藏北高原草地群落的數量分類與排序
2016-12-28 06:05:59王景升姚帥臣王志凱馮繼廣
生態學報 2016年21期
王景升, 姚帥臣, 普 窮, 王志凱, 馮繼廣
1 中國科學院地理科學與資源研究所生態系統網絡觀測與模擬重點實驗室,北京 100101 2 中國人民大學環境學院,北京 100872 3 西藏大學農牧學院高原生態研究所,林芝 860000
藏北高原草地群落的數量分類與排序
王景升1, 姚帥臣2,*, 普 窮3, 王志凱2, 馮繼廣2
1 中國科學院地理科學與資源研究所生態系統網絡觀測與模擬重點實驗室,北京 100101 2 中國人民大學環境學院,北京 100872 3 西藏大學農牧學院高原生態研究所,林芝 860000
采用TWINSPAN數量分類和DCA、CCA排序的方法,對藏北高原草地29個樣點進行統計分析。結果顯示:(1)TWINSPAN數量分類將藏北高寒草地群落劃分成10種類型。(2)樣點DCA排序第一軸基本反映了水分環境梯度,第二軸基本反映了熱量梯度。(3)TWINSPAN分類所劃分的各群落在DCA排序圖上都有各自的分布范圍和界限,說明DCA排序能較好的反應各優勢群落與其環境資源之間的關系。(4)樣點CCA排序表明,影響群落分布的首要環境因子是水分因子(年均降水量)和空間因子(經度),其次是熱量因子(年均溫度),CCA排序進一步闡明了群落分布決定于水分和溫度等環境因子,并間接驗證了TWINSPAN的分類結果。(5)物種CCA排序和TWINSPAN分類結果表明:植物群落中物種的分布格局與植物群落類型的分布格局存在一定的相似性。
藏北高原;草地;數量分類;排序;群落
青藏高原是我國乃至亞洲的江河源,向來有世界屋脊和第三極之稱,是我國及東亞的重要生態安全屏障區[1]。作為青藏高原的主體部分,藏北高原是青藏高原腹地分布最為廣泛、生物區系最為獨特、生態功能最為重要的草地生態系統,不僅是亞洲中部高寒環境中最為典型的自然生態系統之一,在世界高寒地區亦具有代表性[2]。然而,隨全球氣候變化和人類活動加劇,近年來藏北高原高寒草地面臨局部地區嚴重退化問題[3]。目前,對高寒草地的研究多集中于草地退化和沙化的治理、土壤養分流失、生物量動態以及對氣候變化的響應等方面[4- 9]。而對大尺度范圍內高寒草地群落的內部分類、空間分布和生態因子變化特征以及它們之間的相關性的系統研究較少,群落自身的生態特點、內在規律等尚不十分明確。數量分類和排序分析方法能夠系統研究高寒草地群落的內部分類、群叢特征,并能科學地揭示植被—環境間的生態關系[10- 14]。因此,本文應用雙向指示種分類法(Two Way Indicator Species Analysis,TWINSPAN)、典范對應分析(Canonical Correspondence Analysis,CCA)和去趨勢對應分析(Detrended Canonical Correspondence,DCA)等方法,定量分析了環境因子對高寒草地植物群落分布和物種組成的影響。以期闡明高寒草地的地理分布格局并為合理保護利用當地草地資源提供參考。
1 研究區概況
藏北高原又稱“羌塘高原”,位于西藏自治區北部,南自岡底斯—念青唐古拉山脈,北至昆侖山脈,東迄東經91°左右的內外流水系之分水嶺,西止于國界,南北寬達760 km,東西長約1200 km(北緯29°53′—36°32′,東經78°41′—92°16′),面積59.5萬km2,約占西藏自治區土地面積的一半。藏北高原擁有遼闊的天然草地資源,各類天然草地面積共達4800萬hm2,占藏北高原土地面積的81%和西藏自治區天然草地面積的59%,為西藏主要牧區[15]。藏北高原是青藏高原主體,海拔高度在4600—5100 m之間;藏北高原寒冷干燥,大部分地區年平均氣溫低于0 ℃,最暖月均溫不及14 ℃。