利用釀酒酵母表達抗菌肽Lvcrunstin-B
2017-02-17 10:32:28何蘋萍李麗萍陳福艷馬春霞
江西農業學報 2017年2期
黎 銘,何蘋萍,陳 明,李麗萍,王 瑞,黃 婷,陳福艷,馬春霞
(1.廣西壯族自治區水產科學研究院/廣西遺傳育種與健康養殖重點實驗室,廣西 南寧 530021;2.廣西壯族自治區獸醫研究所,廣西 南寧 530001)
利用釀酒酵母表達抗菌肽Lvcrunstin-B
黎 銘1,何蘋萍1,陳 明1,李麗萍1,王 瑞1,黃 婷1,陳福艷1,馬春霞2*
(1.廣西壯族自治區水產科學研究院/廣西遺傳育種與健康養殖重點實驗室,廣西 南寧 530021;2.廣西壯族自治區獸醫研究所,廣西 南寧 530001)
根據凡納濱對蝦抗菌肽Lvcrustin-B的核酸序列,采用PAS(PCR-based Accurate Synthesis)的方法合成Lvcrustin-B基因,通過雙酶切及T4連接酶技術將Lvcrustin-B插入表達載體pYE-GAPα,獲得重組酵母表達載體pYE-GAPα-Lvcrustin-B。pYE-GAPα-Lvcrustin-B經限制性內切酶AVr II線性化后,通過電穿孔法轉化至畢赤酵母細胞GS115,并使用Zeocin進行抗性篩選,從而獲得高拷貝轉化子,篩選出的酵母菌轉化子經擴大培養后使用YPD培養基進行誘導表達。PCR檢測結果表明Lvcrustin-B基因被穩定整合至畢赤酵母染色體。SDS-PAGE電泳及WB實驗顯示,目的重組蛋白Lvcrustin-B可在畢赤酵母中表達,分子量大小為19.4 kD,且表達方式為可分泌性表達,重組菌最佳培養溫度為30 ℃。
抗菌肽;凡納濱對蝦;GS115;蛋白表達
在我國,水產養殖是一個重要的產業,約束其可持續性發展的主要因素是病害問題:各種細菌或者病毒病經常引起魚、蝦大量死亡,從而給養殖業帶來巨大的經濟損失[1-2]。為了預防和控制魚、蝦病害發生,養殖戶不得不大量使用池塘消毒化學藥物或者在飼料中添加各種抗生素藥物[3]。公眾越來越意識到:大量使用化學藥物或者抗生素類藥物會嚴重破壞環境生態以及帶來食品安全問題,因此,業內亟待開發無污染、無殘留、安全有效的新型漁藥用以代替化學或者抗生素類藥物[4]。
