園時,擬采用以下策略: 從每個亞系中選擇一定數量的最佳無性系作為精選群體建園; 在進行下一代雜交育種時,進行亞系間的交配,使育種群體整體的近交程度保持在一個相對較低的水平。【結論】 根據研究結果初步建立了由163株優樹組成的廣西馬尾松第2代育種群體。該群體具有較高的遺傳多樣性,個體間的近交程度較低。根據遺傳距離對第2代育種群體進行亞系劃分,設計出“系間雜交、系內慎交”的高世代雜交育種策略,可有效避免近交,為有計劃地開展馬尾松高世代雜交育種奠定基礎。
馬尾松; 育種群體; SSR; 遺傳多樣性; 親本分析
馬尾松(Pinusmassoniana) 是我國分布最廣的針葉樹種,也是我國南方地區重要的用材、荒山造林和工業原料樹種,其木材和松脂是許多森林工業、林產工業和造紙工業的支柱(周政賢, 2000)。馬尾松于20世紀70年代末列為主要樹種開展研究(周志春等, 1997),歷經“六·五”至“十二·五”近40年的時間,我國馬尾松育種研究取得了豐富的成果。其中,在通用材改良階段(“六·五”和“七·五”計劃期間)以生長量為主要研究性狀,重點開展了速生種源的選擇,完成了馬尾松種源區劃(全國馬尾松種源試驗協作組, 1987),評選出云開大山、南嶺山地等一批馬尾松優良種源(榮文琛等, 1994),并在南方各主要產區建立了一批初級種子園(王章榮等, 1990);在定向改良階段(“八·五”計劃以來),以紙漿材和產脂林為主要目標,開始了定向選育工作,比較系統地研究了木材材性、產脂能力及紙漿性能在不同層次的變異模式和遺傳規律(曾令海等, 1994; 秦國峰等, 1995)。在育種材料收集、育種群體和生產群體建立方面,據不完全統計,各省區共選出優樹5 500多株,建立收集區100 hm2,建立初級無性系種子園和實生種子園1 100 hm2(周志春等, 1997)。浙江等省份已經開展了馬尾松第2代育種研究(張一等, 2010),并建立了第2代種子園(譚小梅等, 2012)。但是對于如何制定馬尾松的高世代育種策略,各地都還沒有進行比較系統的研究(王章榮, 2012)。
廣西是馬尾松主要分布區,也是馬尾松最重要的優良種源區,馬尾松遺傳育種資源極其豐富(楊章旗等, 2001)。廣西馬尾松第1代育種群體于20世紀80年代初完成選擇收集工作,共有464個無性系; 并于80年代末陸續開展子代測定試驗,到21世紀初最早的幾個子代測定林都已達到了15年生的速生期,其生長數據比早期測定時更有價值,為組建高世代育種群體提供了豐富的材料。建立高世代育種群體,應重視群體規模和群體結構這2個關鍵因素。一方面,應滿足長期育種的靈活性保持足夠大的群體規模。維持群體的一定多樣性、長期育種的遺傳增益及開展多個性狀的選擇都需要較寬的遺傳基礎。已有研究結果認為,如果希望有效群體的家系數大于100,遺傳力大于0.25,那么開始選擇的親本應該在300個左右(Jacksonetal., 1972; Cotterilletal., 1989; Lindgrenetal., 1997)。