年均降水量50—400 mm,且80%以上集中在6—8月,年蒸發量大于1500 mm,年平均干燥度指數1.6—20.0,年均風速高于3.0 m/s[16- 19]。天然草地類型主要包括高寒草甸、低草型高寒草原、高寒荒漠草原三大類。其中,高寒草甸的優勢種為高山嵩草(Kobresiapygmaea)、紫花針茅(Stipapurpurea)等,總蓋度40%—80%;高寒草原的優勢種為紫花針茅、青藏苔草(Carexmoorcroftii)、矮火絨草(Leontopodiumnanum)、伊凡苔草(Carexivanovae)等,總蓋度15%—50%;高寒荒漠草原的優勢種為沙生針茅(Stipaglareosa)、鋪散亞菊(Ajaniakhartensis)、紫花針茅等,總蓋度10%—20%[20]。
2 研究方法
2.1 群落調查
采用實驗生態學中典型樣方調查法,在藏北羌塘高原29個樣點上各設置1條100 m樣線,沿樣線設置5個1 m×1 m樣方,樣方間隔20 m,詳細記錄樣方中各物種的高度、蓋度和頻度、群落平均高、總蓋度等參數。樣方合計145個。
2.2 氣象數據
以藏北羌塘高原及其周邊270個氣象站長期監測數據為本底,使用Anualspline插值軟件進行插值,并經過中科院地理資源所那曲生態站2009年在那曲-阿里的樣帶上布設的10套HOBO氣象監測系統實測數據驗證后,通過GIS平臺系統提取獲得。主要包括多年平均氣溫、降雨量、濕潤度等。
2.3 數據處理
計算樣方內各草本植物的重要值,其計算公式如下[21]:草本植物重要值=(相對蓋度 + 相對高度)×100/2野外調查于2014年8月生物量高峰期完成,根據調查數據建立兩個矩陣:由29個樣點145個樣方中74種草本的重要值組成的植被矩陣;由29個樣點和5個環境參數組成的環境屬性矩陣。應用Wintwins 2.3中雙向指示種分析(Two Way Indicator Species Ananlysis,TWINSPAN)進行植被數量分類,應用Canoco for Windows 4.5中的典范對應分析(Canonical Correspondence Analysis,CCA)和去趨勢對應分析(Detrended Canonical Correspondence,DCA)進行樣方和環境參數的排序。
3 結果與分析
3.1 樣方的TWINSPAN數量分類
將29個樣點進行TWINSPAN分類并依據植物群落分類和命名原則,結合調查結果的生態分析和群落生境特征的指示物種或優勢物種命名群落類型,本區植物群落可分為10類(如圖1)。
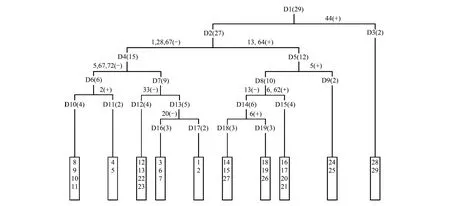
圖1 樣方TWINSPAN分類樹狀圖Fig.1 Dendrogram of the TWINSPAN classification of 29 plots指示物種 1: 矮火絨草;2:矮金蓮花;5:半球齒緣草;6:半臥狗娃花;13:臭棘豆;20:短穗兔耳草Lagotis brachystachya;28:高山嵩草;33:華扁穗草;44:鋪散亞菊;62:西藏燥原薺;64:小墊黃芪;67:伊凡苔草;72:紫花地丁
Ⅰ紫花針茅(Stipapurpurea) + 微藥羊茅(Festucanitidula) + 伊凡苔草(Carexivanovae)群落
包括樣點8—11,共4個樣點,分布在班戈縣。該群落分布于海拔4620—4645 m,年均溫-0.2—0 ℃,年降水量432—466 mm。群落平均蓋度20%,優勢種為紫花針茅、微藥羊茅和伊凡苔草,主要伴生種有矮火絨草、高寒早熟禾(Poakoelzii)、紫花地丁(Stipapurpurea)、圓齒褶龍膽(Gentianacrenulatotruncata)等。
Ⅱ紫花針茅(Stipapurpurea) + 伊凡苔草(Carexivanovae)群落