抗菌肽(Antimicrobial peptides)又叫抗微生物多肽或肽抗生素,是生物細胞特定基因編碼并由特定外界條件誘導產生的一類多肽[5]。抗菌肽在動、植物體內分布廣泛,是天然免疫防御系統的重要組成部分[5-6]。與抗生素比較,抗菌肽有以下特點:可以抗革蘭氏陽性菌和陰性菌,對寄生蟲、真菌甚至病毒具有抵抗作用;分子量小,容易被吸收,殺菌作用迅速,不易產生耐藥性;抗菌濃度小,濃度單位一般在μmol/L級水平。抗菌肽作為傳統抗生素的替代物正越來越受到人們的關注,是目前國際學術研究的活躍領域之一[7]。
抗菌肽作為飼料添加劑具有較好的應用前景。抗菌肽能耐受飼料制粒時的高溫,特別是利用酵母表達的抗菌肽經高溫濃縮工序,可充分殺滅酵母菌體而不導致抗菌肽失活,產品應用后不出現工程菌擴散而導致的環境生態問題[8]。部分抗菌肽具有抵抗胃蛋白酶和胰蛋白酶的能力,使之不被胃腸道分解而進入機體發揮作用[8]。
抗菌肽在甲殼類動物飼料生產中具有廣闊的應用前景。與脊椎動物不同,甲殼類動物不產生特異性抗體,其抵御病原入侵主要依賴非特異性免疫系統。凡納濱對蝦的非特異性免疫系統重要組分在抵御病原入侵過程中扮演重要角色[9-10]。據報道,凡納濱對蝦基因組含有大量抗病免疫基因,比如凝集素、抗菌肽、溶菌酶、過氧化物酶等,這些基因有的已經被應用于魚蝦抗病藥物的開發[9-10]。
Lvcrustin-B是筆者在凡納濱對蝦中新發現的抗菌肽基因。本研究利用分子生物學方法構建該基因的真核表達載體,并轉化酵母菌進行表達,旨在為Lvcrustin-B的生產應用奠定基礎。
1 材料與方法
1.1 材料
1.1.1 試劑和耗材 pYE-GAPα質粒及TOP10菌株(本實驗室保種),GS115(Life公司),Protein Marker(北京全式金),PVDF膜(美國Millipore公司),X光片(美國柯達公司),ECL顯色液(中國普利萊公司),鼠抗His單抗(北京全式金),兔抗鼠HRP二抗(北京全式金),Acr、Bis、Tris等(Sigma公司),SDS(Amresco公司),Tryptone、Yeast Extract(購自OXOID公司),PCR反應管,0.22 μm無菌濾器和透析袋(購自Millipore公司),Ni2+IDA親和層析膠(北京全式金公司),Agarose,DNA膠純化試劑盒、質粒小提試劑盒(北京全式金公司),限制性內切酶(大連寶生物公司),常規生化試劑(均為國產分析純)。
pYE-GAPα質粒(見圖1)為環狀雙螺旋質粒,空載質粒含3080個堿基,含有1個博萊霉素篩選基因Zeocin,多克隆位點可以插入外源基因,多克隆位點上游含有6個組氨酸編碼序列,因此所表達的蛋白含有由6個組氨酸組成的蛋白標簽。