美國東南部地區濕地松(P.elliottii)和火炬松(P.taeda)第2代育種群體、澳大利亞輻射松(P.radiata)育種群體都是由上百株到上千株優樹組成(王章榮, 2012)。另一方面,高世代育種群體也應滿足在近期育種階段內獲得最大增益的要求,并考慮節約育種成本。因此,為了實現有效管理和系統制定雜交計劃,在火炬松、濕地松等樹種的研究中往往將龐大的群體更為精細地劃分為若干個亞系或育種組(Whiteetal., 1993; 2011)。所以建立較大規模育種群體是進行長期可持續多目標遺傳改良的基礎,而對育種群體進行合理的結構劃分是保障高世代育種有計劃進行的重要條件。
本研究以8片林齡20年以上的子代測定林為材料,在基于表型子代測定數據進行單株選擇的基礎上以SSR分子標記為技術手段對第2代育種材料進行父本分析及遺傳多樣性研究,為馬尾松高世代育種研究提供重要材料;并對第2代育種群體進行結構劃分,為高世代育種方案的有效制定提供條件。
1 材料與方法
1.1 試驗材料
本研究調查了南寧市林業科學研究所分別于1988,1992,1994年營建的8個馬尾松第1代無性系種子園的子代測定林(表1),共計自由授粉家系440個(不同年份的子代測定試驗存在相同參試家系)。基于對生長表型數據的統計分析結果選擇子代林中的優良單株作為第2代育種材料,并以無性系形式保存在南寧市林業科學研究所馬尾松種質資源庫。而后采集這些優良無性系的針葉,提取DNA。由于所有的第2代育種材料均選自自由授粉家系,為使其遺傳背景更加清晰,研究中還采集了第1代育種群體全部476個無性系的針葉,提取其DNA用于第2代育種材料的親本分析。同時采集163株第2代候選優樹的針葉,提取DNA。
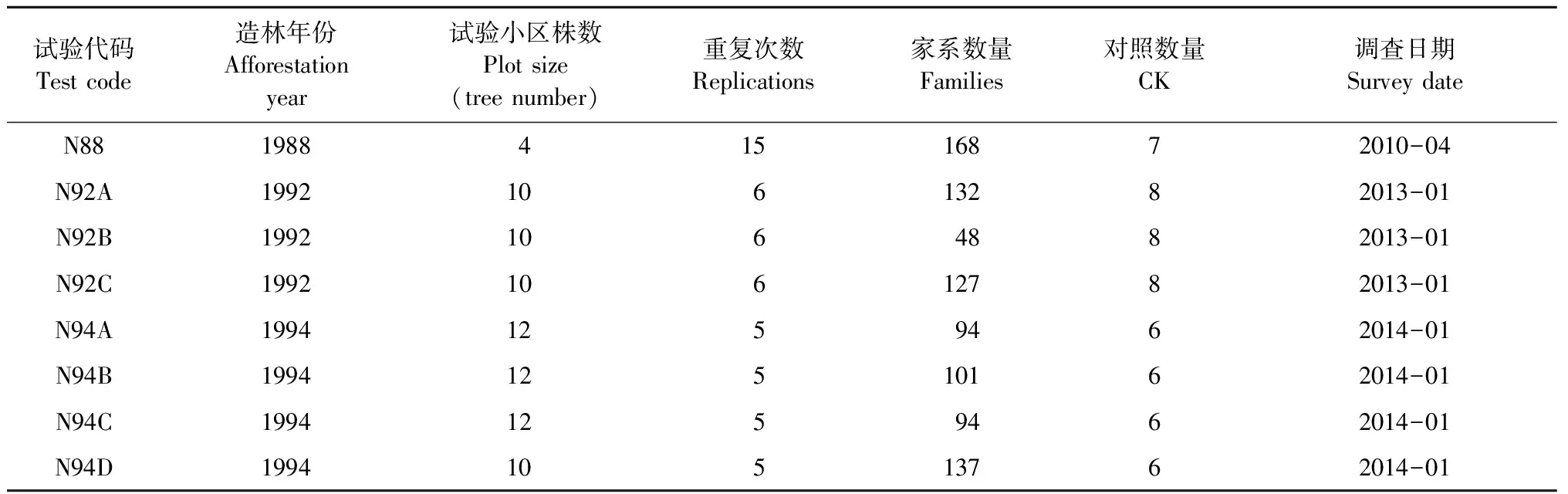
表1 各子代測定試驗概況
1.2 數據調查與統計分析
N88試驗采用2010年測定數據,N92系列3個試驗采用2013年測定數據,N94系列4個試驗采用2014年測定數據,測定樹高、胸徑等指標,并通過下式計算材積(V):
(1)
式中:D為胸徑;H為樹高。
采用SAS8.1軟件,以yijk=μ+Bi+Fj+BFij+Eijk為線性模型對各子代測定林數據分別進行方差分析。式中:yijk表示第i區組第j家系第k個體的表型值;u表示總體平均值;Fj表示家系效應;Bi表示區組效應;BFij表示區組×家系效應;Eijk表示隨機誤差。并根據方差分析結果進一步估算遺傳力:
(2)
(3)
(4)
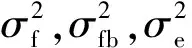
單株選擇所得遺傳增益(ΔGi)及配合選擇所得遺傳增益(ΔGf)(黃少偉等, 2006)的估算公式為:
(5)
(6)
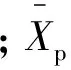