包括樣點4、5,共2個樣點,分布在安多縣。該群落分布于海拔4590—4630 m,年均溫- 1.5℃,年降水量480—520 mm。群落平均蓋度32%,優勢種為紫花針茅和伊凡苔草,主要伴生種有矮金蓮花(Trolliusfarreri)、矮火絨草、短葉羊茅(Festucabrachyphylla)、粗壯嵩草(Kobresiarobusta)、多枝黃芪(Astragaluspolycladus)、高山大戟(Euphorbiastracheyi)、穗三毛(Trisetumspicatum)等。
Ⅲ紫花針茅(Stipapurpurea) + 矮火絨草(Leontopodiumnanum)群落
該群落包括樣點12、13、22、23,共4個樣點,分布在雙湖縣和尼瑪縣。海拔4750—5000 m,年均溫-1.8℃,年降水量273—384 mm。群落平均蓋度13%,優勢種為紫花針茅和矮火絨草,主要伴生種有二裂委陵菜(Potentillabifurca),華扁穗草(Blysmussinocompressus)和青藏苔草等。
Ⅳ高山嵩草(Kobresiapygmaea)群落,包括樣點3、6、7,共3個樣點,主要分布在安多縣。該群落分布于海拔4685—4727 m,年均溫-1.8—-1.7℃,氣候較為濕潤,年降水量517—528 mm。群落平均蓋度達到73%,優勢種為高山嵩草,主要伴生種有矮火絨草、短穗兔耳草(Lagotisbrachystachya)、高寒早熟禾、高山唐松草(Thalictrumalpinum)、西藏風毛菊(Saussureatibetica)、楔葉委陵菜(Potentillacuneata)、紫花針茅等。
Ⅴ高山嵩草(Kobresiapygmaea) + 紫花針茅(Stipapurpurea)群落
包括樣點1、2,共2個樣點,分布于那曲縣。分布海拔4450—4460 m,年均溫0.8℃,年降水量高達570 mm。該群落平均蓋度51%,優勢種為高山嵩草和紫花針茅,主要伴生種有二裂委陵菜、木根香青(Anaphalisxylorhiza)、西藏風毛菊、無心菜(Anaphalisxylorhiza)和楔葉委陵菜等。
Ⅵ紫花針茅(Stipapurpurea)群落
包括樣點14、15、27,該群落分布于雙湖縣和改則縣。分布海拔4600—4860 m,年均溫度較低,氣候寒冷,降水較少。群落平均蓋度17%,優勢種為紫花針茅,主要伴生種有臭棘豆(Oxytropischiliophylla)、二裂委陵菜、華扁穗草、小墊黃芪(Astragaluspuivinalis)等。
Ⅶ紫花針茅(Stipapurpurea) + 華扁穗草(Blysmussinocompressus) + 臭棘豆(Oxytropischiliophylla)群落
該群落包括樣點18、19、26,分布于尼瑪縣和改則縣。分布海拔4550—4600 m,年均溫較高,降水較少。群落總蓋度19%—33%,優勢種為紫花針茅、華扁穗草和臭棘豆,主要伴生種有半臥狗娃花(Heteropappussemiprostratus)、多枝黃芪、二裂委陵菜和小墊黃芪等。
Ⅷ紫花針茅(Stipapurpurea) + 半臥狗娃花(Heteropappussemiprostratus) + 小墊黃芪(Astragaluspuivinalis)群落
包括樣點16、17、20、21,共4個樣點,分布于尼瑪縣。該群落分布于海拔4520—4620 m,年均溫較高,年降水量310—330 mm。群落平均蓋度17%,優勢種為紫花針茅、半臥狗娃花和小墊黃芪,主要伴生種有西藏燥原芥(Ptilotricumwageri)、楔葉委陵菜等。
Ⅸ青藏苔草(Carexmoorcroftii) + 紫花針茅(Stipapurpurea) + 藏布紅景天(Rhodiolasmithii)群落