1.1.2 主要實驗儀器 Allegra 21R臺式高速冷凍離心機(美國BECKMAN公司),臺式高速離心機(德國SORVAL公司),Biologic LP層析系統、Mini Protean II垂直平板電泳系統、Gel Doc2000成像系統、水平電泳系統(美國BIO-RAD公司),PTC-200基因擴增儀(美國MJ Research公司),320-S pH計(美國Mettler Toledo公司),AR5120電子天平(美國AHOM S公司),MultiTemp III恒溫水浴鍋、Hofer ΜV-25紫外透射儀(美國Amersham Pharmacia公司),雪花狀制冰機(日本SANYO公司),JY92-2D超聲波細胞粉碎機(中國新芝科器研究所),蛋白核酸檢測儀(南京大學普陽科學儀器廠),Gene Pulser Xcell電穿孔系統(美國BIO-RAD),超凈工作臺(中國蘇凈集團),NANODROP2000(美國Thermo公司)。
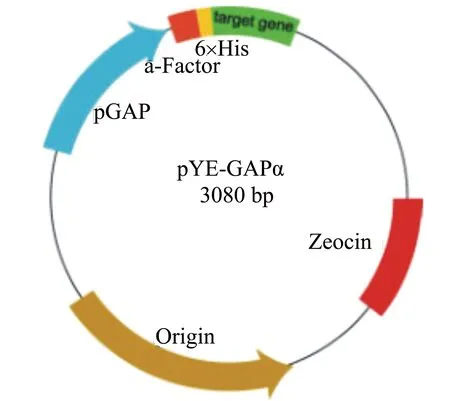
圖1 pYE-GAPα質粒示意圖
1.2 方法
1.2.1 pYE-GAPα-Lvcrustin-B質粒的構建 通過凡納濱對蝦轉錄組測序及分析結果獲得Lvcrustin-B序列,采用PAS(PCR-based Accurate Synthesis)的方法合成基因Lvcrustin-B,使用雙酶切(EcoR I和Xba I)對目的基因片段酶切產生特異接口,然后利用T4連接酶將酶切片段連接至pYE-GAPα載體相應位點,獲得重組質粒pYE-GAPα- Lvcrustin-B。將重組質粒轉化到大腸桿菌感受態細菌TOP10,將重組菌涂布到含有氨芐青霉素(終濃度為100 μg/mL)的LA平板,然后放置在37 ℃的恒溫箱內培養,約18 h后挑取陽性菌落,放入LB液體培養基進行擴大培養。
1.2.2 pYE-GAPα-Lvcrustin-B質粒的提取及鑒定 取擴大培養菌液,放入1.5 mL離心管,以3000 r/min離心,拋棄上清,所得細菌沉淀用于質粒提取,提取方法參照質粒提取試劑盒(北京全式金)操作說明。將提取的質粒分成2份,取其中1份交由華大基因公司進行測序,1份進行雙酶切(EcoR I和Xba I)鑒定,酶切鑒定方法參照限制性內切酶說明書(大連寶生物公司)。