變異系數的計算公式為:
(7)
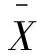
1.3 第2代育種材料的選擇方法
采用以下3種方法篩選第2代育種群體材料:一是采用配合選擇(combined selection),即從優良家系中選擇優良單株,以測定數據為參考,從每個子代測定林材積均值最大的前20%家系中選擇單株材積排名在全林分的前15%、干形和冠形均優秀的個體且排除孤立木與邊緣木;二是在全部試驗林內開展優良單株選擇,從每個試驗中單株材積排名在全林分的前2%單株中選擇干形、冠形均優秀的個體;三是在全部試驗林內開展優良干形單株選擇,從每個試驗中單株材積排名在全林分的前20%單株中選擇干形完全通直優秀的個體。
1.4 遺傳多樣性及親本分析
1.4.1 DNA的提取 松樹針葉DNA提取采用CTAB裂解-硅珠吸附法(Doyleetal., 1990)。純化后DNA經分光光度計檢測純度后置于4 ℃冰箱保存備用。
1.4.2 引物來源及SSR-PCR反應條件 所用的SSR 引物根據馬尾松基因組DNA 測序所得序列設計開發,共計318對(Fengetal., 2013)。篩選16對多態性高的SSR引物用于本研究。PCR反應體系為10 μL: Tris-HCl 10 mmol·L-1pH8.0,KCl 50 mmol·L-1, Mg2+2.5 mmol·L-1,dNTP(dATP,dCTP,dGTP,dTTP各0.2 mmol·L-1),引物2.5 pmol,Taq聚合酶0.8 U,DNA 10~20 ng。擴增反應程序采用Touch-down PCR: 94 ℃15 s,60 ℃15 s(Δt=-0.5 ℃),72 ℃30 s,16個循環; 再進入94 ℃15 s,52 ℃15 s,72 ℃30 s,10個循環; 最后72 ℃延伸15 min。SSR-PCR產物在8%的聚丙烯酰胺凝膠上電泳,銀染檢測并照相記錄。
1.4.3 數據分析 采用POPGENE32軟件(Yehetal., 1997)計算群體內的觀察等位基因數目(NA)、有效等位基因數目 (Ne)、 Shannon多樣性指數(I)、各位點的觀察雜合度(Ho)。采用Coancestry Version 1.0軟件(Wang, 2011)計算群體內的共祖系數Θ(Ritland, 2009; Lynchetal., 1999; Milligan, 2003; Wang, 2007)。利用基于最大似然法的CERVUS2.0軟件,對第2代優樹進行父本分析。該軟件的原理參見文獻(Marshalletal., 1998; 何田華等, 2001)。
2 結果與分析
2.1 馬尾松生長性狀的遺傳力
采用SAS8.1軟件對8個子代測定林的樹高、胸徑和材積3項指標進行方差分析并估算遺傳力(表2)。方差分析結果表明,在所有子代試驗中3項指標在家系間均存在極顯著差異(α<0.01)。這說明產自馬尾松第1代育種群體的自由授粉家系間存在豐富的遺傳變異,有很大遺傳改良潛力。