包括樣點24、25,分布于措勤縣。該群落分布海拔4755—4815 m,年均溫-0.8 ℃,年均降水量230—270 mm。群落平均蓋度22%,優勢種為青藏苔草、紫花針茅和藏布紅景天,主要伴生種有半球齒緣草(Eritrichiumhemisphaericum)、垂穗披堿草(Elymusnuta)、短軸嵩草(Kobresiavidua)、裂葉獨活(Heracleummillefolium)、沙蒿(Artemisiadesertorum)、無心菜、西藏風毛菊、小墊黃芪等。
Ⅹ沙生針茅(Stipaglareosa) + 紫花針茅(Stipapurpurea) + 鋪散亞菊(Ajaniakhartensis)群落
該群落包括樣點28、29,分布于革吉縣。群落分布海拔4430—4490 m,年均溫較高,降水較少,氣候非常干旱。群落平均蓋度較低,只有11%,物種多為耐干旱植物,優勢種為沙生針茅、紫花針茅和鋪散亞菊,主要伴生種有西藏燥原芥、固沙草(Orinusthoroldii)等。
3.2 樣點DCA排序分析
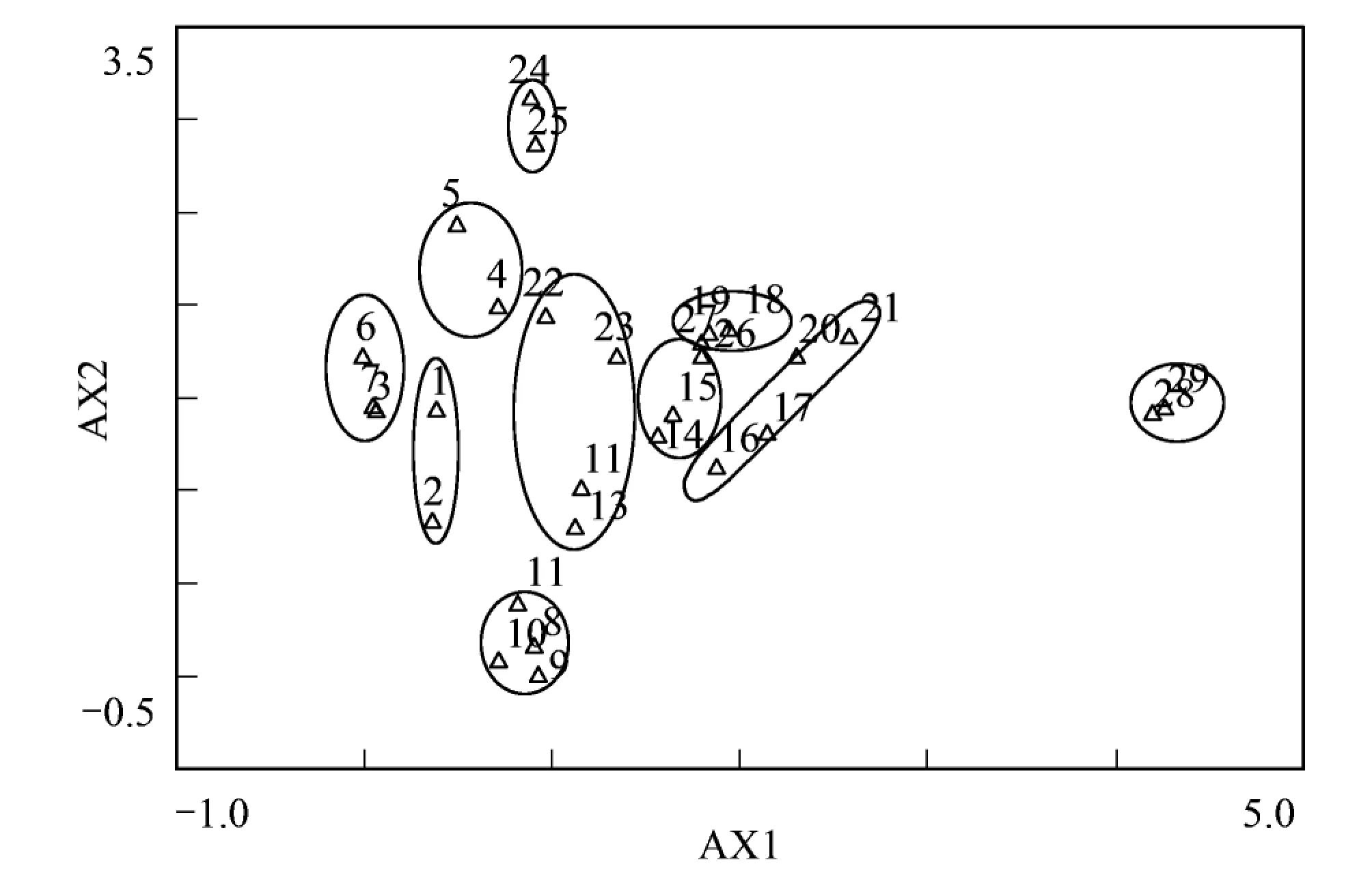
圖2 樣點DCA二維排序圖Fig.2 Two-dimensional DCA ordination diagram of 29 plots
采用DCA對研究區29個樣點進行排序分析,結果如圖2,得到前4個排序軸的特征值分別為0.681、0.459、0.196、0.119。第一二軸特征值較大,說明這兩個排序軸包含的生態信息量多,具有更重要的生態意義。從二維排序圖可以看出,TWINSPAN分類結果所得的群落類型在DCA二維排序圖上均有自己的分布范圍和界限,說明DCA排序較好的反應了各樣方之間和樣方與環境之間的關系。從排序軸看,第一軸基本反映了水分環境梯度,從左到右,環境條件由較為濕潤過渡到干旱。第二軸反映了溫度梯度的變化,即第二軸從下往上,年均溫逐漸降低。樣點的DCA排序圖綜合反映了植物群落和生境的空間梯度,與TWINSPAN的分類結果完全匹配。
3.3 樣點CCA排序
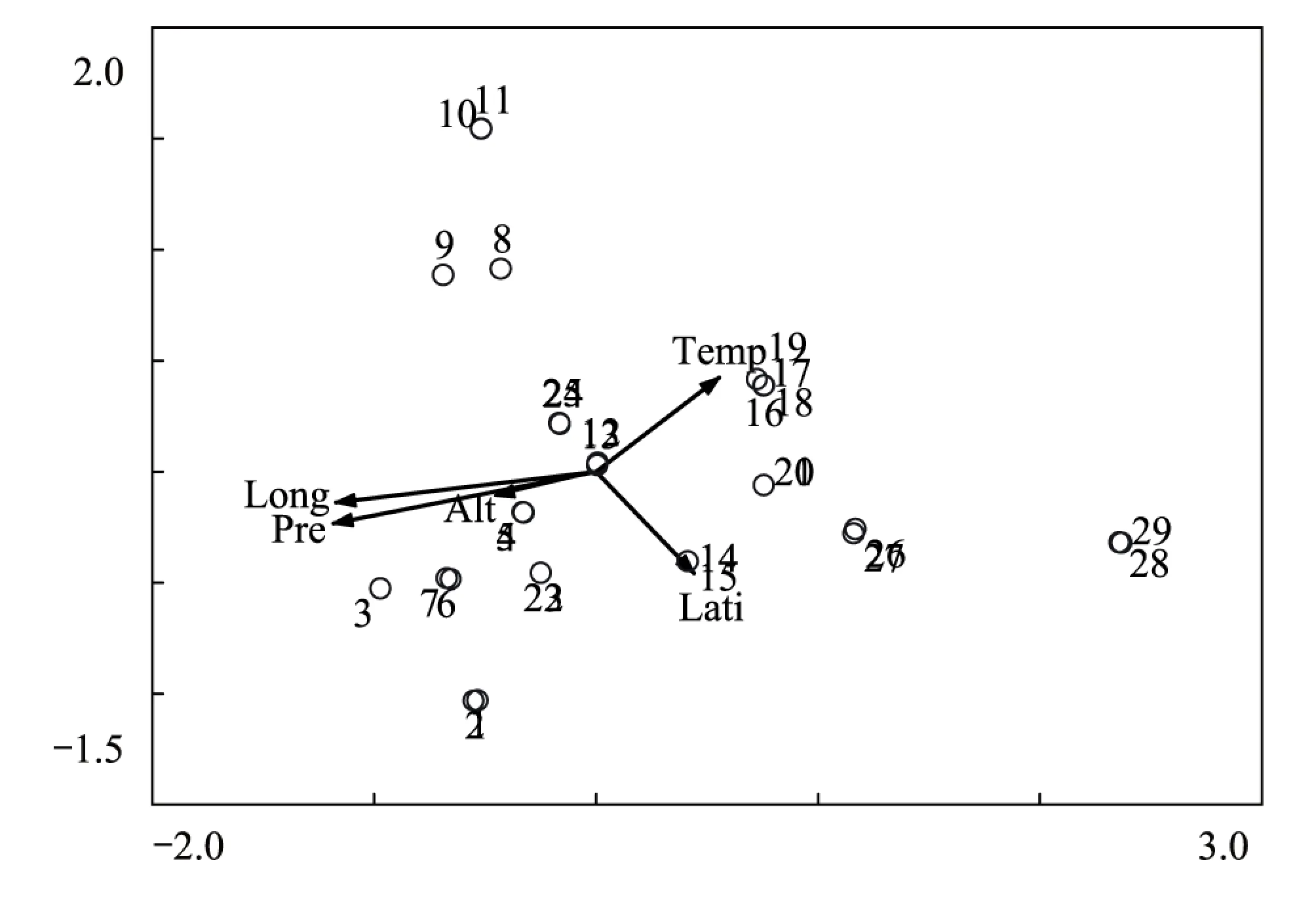
圖3 樣點CCA排序圖Fig.3 CCA ordination diagram of 29 plots
典范對應分析(Canonical Correspondence Analysis,CCA),是一種基于單峰模型的排序方法,樣方排序與對象排序對應分析,而且在排序過程中結合多個環境因子,因此可以把樣方、對象與環境因子的排序結果表示在同一排序圖上[10]。為進一步探討群落分布與環境因子之間的關系,對29個樣點和5個環境因子進行CCA排序,結果如圖3。排序圖中箭頭表示環境因子,箭頭連線的長度代表著相應環境因子與研究對象分析相關程度的大小,越長代表其對所研究對象的分布影響越大;箭頭連線與排序軸夾角余弦值代表其與排序軸的相關性大小。沿排序圖第一軸,隨著年降水量的逐漸減少,群落從排序圖最左側生境相對濕潤的高山嵩草群落和高山嵩草 + 紫花針茅群落過渡到中間較為干旱的紫花針茅群落、紫花針茅 + 半臥狗娃花 + 小墊黃芪群落、青藏苔草 + 紫花針茅 + 藏布紅景天群落,再到最右側干旱的沙生針茅 + 紫花針茅 + 鋪散亞菊群落。沿排序軸第二軸從上向下,隨著緯度增加,溫度降低,群落由紫花針茅 + 微藥羊茅 + 伊凡苔草群落過渡到高山嵩草群落。從排序結果可以看出,影響群落分布的首要環境因子是水分因子(年均降水量)和空間因子(經度),其次是熱量因子(年均溫度)。即是說,首先,水分條件的差異決定著研究區的植被被分為三種類型:相對濕潤的高寒草甸、較為干旱的高寒草原和干旱高寒荒漠草原,其次,在水分條件相差不大的情況下,由于溫度的不同,3種植被類型又被分為不同的群落。此外,排序圖中可以看出TWINSPAN劃分的十類群落在CCA排序中基本都有各自的分布范圍和界限,這說明TWINSPAN的分類結果和樣方CCA排序是一致的。