1.2.3 pYE-GAPα-Lvcrustin-B電轉酵母細胞GS115 冰浴電轉杯,將10 μL線性化質粒pYE- GAPα-Lvcrustin-B加入到裝有80 μL畢赤酵母感受態酵母細胞的1.5 mL EP管內,混勻后加入到直徑為0.2 cm的電轉化杯中,將310 μL等滲溶液加入到電轉化杯中,然后把電轉化杯冰浴5 min。電擊條件為:電壓1700 V、時間8 ms、電擊2次。電擊完成后,將1 mL冰上預冷的濃度為1 mol/L的山梨醇溶液加入到電擊轉化杯里,用槍頭輕輕地吹打溶液,使之均勻。把電轉化杯中的全部液體轉移到新的2 mL EP管中,在30 ℃下靜置培養2 h。分別吸取50、100和200 μL線性化質粒pYE-Lvcrustin-B-capsid的混合液,將其涂布于含有100 μg/mL Zeocin抗生素的YPD平板上,在30 ℃下恒溫培養48 h,待平板長出菌落后,用接種環挑取平板上生長的單菌,將它接入到裝有10 YPD液體培養基的試管中(抗生素Zeocin濃度為100 μg/mL),在30 ℃下以180 r/min震蕩過夜培養。
1.2.4 PCR鑒定陽性克隆菌株 挑選6株陽性克隆,分別提取基因組DNA(編號分別為1、2、3、4、5、6),使用引物5′pGAP priming(5′pGAP-F:5′-GTCCCTATTTCAATCAATTGAA-3′)和3′AOX1 priming(3′AOX1:5′-GCAAATGGCATTCTGACATCC-3′)進行PCR擴增。
1.2.5 小試表達 取上述鑒定為陽性的表達菌株(3號菌株),將其接入裝有9 YPD(抗生素Zeocin 200 μg/mL)的試管中,在30 ℃下以220 r/min培養24 h,然后取500 μL接入到50 YPD(抗生素Zeocin 200 μg/mL)液體培養基中,在30 ℃下以220 r/min培養,當培養時間達0、24、48、72、96 h時分別從培養基中取樣,以10000 r/min離心2 min,收集上清液,使用SDS-PAGE電泳檢測結果。
1.2.6 WB鑒定重組蛋白 取鑒定為陽性的表達菌株,加loading buffer,煮沸冷卻后取20 μL上樣。電泳條件:5%濃縮膠,90 V 30 min;10%分離膠,120 V 20 min。轉膜:分別疊放好3層濾紙、SDS-PAGE膠和PVDF膜(PVDF膜事先需要用甲醇活化15 s左右),使用轉膜儀進行濕法轉膜,恒壓100 V 60 min。封閉:用5%脫脂奶粉PBST溶液封閉膜,搖床37 ℃ 2 h,用PBS漂洗5 min。抗體孵育及染色(一抗為Mouse Anti-His mAb)。一抗孵育:抗體1∶500稀釋,4 ℃過夜; PBST在37 ℃漂洗5 min×3。二抗孵育:抗體1∶1500稀釋,37 ℃ 1 h;PBST在37 ℃漂洗5 min×4。曝光2 min,獲取圖像。