表2 馬尾松各子代測定試驗遺傳分析
2.2 優樹選擇與增益
2.2.1 配合選擇 以測定數據為參考,從每個試驗材積均值最大的前20%家系中選擇單株材積、干形、冠形等綜合表現最為優秀的1個單株,共選出95株優樹作為馬尾松第2代育種群體材料;平均增益為11.53%~45.37%(表3)。
2.2.2 單株選擇 以測定數據為參考,從每個試驗單株材積排名在全林分的前2%單株中選擇干形、冠形均優秀的個體(入選家系與配合選擇不重復),共選出40株優樹作為馬尾松第2代育種材料收入馬尾松基因庫保存;平均增益為12.71%~57.64%(表4)。
2.2.3 優良干形單株選擇 以測定數據為參考,從每個試驗單株材積排名在全林分的前20%單株中選擇干形完全通直的優秀個體(入選家系與配合選擇、單株選擇不重復)。除N92B試驗因候選優樹與配合選擇和單株選擇結果完全重復沒有新優樹入選外,其他7個試驗林共選出28株優樹作為馬尾松第2代育種材料收入馬尾松基因庫保存;平均增益為8.25%~40.25%(表5)。
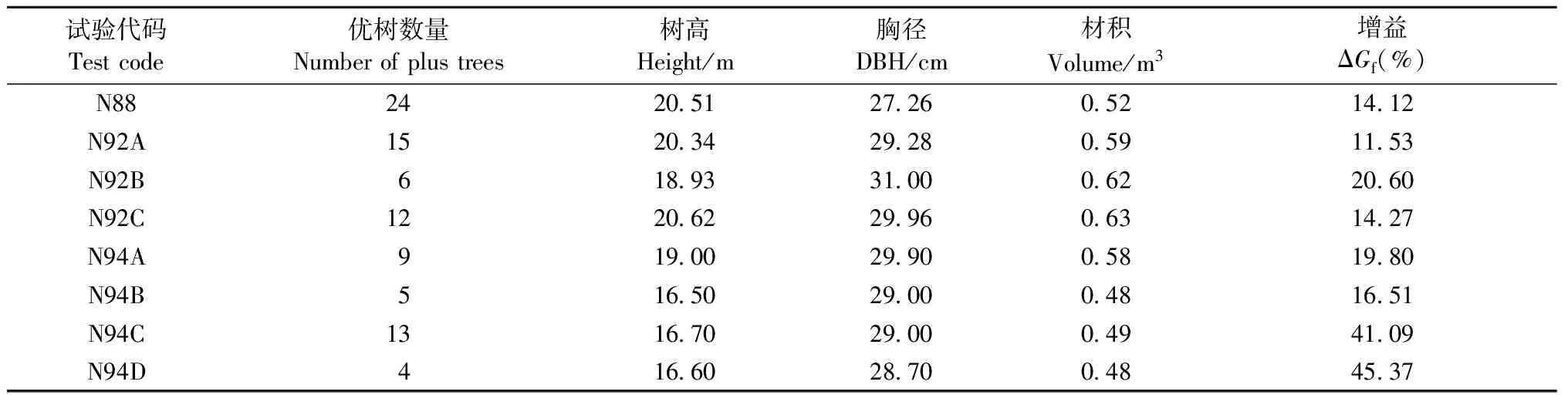
表3 配合選擇結果
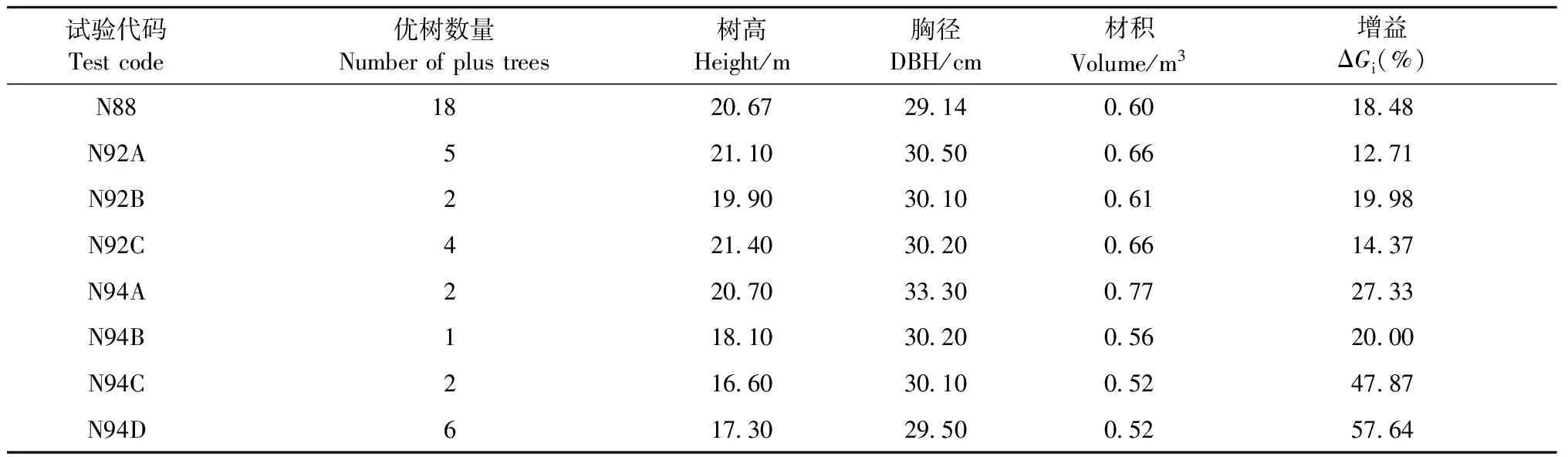
表4 單株選擇結果
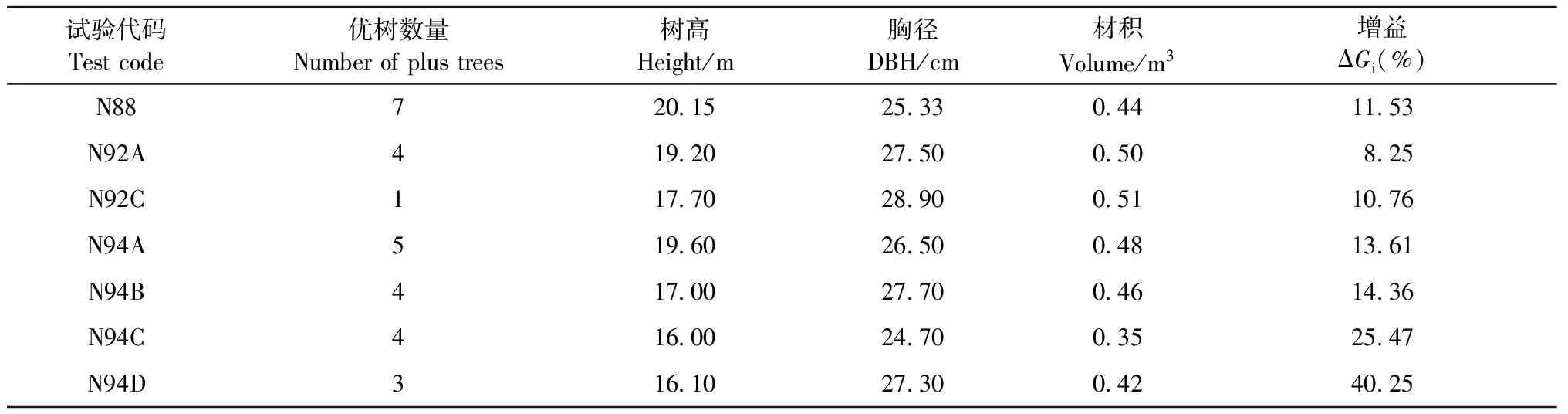
表5 干形選擇結果
2.3 第2代育種群體遺傳多樣性分析
采用16對SSR引物在163個樣本中共檢測到16個位點45個等位變異,多態率為100%。每個位點平均觀察等位基因數(NA)為2.7,平均有效等位基因數(Ne)為1.54。表6列出了馬尾松第2代育種群體不同位點的遺傳多樣性參數。不同位點的多樣性參數差別很大,PF 784位點的Shannon多樣性指數(I)最高(1.03),PF 620位點的多樣性指數最低(0.04),平均為0.49,平均觀測雜合度為0.32。
采用Coancestry Version 1.0軟件計算得到第2代育種群體163個無性系在16個位點的平均共祖系數為0.042,育種群體狀態數為11.9。
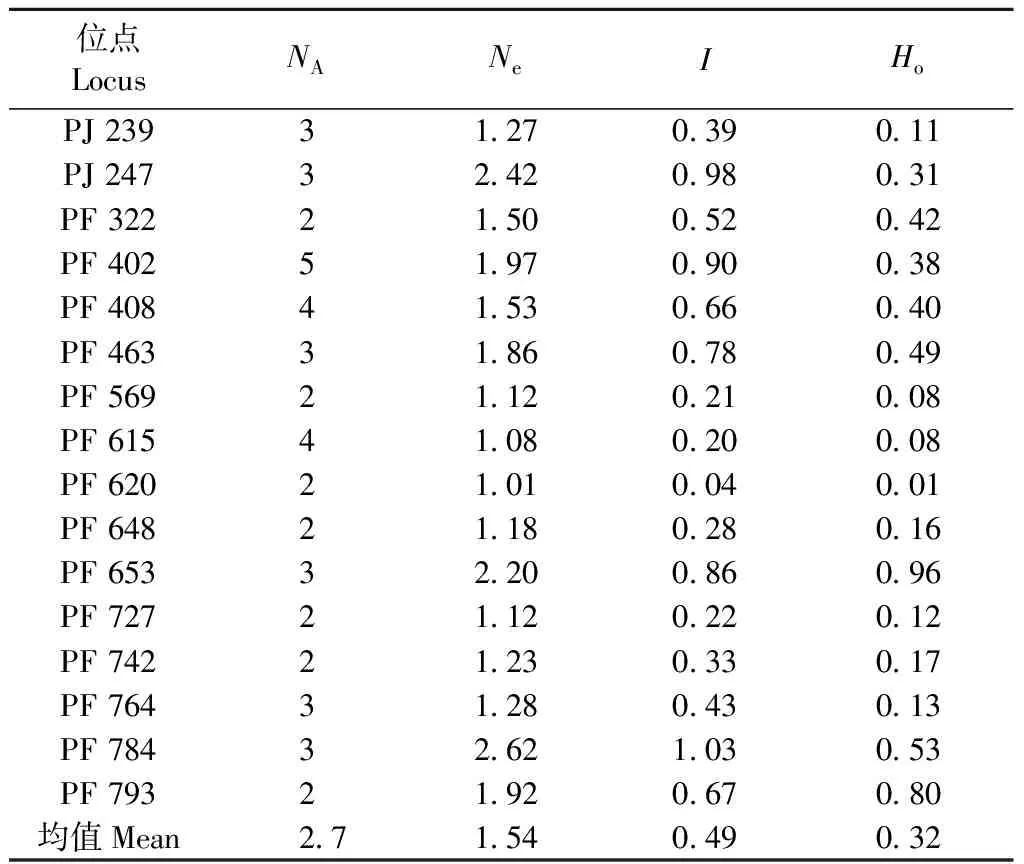
表6 馬尾松第2代育種群體在16個遺傳位點的遺傳多樣性*NA: 平均位點觀察等位基因數Allele number; Ne: 平均有效等位基因數Effective allele number; I: 平均Shannon 多樣性指數Shannon index; Ho: 平均觀測雜合度Observed heterozygosity.
2.4 第2代育種材料的親本分析