3.4 物種的CCA排序
采用CCA對74個物種進行數量分析,結果如圖4所示。由圖可以看出,物種的分布格局與圖3樣點DCA中樣點的分布格局相似。物種CCA第一軸反映了各物種分布生境的水分條件,沙生針茅、固沙草、西藏燥原芥等耐旱植物分布在排序圖最右側較為干旱的區域,高山嵩草、無莖黃鵪菜、短穗兔耳草、西藏風毛菊等分布生境較為濕潤的植物則分布在排序圖左側。半臥狗娃花、斑唇馬先蒿、冰川棘豆、梭羅草等與溫度相關性比較大,均分布在相對溫暖的區域,而紫花針茅、伊凡苔草、二裂委陵菜、華扁穗草、矮火絨草等分布在中央,說明這幾種植物生態位適應性較強,分布較為廣泛。結合分類結果和樣點DCA排序圖,可以看到植物群落中物種的分布格局與植物群落類型的分布格局有很大的相關性。
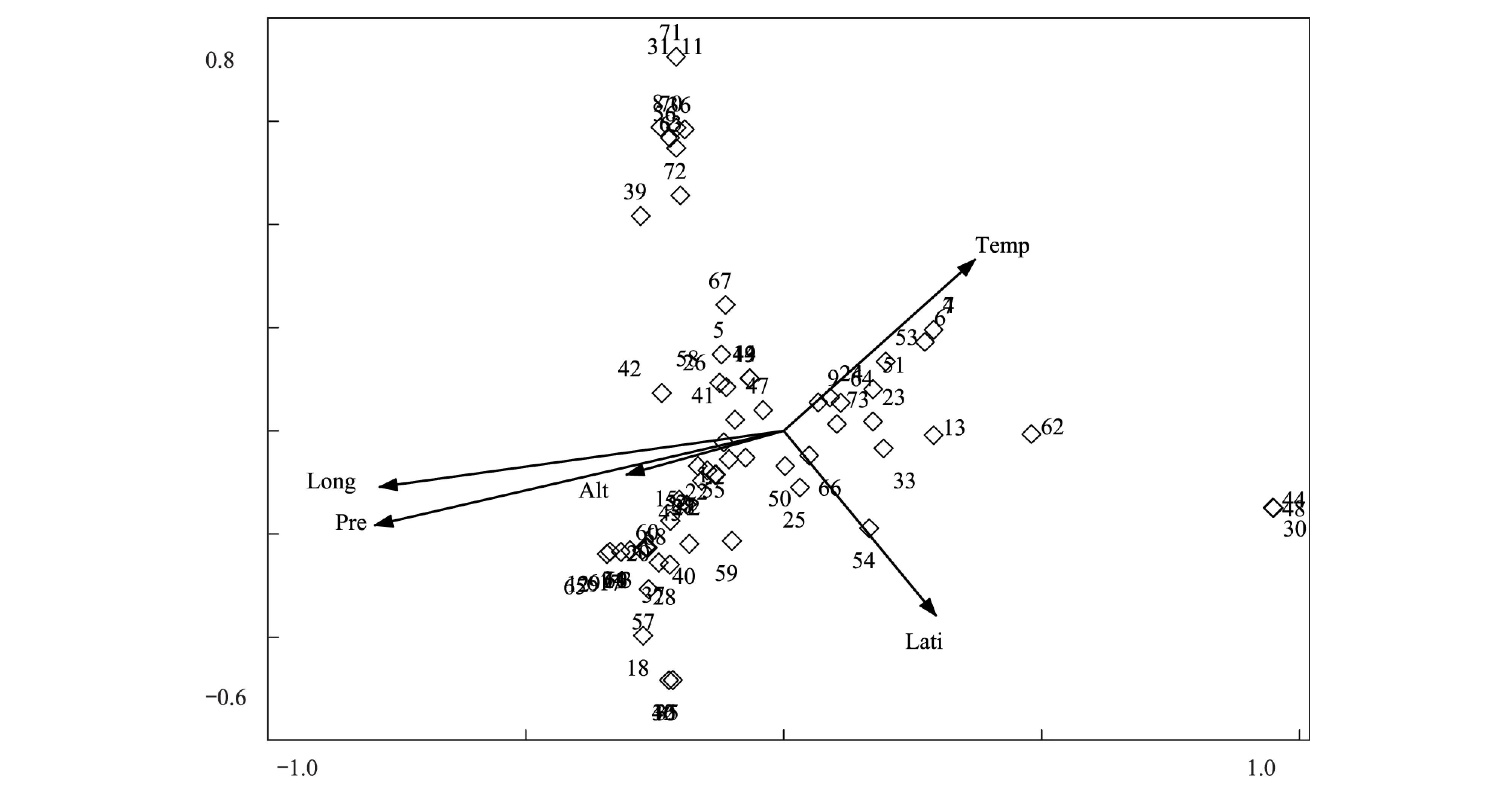
圖4 物種CCA排序圖Fig.4 CCA ordination diagram of 74 species3: 白蒿 Leontopodium dedekensii; 4: 斑唇馬先蒿 Pedicularis longiflora; 7: 冰川棘豆 Oxytropis glacialis; 8: 藏菠蘿花 Incarvillea compacta; 9: 藏布紅景天; 10: 藏豆 Stracheya tibetica; 11: 藏沙蒿 Artemisia wellbyi; 12: 策勒蒲公英 Taraxacum qirae; 14: 垂穗披堿草; 15: 粗壯嵩草; 16: 單子麻黃 Ephedra monosperma; 17: 墊狀點地梅 Androsace tapete; 18: 釘柱委陵菜 Potentilla saundersiana; 19: 獨一味 Lamiophlomis rotate; 21: 短葉羊茅; 22: 短軸嵩草;23: 多枝黃芪; 24: 二花棘豆 Oxytropis biflora; 25: 二裂委陵菜;26: 高寒早熟禾; 27: 高山大戟; 29: 高山唐松草;30: 固沙草; 31: 禾葉點地梅 Androsace graminifolia; 32: 黑包風毛菊 Saussurea melanotrica; 34: 金露梅 Potentilla fruticosa; 35: 蕨麻委陵菜 Potentilla anserine; 36: 賴草 Leymus secalinus; 37: 蘭石草 Lancea tibetica; 38: 藍翠雀花 Delphinium caeruleum; 39: 狼毒 Euphorbia fischeriana; 40: 藜 Chenopodium album; 41: 裂葉獨活; 42: 木根香青; 43: 螃蟹甲 Phlomis younghusbandii; 45: 青藏苔草; 46: 弱小火絨草; 47: 沙蒿; 48: 沙生針茅; 49: 少花棘豆 Oxytropis pauciflora; 50: 四數獐牙菜 Swertia tetraptera; 51: 碎米薺 Cardamine hirsute; 52: 穗三毛 Trisetum spicatum; 53: 梭羅草 Roegneria thoroldiana; 54: 團墊黃芪 Astragalus arnoldii; 55: 微孔草 Microula sikkimensis; 56: 微藥羊茅; 57: 無莖黃鵪菜 Youngia simulatrix;58: 無心菜; 59: 西伯利亞蓼 Polygonum sibiricum; 60: 西藏風毛菊; 61: 西藏棱子芹 Pleurospermum hookeri; 63: 纖桿蒿 Artemisia demissa; 65: 小葉棘豆 Oxytropis microphylla; 66: 楔葉委陵菜; 68 羽葉釘柱委陵菜 Potentilla saundersiana; 69: 羽毛委陵菜 Potentilla plumose; 70: 圓齒褶龍膽; 71: 脹果棘豆 Oxytropis stracheyana; 73: 紫花針茅; 74: 紫菀 Aster tataricus