2 結果與分析
2.1 pYE-GAPα-Lvcrustin-B質粒的構建結果
基于PAS(PCR-based Accurate Synthesis)的方法合成基因Lvcrustin-B,通過克隆位點EcoR I和Xba I連入酵母表達載體pYE-GAPα,轉入Top10克隆菌株,然后抽提陽性菌株質粒送華大基因公司測序。測序結果如圖2所示,其中下劃線區域為Lvcrustin-B基因區域,灰色區域為酶切位點。

圖2 陽性菌株pYE-GAPα-Lvcrustin-B質粒的測序結果
將測序結果與設計的預期序列進行比對,比對結果顯示兩者100%匹配。截取部分比對序列,如圖3所示。
圖3 部分序列比對結果
2.2 pYE-GAPα-Lvcrustin-B酶切鑒定結果
用限制性內切酶EcoR I和Xba I對抽提得到的表達質粒pYE-GAPα-Lvcrustin-B進行酶切,酶切產物經瓊脂糖凝膠電泳,結果見圖4。
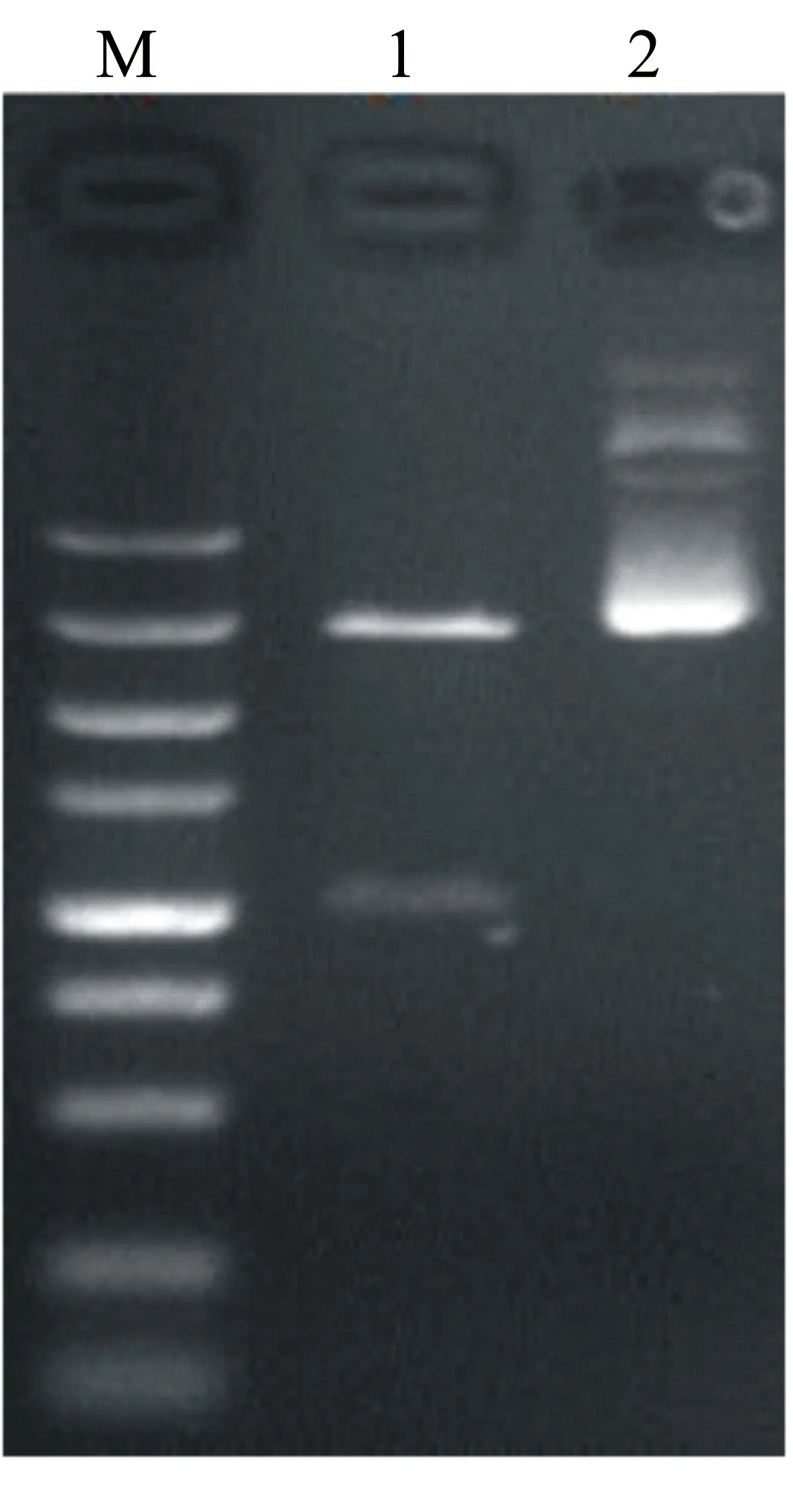
M為核酸分子量標尺Marker,由下至上各條帶的大小分別為100、250、500、750、1000、1500、2000、3000、5000 bp;1為酶切后質粒;2為酶切前質粒。
圖4 pYE-GAPα-Lvcrustin-B質粒酶切鑒定結果
2.3 將pYE-GAPα-Lvcrustin-B電轉化至酵母細胞GS115
實驗結果顯示,培養品經48 h培養后長出多個白色菌落(圖5)。

圖5 pYE-GAPα-Lvcrustin-B電轉化GS115的結果
2.4 PCR鑒定陽性克隆菌
挑選6株陽性克隆,分別提取基因組DNA,使用引物5′pGAP priming和3′AOX1 priming對目的基因進行PCR鑒定,結果顯示均為陽性。PCR鑒定陽性克隆子,預期條帶大小約1.0 kD(圖6)。
2.5 小試表達SDS-PAGE電泳檢測結果
SDS-PAGE電泳檢測結果(圖7)顯示,培養24、48、72、96 h時菌株均有陽性,目的蛋白條帶如箭頭所示。
2.6 Lvcrustin-B蛋白WB鑒定結果