根據16對SSR引物的擴增結果,采用CERVUS2.0軟件對得到的第2代育種材料進行父本分析。163個第2代群體成員中有57個個體在置信度95%的情況下可以確定父本,另有102個個體在置信度80%的情況下能夠確定父本。根據父本分析結果,父母本明確的159個第2代育種材料共來自201個親本,其中母本129個、父本114個。在所有的親本中,桂GC800A的父本貢獻率最高(4.14%),共有7個子代;桂GC443A的母本貢獻率最高(2.37%),共有4個子代。
2.5 第2代育種群體的亞系劃分
采用POPGENE32軟件計算得到163個無性系間的Nei’s 標準遺傳距離(GD),共得到13 366對無性系間遺傳距離。其中桂GC269D與桂GC848D間GD最大,為0.56; 最小的GD出現在桂GC849A與桂GC858A間,為0.014。采用軟件NTSYS,根據Nei’s遺傳距離,按非加權類平均法(UPGMA)對163個無性系進行聚類分析。根據聚類結果將遺傳距離等于0.06作為劃分亞系的最低標準,即GD小于0.06的分支不再獨立劃分亞系。163個無性系分為10個亞系(GC2-A—GC2-J),每個亞系無性系數目5~34個不等。由于GC2-A,GC2-C,GC2-D,GC2-E,GC2-F,GC2-I,GC2-J所含個體較多,為了減少因亞系過大而帶來的雜交設計難度,又在這些亞系內按遺傳距離分為2~4個組,每組5~8個無性系。在此基礎上,為便于開展亞系間雜交設計,將亞系及亞系內組重新進行聚類分析(圖1),簡化并明晰各個亞系及其組間的遺傳關系。由于亞系間親緣關系較遠,進行第3代雜交育種可以組為單位在不同亞系間進行雜交設計,而亞系內親緣關系較近不宜進行系內雜交。
3 討論
3.1 第2代育種材料的選擇
林木改良中常以全同胞子代選育第2代育種材料,這樣既可以獲得一般配合力又可以獲得特定雜交組合產生的特殊配合力,而且子代遺傳譜系清楚,在改良過程中可以有效地避免近交。但該策略的缺陷在于雜交成本較高,難以進行大規模雜交試驗,而小規模的雜交試驗因為親本數量有限,往往導致子代的遺傳基礎過窄,不利于更高世代的改良;同時風媒傳粉的樹種在雜交過程中在很大程度上還要受到花粉污染的干擾。而以半同胞子代選育第2代育種材料,不受雜交試驗的高成本限制,參與親本數量大,子代遺傳多樣性較高; 其缺點是所獲遺傳增益可能較全同胞子代相對較低,且父本不明確,但通過提高選擇強度和進行親本分析可以彌補上述缺點。
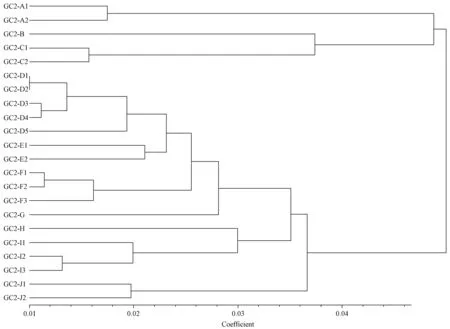
圖1 馬尾松第2代育種群體各亞系遺傳聚類Fig.1 Each substrain genetic distance in second generation of Masson pine breeding population
具體就廣西馬尾松而言,以林齡20年以上的半同胞測定林作為材料選擇第2代育種群體,既節省育種成本又縮短了育種周期,而且進入速生期后的表型表現比早期測定更有價值。另一方面,由于運用SSR分子標記技術,可以較為準確地推測父本,使第2代育種材料的親緣關系更為清楚。