4 結論與討論
TWINSPAN數量分類將藏北草原29個樣點劃分為10種群落類型,即紫花針茅 + 微藥羊茅 + 伊凡苔草群落;紫花針茅 + 伊凡苔草群落;紫花針茅 + 矮火絨草群落;高山嵩草群落;高山嵩草 + 紫花針茅群落;紫花針茅群落;紫花針茅 + 華扁穗草 + 臭棘豆群落;紫花針茅 + 半臥狗娃花 + 小墊黃芪群落;青藏苔草 + 紫花針茅 + 藏布紅景天群落;沙生針茅 + 紫花針茅 + 鋪散亞菊群落。采用DCA對29個樣點進行排序分析,各群落在排序圖上都有各自的分布范圍和界限,可見DCA排序能較好的反應各樣方之間和樣方與環境之間的關系,同時TWINSPAN的分類結果也在排序圖上得到較好的印證。樣方DCA排序第一軸基本反映了水分環境梯度,第二軸基本反映了溫度梯度。
樣點CCA排序結果表明,影響群落分布的首要環境因子是水分因子(年均降水量)和空間因子(經度),其次是熱量因子(年均溫度),CCA排序進一步闡明了群落分布于環境之間的關系,并再次驗證了TWINSPAN的分類結果。物種CCA的排序結果顯示,植物群落中物種的分布格局與植物群落類型的分布格局存在一定的相似性。
對于DCA排序軸的生態解釋,不同學者的研究結果有所不同,有些學者認為,DCA第一軸反映水分環境梯度,第二軸反映熱量環境梯度[21- 24]。部分學者則認為DCA第一軸反映(海拔)熱量環境梯度,第二軸反映水分環境梯度[25- 28]。還有學者認為,DCA排序第一軸反映了群落海拔高度的梯度變化,第二軸反映了群落坡度的梯度變化或者坡向的梯度變化[29-30]。Burke等人的研究則表明,在一定程度上,DCA第一軸反映土壤性質和養分梯度,而在小環境中,水分條件和干擾程度決定著植被的空間分布[31]。由此,我們可以看出,DCA排序軸的環境解釋在不同的研究地點和尺度,有可能得出不同的研究結論。在降雨相對豐富地區,可能熱量因素更大程度上影響了植被的組成和空間分布,而在西部黃土高原等干旱地區,水分條件可能成為影響植被空間分布的主導因素,而在干旱荒漠地區,水分條件相差不大的地區,土壤養分含量也有可能成為影響植被分布的關鍵因子[24]。藏北高原地處大陸腹地,平均海拔4600m以上,年均溫相差不大,年均降水量較少,且由于自身高原地貌的影響,降水隨著經度的減小而逐漸減少。在較低的降水水平下,降水的減少會降低土壤中的有效水分、改變土壤的理化性質、影響植物對養分的吸收、運輸和利用,進而影響群落中各類型植物的比例和重要性。物種的CCA排序結果顯示:在環境較為濕潤的高寒草甸中,群落中濕生植物占絕對優勢,而在干旱的荒漠草原,旱生植物則占有絕對優勢。這充分說明水分條件在很大程度上影響著物種的分布以及群落的結構組成,因此可以認為水分條件是影響藏北草原植被空間分布的主導因素。
綜上,本文采用TWINSPAN分類結合DCA和CCA排序的研究方法,對藏北草地的數量分析取得了良好的分類結果,并初步揭示了高寒草地植物群落及環境因子的分布格局。可為藏北草原的保護和利用,以及相關的植被群落研究提供理論依據。
致謝:本文實驗樣地以中國科學院地理科學與資源研究所藏北高寒草甸生態系統研究站(那曲站)為依托,樣地調查及制圖過程中得到了站內工作人員和武建雙博士、田源和曲蕓瑩碩士的幫助,石培禮研究員和付剛老師對文章寫作給予幫助,特此致謝。
[1] 孫鴻烈, 鄭度, 姚檀棟, 張鐿鋰. 青藏高原國家生態安全屏障保護與建設. 地理學報, 2012, 67(1): 3- 12.