經Western blot驗證,純化的蛋白被抗his抗體識別,條帶大小與預期相符,證明Lvcrustin-B表達成功(圖8)。
3 討論
作為先天防御系統的主要成分,抗菌肽具有廣譜的生物學活性,殺菌迅速,不受傳統抗生素耐藥菌株的影響,與典型的抗生素具有協同作用的特點。隨著對抗菌肽結構與功能的破譯,以及基因工程等批量生產手段的出現,抗菌肽有望成為防治水產養殖主要病害的新型藥物[11]。現有相關研究表明,將抗菌肽制劑添加到水產飼料中不僅可以提高水產動物的抗病免疫力,甚至還可以提高水產品質量,從而緩解水產養殖中細菌耐藥性及水產品抗生素污染等問題。陳冰等在凡納濱對蝦的研究中發現,在一定添加水平范圍內,家蠅抗菌肽能顯著提高凡納濱對蝦的存活率、增重率、特定生長率[12]。王四新等研究發現,在飼料中添加100~150 mg/kg的凡納濱對蝦抗菌肽能顯著提高草魚的生長速度和相對增重率,而添加150 mg/kg天蠶素抗菌肽對草魚存活率和肥滿度的提高效果不明顯[13]。林鑫等(2013年)將飼喂添加抗菌肽飼料的錦鯉感染維氏氣單胞菌,10 d后其累積死亡率顯著降低,充分證實口服抗菌肽可以顯著提高錦鯉的抗病能力[14]。
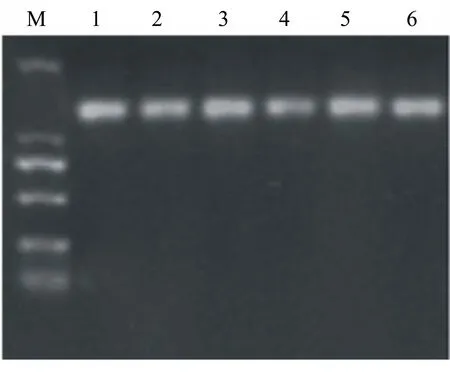
M為DNA Marker,從下到上各條帶的大小分別為100、250、500、750、1000、2000 bp;1~6為陽性克隆子PCR條帶。
圖6 PCR鑒定陽性克隆菌的結果

M: protein marker; 1、2、3、4、5: pGAPZaA-Lvcrustin-B-capsid轉化GS115菌株分別培養0、24、48、72、96 h。
圖7 小試表達SDS-PAGE電泳檢測結果
盡管至今有數千種天然抗菌肽已經被發現,但目前可應用于生產實踐的相關產品比較少,重要原因在于抗菌肽工業化生產技術有待突破。抗菌肽最初是直接從動植物組織中提取獲得的,由于其分子量小、組織中含量低,因此分離提純存在一定的困難。化學合成抗菌肽可以實現規模化生產,但目前使用該方法成本較高,目前國內肽合成價約150元/mg , 而多肽作為藥物廣泛應用的社會可承受價格上線為10美元/mg。小于10個氨基酸殘基的多肽合成較為經濟,大于10 個殘基的多肽合成成本較高[15],同時由于缺乏天然化學修飾,其活性也難以保證。所以通過基因工程在微生物中直接表達抗菌肽基因是最佳方案。本研究利用畢赤酵母GS115成功表達了對蝦抗菌肽Lvcrustin-B,與動植物組織提取法、化學合成方法相比,其操作簡便,生產成本較低,非常適用于未來的大規模發酵生產。