3.2 第2代育種群體的遺傳多樣性水平
本研究構建的第2代育種群體的平均等位基因數為2.45,多態率為100%,平均有效等位基因數(Ne)為1.56,Shannon多樣性指數(I)平均為0.49,平均觀測雜合度為0.32,均低于廣西馬尾松第1代育種群體,這說明在較大選擇強度下會損失一部分遺傳多樣性。而這一結果高于浙江淳安馬尾松第2代育種群體(I=0.476)(譚小梅等, 2012),說明廣西馬尾松第2代育種群體的遺傳多樣性仍處于較高水平。
3.3 第2代育種群體的亞系劃分
通過種內雜交獲得增益的一般方法是:一方面將育種群體分組成亞系并隔離; 另一方面對組內成員間選用適當的交配設計開展雜交。整個育種群體分為主群體和精選群體2大部分。主群體各個亞系的成員按育種值高低進行排名,由高育種值的個體組成精選群體建立種子園。高世代育種時每個亞系各自進行系內雜交選擇優秀子代組建該亞系的第2代群體,再從各個亞系選出最好的個體組建第2代精選群體,這樣可以保證精選群體個體間近交系數維持在低水平; 但缺點是如果亞系內成員數目較少則經過2,3個世代亞系內的近交系數會明顯上升,若亞系內成員數目較多雜交的工作將十分巨大。美國的火炬松與濕地松均采用這種策略(Whiteetal., 1993; 2011)。
本研究則根據遺傳距離將親緣較近的個體劃歸一個亞系,在建立種子園時從每個亞系中選擇一定數量的最佳無性系作為精選群體建園。在進行下一代雜交育種時,不進行亞系內交配,而以亞系為單位進行亞系間的交配。雜交方案以測交設計為主,即所有母本來自同一個亞系而所有父本來自另一個亞系,這樣可以保證雜交雙親的親緣關系較遠。從雜交子代中選擇優良個體組建第3代育種群體后繼續根據遺傳距離劃分亞系,這樣每一代的精選群體近交程度會比上一代有所上升,但育種群體整體的近交程度卻能保持在一個相對較低的水平。
4 結論
采用配合選擇與單株選擇相結合的方法進行馬尾松第2代育種材料選擇,可以較好地兼顧育種群體的遺傳增益與遺傳多樣性,據此初步建立了由163株優樹組成的廣西馬尾松第2代育種群體。該群體具有較高遺傳多樣性,個體間的近交程度較低。根據遺傳距離對第2代育種群體進行亞系結構劃分,設計出“系間雜交、系內慎交”的高世代雜交育種策略,可以有效地避免近交,為馬尾松有計劃地開展高世代雜交育種奠定基礎。
何田華,葛 頌. 2001.植物種群交配系統、親本分析以及基因流動研究.植物生態學報, 25(2):144-154.
(He T H, Ge S. 2001. Mating system, paternity analysis and gene flow in plant populations. Acta Phytoecologica Sinica, 25(2):144-154. [in Chinese])
黃少偉, 鐘偉華, 陳炳銓. 2006.火炬松半同胞子代配合選擇的遺傳增益估算. 林業科學, 42(4): 33-37.