[2] 張榮祖, 鄭度, 楊勤業. 西藏自然地理. 北京: 科學出版社, 1982.
[3] 王景升, 張憲洲, 趙玉萍, 秦泗國, 武建雙. 羌塘高原高寒草地生態系統生產力動態. 應用生態學報, 2010, 21(6): 1400- 1404.
[4] 武建雙, 李曉佳, 沈振西, 張憲洲, 石培禮, 余成群, 王景升, 周宇庭. 藏北高寒草地樣帶物種多樣性沿降水梯度的分布格局. 草業學報, 2012, 21(3): 17- 25.
[5] 張生楹, 陳谷, 陳建綱, 張德罡. 藏北高原高寒草地土壤有效養分含量研究. 草原與草坪, 2012, 32(3): 6- 10.
[6] 蔡曉布, 張永青, 邵偉. 藏北高寒草原草地退化及其驅動力分析. 土壤, 2007, 39(6): 855- 858.
[7] 趙景學, 陳曉鵬, 曲廣鵬, 多吉頓珠, 尚占環. 藏北高寒植被地上生物量與土壤環境因子的關系. 中國草地學報, 2011, 33(1): 59- 64.
[8] 楊秀海, 卓嘎, 羅布. 藏北高原氣候變化與植被生長狀況. 草業科學, 2011, 28(4): 626- 630.
[9] 付剛, 周宇庭, 沈振西, 張憲洲, 武建雙. 藏北高原高寒草甸地上生物量與氣候因子的關系. 中國草地學報, 2011, 33(4): 31- 36.
[10] 張金屯. 數量生態學. 北京: 科學出版社, 2004.
[11] Oldeland J, Dorigo W, Lieckfeld L, Lucieer A, Jürgens N. Combining vegetation indices, constrained ordination and fuzzy classification for mapping semi-natural vegetation units from hyperspectral imagery. Remote Sensing of Environment, 2010, 114(6): 1155- 1166.
[12] Ermakov N, Makhatkov I. Classification and ordination of north boreal light-coniferous forests of the West Siberian Plain. Plant Biosystems, 2011, 145(S1): 199- 207.
[13] Dale E E Jr, Ware S, Waitman B. Ordination and classification of bottomland forests in the lower Mississippi alluvial plain. Castanea, 2007, 72(2): 105- 115.
[14] Ahmad S S, Quratulann. Vegetation classification in ayubia national park, pakistan using ordination methods. Pakistan Journal of Botany, 2011, 43(5): 2315- 2321.
[15] 胡自治. 青藏高原的草業發展與生態環境. 北京: 中國藏學出版社, 2000: 1- 5, 19- 22, 100- 124.
[16] 毛飛, 盧志光, 鄭凌云, 張佳華. 近40年那曲地區日照時數和風速變化特征. 氣象, 2006, 32(9): 77- 83.
[17] 毛飛, 盧志光, 張佳華, 鄭凌云. 近40年那曲地區氣候特征分析. 高原氣象, 2007, 26(4): 708- 715.
[18] 王景升, 張憲洲, 趙玉萍, 沈振西, 石培禮, 余成群. 藏北羌塘高原氣候變化的時空格局. 資源科學, 2008, 30(12): 1852- 1859.
[19] 毛飛, 盧志光, 張佳華, 侯英雨. 近20年藏北地區AVHRR NDVI與氣候因子的關系. 生態學報, 2007, 27(8): 3198- 3205.
[20] 李明森. 藏北高原草地資源合理利用. 自然資源學報, 2000, 15(4): 335- 339.
[21] 馬克平, 劉玉明. 生物群落多樣性的測度方法: I α多樣性的測度方法(下). 生物多樣性, 1994, 2(4): 231- 239.
[22] 張峰, 張金屯, 張峰. 歷山自然保護區豬尾溝森林群落植被格局及環境解釋. 生態學報, 2003, 23(3): 421- 427.
[23] 張桂蓮, 張金屯, 程林美. 山西南部山地白羊草群落的數量分類和排序. 草業學報, 2003, 12(3): 63- 69.
[24] 董林水, 張旭東, 周金星, 宋愛云. 晉西黃土丘陵與土石山區交錯地帶灌木種的數量分類與排序. 生態學報, 2007, 27(7): 3072- 3080.
[25] 岳鵬鵬, 盧學峰, 葉潤蓉, 周玉碧, 趙建忠, 彭敏. 長江源區紫花針茅草原群落數量分類與排序. 廣西植物, 2014, 34(5): 635- 641.