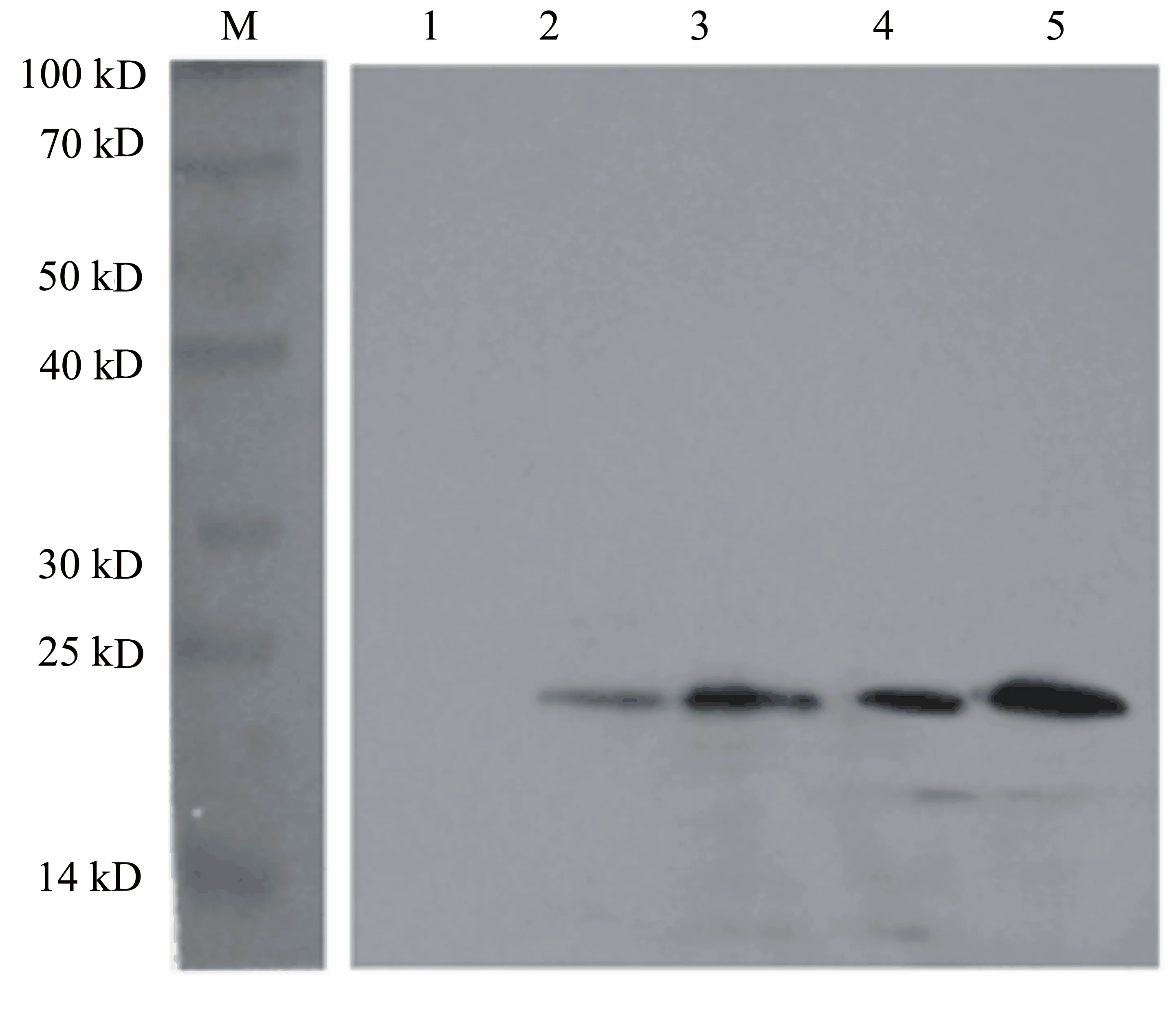
M: protein marker;1:空白對照;2~5:不同濃度的Lvcrustin-B蛋白樣品。
圖8 Lvcrustin-B蛋白WB鑒定結果
利用基因工程方法在微生物中表達抗菌肽有兩種方法,一種是原核表達方法,另外一種是真核表達方法。這兩種方法各有優點:原核表達方法的表達量比較高,但是不具有真核生物的基因表達調控機制和蛋白質翻譯后加工修飾能力,其產物往往形成沒有活性的包涵體,須經過變性、復性等處理才能應用,某些表達菌內毒素含量高,表達產物需要經過嚴格的純化處理;利用真核表達方法在微生物中表達抗菌肽時,由于經過真核細胞內特有的各種化學修飾,因此表達出來的抗菌肽活性較高,利用酵母菌表達還可以避免內毒素的出現,但缺點是表達載體構建較為困難,有些重組蛋白不適合在真核細菌中表達,因此表達量較低[16-17]。本研究利用畢赤酵母GS115表達對蝦抗菌肽Lvcrustin-B,在30 ℃下以220 r/min培養24 h后,取發酵液離心后的上清液進行SDS電泳及Western blot分析,發現從24 h開始就檢測到有重組蛋白的存在,而且隨著培養時間的延長,重組蛋白的含量越來越高,說明利用畢赤酵母GS115表達Lvcrustin-B是可行的,而且分泌表達更有利于表達產物的提純。以往有不少研究利用酵母菌表達重組蛋白,往往需要甲醇的誘導,因此表達產物中可能存在著甲醇的殘留,因此表達產物需要進一步提純去除甲醇成分才能作為藥物或者添加劑使用。本研究采用pYE-GAPα質粒構建重組表達載體,實驗結果表明,該載體轉化的酵母菌不需要甲醇誘導即可表達,更有利于今后表達產物的使用。
[1] 吳淑勤,王亞軍.我國水產養殖病害控制技術現狀與發展趨勢[J].中國水產,2010(8):9-10.
[2] 卓然江,蔡小琴,洪徐鵬,等.南美白對蝦“偷死病”的病因分析及防治措施[J].科學養魚,2015(8):58-59.
[3] 唐江芳.水產藥物殘留的研究現狀[J].南方農業,2007(1):39-41,52.
[4] 潘葳,羅欽,劉文靜,等.水產品與水產飼料中藥物殘留問題的分析及對策[J].福建農業學報,2011(6):1096-1100.
[5] 牛德料,孟日增,劉彩俠.抗菌肽的研究進展[J].江西飼料,2013(3):21-24.