(Huang S W, Zhong W H, Chen B Q. 2006. Estimation on genetic gains of combined selection for growth traits of half-sib progeny ofPinustaeda. Scientia Silvae Sinicae, 42(4):33-37. [in Chinese])
秦國峰,周志春,李光榮,等. 1995.馬尾松造材紙的變異、遺傳和選擇育種策略.林業科學研究, 8(5): 506-513.
(Qin G F, Zhou Z C, Li G R,etal. 1995.Variation, genetics and strategy on selection breeding of pulp wood of Masson pine. Forest Research, 8(5): 506-513. [in Chinese])
全國馬尾松種源試驗協作組. 1987.馬尾松種源變異及種源區劃分的研究. 亞熱帶林業科技,15 (2):81-89.
(Provenance Test of Masson Pine National Cooperative Group. 1987. Study on provenance variation and the provenance division of Masson pine. Subtropical Forestry Science and Technology, 15(2):81-89.[in Chinese])
榮文琛,岳水林,趙世遠, 等. 1994.馬尾松造林區優良種源選擇. 林業科學研究,7(5):542-549.
(Rong W C, Yue S L, Zhao S Y,etal.1994. Selection of fine provenance of Masson pine for afforestation regions. Forest Research, 7(5):542-549. [in Chinese])
譚小梅,周志春,金國慶,等. 2012.馬尾松二代無性系種子園遺傳多樣性和交配系統分析.林業科學, 48(2):69 -74.
(Tan X M, Zhou Z S, Jin G Q,etal. 2012. Genetic diversity and mating system analysis ofPinusmassonianain a second-generation clonal seed orchard. Scientia Silvae Sinicae, 48(2):69 -74. [in Chinese])
王章榮,秦國峰,陳天華. 1990.馬尾松種子園建立技術論文集. 北京:學術書刊出版社.
(Wang Z R, Qin G F, Chen T H.1990. The proceedings of establishing techniques for the seed orchard of Masson pine. Beijing: Academic Books and Periodicals Publishing Company. [in Chinese])
王章榮. 2012.林木高世代育種原理及其在我國的應用. 林業科技開發, 26(1):1-5.
(Wang Z R. 2012. The application of the principle of high generation tree breeding in China. China Forestry Science and Technology, 26(1):1-5. [in Chinese])
楊章旗,覃開展. 2001.馬尾松種質基因資源選擇、保存和利用研究.廣西林業科學, 30(1):19-22.
(Yang Z Q, Qin K Z. 2001.Studies on collection, preservation and utilization of Masson pine germplasm gene resource. Guangxi Forestry Science,30(1):19-22. [in Chinese])
曾令海,李召青,王以珊, 等. 1994.馬尾松高產脂力類型的選擇及子代測定的遺傳評價.廣東林業科技, (2): 35-38.
(Zeng L H, Li Z Q, Wang Y S,etal. 1994. The selection method of Masson pine plus trees with high yield of resin and genetic evaluation of progeny test. Guangdong Forestry Science and Technology, (2): 35-38. [in Chinese])
張 一,譚小梅,周志春, 等. 2010.馬尾松二代育種親本主要生長性狀和ISSR遺傳變異.分子植物育種, 8(3):501-510.
(Zhang Y, Tan X M, Zhou Z C,etal. 2010. Genetic variation of the second-generation breeding population of Masson pine based on major growth traits and ISSR. Molecular Plant Breeding, 8(3):501-510.[in Chinese])
周政賢. 2000.中國馬尾松. 北京:中國林業出版社.
(Zhou Z X. 2000. Chinese Masson pine. Beijing: China Forestry Publishing House. [in Chinese])
周志春,秦國峰,李光榮, 等. 1997.馬尾松遺傳改良的成就、問題和思考. 林業科學研究, 10(4):435-442.
(Zhou Z C, Qin G F, Li G R,etal. 1997. Achievements, problems and its countermeasures of genetic improvement of Masson pine. Forest Research, 10(4):435-442. [in Chinese])
Cotterill P P, Dean C A, Cameron J,etal. 1989. Nucleus breeding: a new strategy for rapid improvement under clonal forestry∥Gibson G L, Griffin A R, Matheson A C,etal. Breeding tropical trees: population structure and genetic improvement strategies in clonal and seedling forestry(proceedings of a conference held in Pattaya, Thailand, 28 November-3 December 1988). Arlington: Winrock International, 39-51.
Doyle J J, Doyle J L. 1990. Isolation of plant DNA from fresh tissue. Focus, 12:13-15.
Feng Y H, Yang Z Q, Wang J,etal. 2013. Development and characterization of SSR markers fromPinusmassonianaand their transferability toP.elliottii,P.caribaeaandP.yunnanensis. GMR, 13(1): 1508-1513.
Jackson N, Turner H N. 1972. Optimal structure for a cooperative nucleus breeding system. Proc Aust Soc Anim Prod, 9: 55-64
Lindgren D, Wei R P, Lee S J. 1997. How to calculate optimum family number when starting a breeding program. Forest Science, 43(2): 206-212.
Lynch M, Ritland K. 1999. Estimation of pairwise relatedness with molecular markers. Genetics, 152(4): 1753-1766.