[26] 史作民, 劉世榮, 程瑞梅, 蔣有緒. 河南寶天曼植物群落數量分類與排序. 林業科學, 2000, 36(6): 20- 27.
[27] 劉秋鋒, 康慕誼, 劉全儒. 中條山混溝地區森林喬木種的數量分類與環境解釋. 植物生態學報, 2006, 30(3): 383- 391.
[28] 岳鵬鵬, 孫菁, 盧學峰, 葉潤蓉, 周玉碧, 楊仕兵, 彭敏. 青海高原紫花針茅草原群落數量分類與排序. 西北植物學報, 2014, 34(5): 1047- 1054.
[29] 何惠琴, 李紹才, 孫海龍, 劉世昌, 熊文蘭. 錦屏水電站植被數量分類與排序. 生態學報, 2008, 28(8): 3706- 3712.
[30] 陳云, 王海亮, 韓軍旺, 韋博良, 賈宏汝, 葉永忠, 袁志良. 小秦嶺森林群落數量分類、排序及多樣性垂直格局. 生態學報, 2014, 34(8): 2068- 2075.
[31] Burke A. Classification and ordination of plant communities of the Naukluft Mountains, Namibia. Journal of Vegetation Science, 2001, 12(1): 53- 60.
Quantitative classification and ordination of grassland communities on the Northern Tibetan Plateau
WANG Jingsheng1, YAO Shuaichen2,*, PU Qiong3, WANG Zhikai2, FENG Jiguang2
1KeyLaboratoryofEcosystemNetworkObservationandModeling,InstituteofGeographicSciencesandNaturalResourcesResearch,ChineseAcademyofSciences,Beijing100101,china2SchoolofEnvironmentandNaturalResources,RenminUniversityofChina,Beijing100872,China3TibetInstituteofPlateauEcology,TibetAgriculturalandAnimalHusbandryCollege,Linzhi860000,China
The relationship between vegetation and the environment is one of the fundamental questions in understanding plant species composition and community distribution along an environmental gradient. Understanding vegetation-environment correlations may help predict possible shifts in plant communities in response to climate and land use changes. There are 48 million hm2of natural grasslands on the Northern Tibetan Plateau, accounting for 59% of the total grassland area in the Tibet Autonomous Region. However, these alpine grasslands are threatened by global climate change and intense human activities in recent decades, and are now widely degraded. Quantitative classification and ordination are important methods in examining the internal classification of plant communities, characteristics of their associations, and in revealing the ecological relationships between vegetation and the environment. TWINSPAN classification and DCA,CCA ordination were used to conduct a classification and ordination of the plant communities of 29 plots in the northern Tibetan grassland. Our results indicate that: (1) Northern Tibetan alpine meadows are classified into 10 associations:Stipapurpurea+Festucanitidula+Carexivanovae;Stipapurpurea+Carexivanovae;Stipapurpurea+Leontopodiumnanum;Kobresiapygmaea;Kobresiapygmaea+Stipapurpurea;Stipapurpurea;Stipapurpurea+Blysmussinocompressus+Oxytropis;Stipapurpurea+Heteropappussemiprostratus+Astragaluspuivinalis;Carexmoorcroftii+Stipapurpurea+Rhodiolasmithii; andStipaglareosa+Stipapurpurea+Ajaniakhartensis; (2) The first axis of DCA basically reflected a moisture gradient, and the second axis indicated a thermal gradient; (3) Each community divided by TWINSPAN classification had its specific distribution and boundaries on the DCA ordination diagram, which indicated that DCA ordination can explain the relationships between the plots and environment; (4) CCA plots indicated that the primary environmental factors controlling the distribution of communities were rainfall (average annual rainfall) along longitude, followed by temperature (average annual temperature); CCA sorting further elaborated the relationship between plant community distribution and the environment, and indirectly supported the results of the TWINSPAN classification; (5) Quantitative analysis of 74 species by CCA indicated that the distribution pattern of species and plots in DCA was similar. The first axis of species CCA reflected water conditions of species distribution. Taken together, we conclude that the distribution patterns of plant species communities closely mirror each other. In general, the present study provides an ecological interpretation of the distribution of plant species communities along an environmental gradient on the Northern Tibetan Plateau.
Northern Tibetan Plateau; grassland; quantitative classification; ordination; community
國家科技支撐課題資助項目(2011BAC09B03);科技部973課題資助項目(2013CB956300);國家自然科學基金資助項目(41271067)
2015- 04- 24;
日期:2016- 03- 03
10.5846/stxb201504240848
*通訊作者Corresponding author.E-mail: yaoshuaichen@yeah.net
王景升, 姚帥臣, 普窮, 王志凱, 馮繼廣.藏北高原草地群落的數量分類與排序.生態學報,2016,36(21):6889- 6896.
Wang J S, Yao S C, Pu Q, Wang Z K, Feng J G.Quantitative classification and ordination of grassland communities on the Northern Tibetan Plateau.Acta Ecologica Sinica,2016,36(21):6889- 6896.
猜你喜歡
兒童時代·幸福寶寶(2022年12期)2022-12-09 11:24:14
中學生數理化·七年級數學人教版(2022年11期)2022-02-14 07:14:12
數學小靈通(1-2年級)(2021年4期)2021-06-09 06:25:56
大眾健康(2021年6期)2021-06-08 19:30:06
科普童話·學霸日記(2020年1期)2020-05-08 16:45:11
中學生數理化·七年級數學人教版(2019年4期)2019-05-20 10:06:32
小天使·一年級語數英綜合(2019年2期)2019-01-10 11:57:30
中學生數理化·七年級數學人教版(2018年6期)2018-06-26 08:36:06
兒童繪本(2018年5期)2018-04-12 16:45:32
初中生世界·七年級(2017年9期)2017-10-13 22:27:46