[6] 孫龍.兩種鞘翅目昆蟲抗菌肽的分離純化與活性研究[D].北京:中國林業科學研究院,2012.
[7] 胡夢紅,王有基.傳統抗生素替代物抗菌肽在水產養殖中的應用前景[J].北京水產,2007(1):47-49.
[8] 李莉蓉.以DNA為靶標的抗菌肽設計及抑菌機制研究[D].無錫:江南大學,2013.
[9] 于愛清.蝦蟹類免疫相關基因的研究[D].上海:華東師范大學,2014.
[10] 羅展.凡納濱對蝦兩種C-型凝集素的基因克隆和功能研究[D].北京:中國科學院研究生院海洋研究所,2010.
[11] 李曉華,陶冉,孫杰.對蝦抗菌肽的研究進展及其在水產養殖業的應用前景[J].安徽農業科學,2009(1):167-168.
[12] 陳冰,曹俊明,陳平潔,等.家蠅抗菌肽對凡納濱對蝦生長性能及免疫相關指標的影響[J].中國水產科學,2010(2):258-266.
[13] 王四新,季海峰,劉輝,等.抗菌肽對草魚生長性能的影響[J].飼料研究,2011(4):29-31.
[14] 林鑫,毛述宏,楊陽,等.飼料中添加抗菌肽對錦鯉生長、非特異性免疫力和抗病力的影響[J].動物營養學報,2013(8):1860-1865.
[15] 孔凡紅,仲維霞,崔勇,等.生物抗菌肽畢赤酵母表達系統研究進展[J].中國病原生物學雜志,2009(9):700-702.
[16] 劉曉明,姜寧,張愛忠,等.雜合抗菌肽在畢赤酵母中的表達及其活性測定[J].中國生物工程雜志,2016(2):81-89.
[17] 劉忠淵.利用畢赤酵母系統表達功能蛋白的研究[D].烏魯木齊:新疆大學,2004.
(責任編輯:黃榮華)
Expression ofLitopenaeusvannameiAntimicrobial Peptide Lvcrunstin-B inPichiapastoris
LI Ming1, HE Ping-ping1, CHEN Ming1, LI Li-ping1, WANG Rui1,HUANG Ting1, CHEN Fu-yan1, MA Chun-xia2*
(1. Guangxi Academy of Fishery Sciences/Guangxi Key Laboratory of Aquatic Genetic Breeding and Healthy Aquaculture, Nanning 530021, China; 2. Guangxi Veterinary Research Institute, Nanning 530001, China)
According to the nucleotide sequence of antimicrobial peptide Lvcrunstin-B inLitopenaeusvannamei, the method of PAS (PCR-based Accurate Synthesis) was used for the synthesis of gene Lvcrustin-B. The synthetic gene Lvcrustin-B was doubly digested by EcoR I and Xba I, then it was ligated into the expression vector pYE-GAPα by using T4 ligase method, and the obtained recombinant yeast expression vector was named as pYE-GAPα-Lvcrustin-B. The plasmid pYE-GAPα-Lvcrustin-B was transformed intoPichiapastorisGS115 through electroporation method after linearizing by AVr II method. The transformants were screened according to their resistance to Zeocin, and then high-copy transformants were obtained. After being cultivated in a large-scale, the screened yeast transformants were induced to express with YPD medium. The results of PCR testing showed that the gene Lvcrustin-B had been steadily integrated into the chromosome ofPichiapastoris. The results of SDS-PAGE electrophoresis and Western blot experiment confirmed that the recombinant target protein Lvcrustin-B could express in a secretory manner in the yeast GS115, its molecular weight was 19.4 kD, and the optimum culture temperature for the recombinant yeast was 30 ℃.
Antimicrobial peptides;Litopenaeusvannamei; GS115; Protein expression
2016-08-31
廣西水產畜牧科技推廣應用項目(桂漁牧科201528012);廣西科學基金項目(2015GXNSFAA139067)。
黎銘(1980─),副研究員,主要從事海洋生物研究工作。*通訊作者:馬春霞。
TQ927
A
1001-8581(2017)02-0009-05