Marshall T C, Slate J, Kruuk L. 1998. Statistical confidence for likelihood-based paternity inference in natural populations. Mol Ecol, 7(5): 639-655.
Milligan B G. 2003. Maximum-likelihood estimation of relatedness. Genetics, 163(3): 1153-1167.
Ritland K. 2009. Estimators for pairwise relatedness and individual inbreeding coefficients. Genet Res, 67(2): 175-185.
Wang J L. 2007. Triadic IBD coefficients and applications to estimating pairwise relatedness. Genet Res, 89(3): 135-153.
Wang J L. 2011. COANCESTRY: a program for simulating, estimating and analysing relatedness and inbreeding coefficients. Molecular Ecology Resources, 11(1): 141-145.
White T L,Hodge G R,Powell G L.1993. An advanced-generation tree improvement plan for slash pine in the southeastern United States. Silvae Genetica, 4(2): 359-371.
White T L, Huber D A, Powell G L. 2011. Third-cycle breeding strategy for Slash pine by the cooperative forest genetics research program. Digital Library Okstate Edu.
Yeh F C, Yang R C, Boyle T B,etal.1997. POPGENE, the user-friendly shareware for population genetic analysis.Molecular Biology and Biotechnology Centre, University of Alberta, Canada, 10.
(責任編輯 徐 紅)
Construction of Second Generation Breeding Population ofPinusmassonianain Guangxi
Feng Yuanheng1,2Li Huogen2Yang Zhangqi1Huang Yongli3Luo Qunfeng1Zhang Yuan2
(1.GuangxiInstituteofForestryScienceMassonPineEngineeringTechnologyResearchCenterofStateForestryAdministrationMassonPineEngineeringTechnologyResearchCenterofGuangxiNanning530002; 2.KeyLaboratoryofForestGeneticsandBiotechnologyofMinistryofEducationNanjingForestryUniversityNanjing210037; 3.NanningForestryDivisionNanning530107)
【Objective】This study was based on 8 older-than-15-years progeny tests of the first generation breeding population of Masson pine(Pinusmassoniana) in Guangxi. On the basis of comprehensive evaluation of breeding objectives and genetic diversity, a second generation breeding populations of Masson pine was established. 【Method】Progeny growth data was analyzed using the SAS software which was based on linear model, and according to the results, second generation plus trees were selected. SSR markers were used to analyze genetic diversity of the second generation plus trees, parental analysis and genetic distance estimation. According to the genetic distance among the 2nd-generation plus trees, the second generation breeding populations was structured.【Result】In the progeny tests, the two families were significantly different in growth tratits. Most progenies have a family heritability above the medium level (h2≥0.2) in volume which was suitable for the selection of superior families. Based on this, selection of second generation of breeding population can use the combined individual selection and mass selection. 163 trees were selected from the progeny tests, and the average genetic gain was 21.95%. The genetic diversity of the second generation breeding materials was studied by using 16 pairs of SSR primers. A total of 45 alleles were detected at 16 loci. The mean number of alleles (Na) per locus was 2.7, polymorphism rate was 100%; the mean number of effective alleles (Ne) per locus was 1.54; Shannon diversity index (I) was 0.49 and the average observed heterozygosity (Ho) was 0.32. Using the Coancestry Version 1.0 software to calculate the average second-generation breeding population, the coancestry coefficient was 0.042 and the breeding population status number was 11.9. According to the 16 pairs of SSR primers amplified, second generation breeding population of male parents were analyzed by the CERVUS2.0 software, and the results indicated that the second generation of the 163 tress, male parent could be determined for 57 individuals under the 95% confidence level, and for other 102 individuals under the 80% confidence level. In order to avoid inbreeding in advanced generations, the second generation breeding populations were clustered with the genetic distance index. According to the results of distance clustering, the 163 individuals were divided into 10 sub-lines, numbered from Gui GC2-A-Gui GC2-J. In the establishment of the second generation seed orchard of Masson pine, following strategies was proposed for 2nd-generation seed orchard: a certain number of best clones from each sub-line were chosen to establish 2ndgeneration seed orchard; mating among the sub-lines was adopted in cross breeding of the next generation, so that the overall level of inbreeding in the breeding population can be maintained at a relatively low level. 【Conclusion】According to preliminary results, the second generation breeding population of Masson pine in Guangxi was composed of 163 trees. This population had a high genetic diversity, and a low degree of inbreeding among individuals. The second generation breeding population was clustered by the genetic distance, and the strategies of advanced-generation cross breeding were designed: large scale hybridization was carried out among the sub-lines, but only the small scale hybridization experiment was carried out in the sub-lines. The strategy can be applied to effectively avoid inbreeding, laying a foundation for advanced-generation cross breeding of Masson pine.
Pinusmassoniana; breeding population; SSR; genetic diversity; parentage analysis
10.11707/j.1001-7488.20170107
2015 -12-31;
2016-05-19。
廣西八桂學者專項經費; 國家科技支撐計劃(2015BAD09B0102); “十二五”農村領域國家科技計劃課題子專題(2012BAD01B0202-04)。
S718.46
A
1001-7488(2017)01-0054-08
* 楊章旗為通訊作者。
