Peroxiredoxin-1基因沉默對轉化生長因子β1促進肺成纖維細胞Ⅰ、Ⅲ型膠原合成的影響*
2016-12-19 01:18:43劉寶欣劉英宇魏中秋梁婷婷范玉磊
重慶醫學 2016年29期
關鍵詞:水平
劉寶欣, 劉英宇,魏中秋, 梁婷婷,范玉磊,楊 方,孫 影△
(1.華北理工大學附屬唐山市工人醫院呼吸內科,河北唐山 063000;2.華北理工大學基礎醫學院病理學系,河北唐山 063000)
?
·技術與方法·
Peroxiredoxin-1基因沉默對轉化生長因子β1促進肺成纖維細胞Ⅰ、Ⅲ型膠原合成的影響*
劉寶欣1, 劉英宇1,魏中秋2, 梁婷婷2,范玉磊2,楊 方2,孫 影2△
(1.華北理工大學附屬唐山市工人醫院呼吸內科,河北唐山 063000;2.華北理工大學基礎醫學院病理學系,河北唐山 063000)
目的 探討Peroxiredoxin-1(Prx-1)基因沉默對轉化生長因子β1(TGF-β1)促進肺成纖維細胞合成Ⅰ和Ⅲ型膠原的影響及其作用機制。方法 體外培養肺成纖維細胞分為4組:對照組(0.4%血清),TGF-β1組(5μg/L),TGF-β1+陰性轉染組(5μg/L TGF-β1+陰性對照siRNA),TGF-β1+Prx-1siRNA轉染組(5μg/L TGF-β1+Prx-1siRNA)。利用脂質體Lipo2000將陰性對照siRNA和Prx-1siRNA分別轉染到TGF-β1+陰性轉染組和TGF-β1+ Prx-1siRNA轉染組的肺成纖維細胞中,轉染48h后用于后續實驗。實時熒光定量PCR(RT-PCR)檢測轉染后各組的Prx-1mRNA表達水平,蛋白免疫印跡法(Western blot)檢測磷酸化Akt(p-Akt)及Ⅰ和Ⅲ型膠原蛋白水平,2,7-二氯熒光素二乙酸檢測活性氧(ROS)水平。結果 (1) Prx-1siRNA轉染肺成纖維細胞后,TGF-β1+ Prx-1siRNA轉染組的Prx-1mRNA表達明顯降低。(2) 與對照組比較,TGF-β1組Ⅰ和Ⅲ 型膠原、ROS及p-Akt蛋白水平均明顯增加(0.34±0.06vs. 0.58±0.06、0.42±0.05vs. 0.56±0.06、2988±379vs. 4315±580和0.29±0.05vs. 0.66±0.07,差異有統計學意義(P<0.05)。與TGF-β1組比較,TGF-β1+陰性轉染組Ⅰ和Ⅲ型膠原、ROS及p-Akt水平無明顯變化(P>0.05),但TGF-β1+Prx-1siRNA轉染組Ⅰ和Ⅲ 型膠原、ROS及p-Akt蛋白水平均進一步增高(0.58±0.06vs. 0.79±0.09、0.56±0.06vs. 0.77±0.08、4315±580vs. 5841±782和0.66±0.07vs. 0.93±0.15,差異有統計學意義(P<0.05)。結論 TGF-β1能夠誘導肺成纖維細胞生成ROS,并由此促進Akt的激活和肺成纖維細胞合成Ⅰ和Ⅲ型膠原;而沉默Prx-1基因可激活ROS/Akt通路,從而有助于TGF-β1促進肺成纖維細胞合成膠原。
活性氧;轉化生長因子-β1;膠原;Peroxiredoxin-1
矽肺是長期吸入二氧化硅粉塵而引起的一種職業病,其主要病理變化是肺組織彌漫性纖維化,Ⅰ和Ⅲ型膠原的表達增多是其中重要原因之一[1]。轉化生長因子β1(transforming growth factro-β1,TGF-β1)在矽肺肺組織中表達增高,具有促纖維化作用。筆者前期研究顯示,TGF-β1可通過上調活性氧(reactive oxygen specie,ROS)激活C-Jun蛋白激酶(JNK)細胞內信號傳導通路,從而促進肺成纖維細胞合成Ⅰ和Ⅲ型膠原[2-3]。最近Lu等[4]報道博來霉素通過激活ROS/Akt通路促進肺組織纖維化。Peroxiredoxin 家族是一類新發現的過氧化物酶,其中,Peroxiredoxin 1(Prx-1)在哺乳動物中廣泛表達,可快速有效地清除細胞內產生的ROS,并抑制ROS介導的細胞內傳導通路激活[5]。但在矽肺纖維化過程中,目前并不清楚ROS/Akt通路是否介導了TGF-β1促肺成纖維細胞Ⅰ、Ⅲ 型膠原合成增加的作用,及Prx-1是否通過抑制ROS/Akt通路激活來抑制TGF-β1的促纖維化作用。本項目利用TGF-β1刺激正常及轉染Prx-1siRNA的肺成纖維細胞,檢測Ⅰ和Ⅲ型膠原、ROS及Akt磷酸化的變化,探討Prx-1對ROS/Akt通路介導的TGF-β1促纖維化作用的影響。
1 材料與方法
1.1 材料 人胚胎肺成纖維細胞MRC-5購自中國科學院細胞庫;TGF-β1購自美國Peprotech公司;總Akt(T-Akt)及磷酸化Akt(p-Akt)抗體購自美國Cell Signaling公司;Prx-1抗體購自美國Abcam產品公司;Ⅰ、Ⅲ型膠原抗體和GAPDH抗體購自武漢博士德生物有限公司;Prx-1siRNA及Prx-1基因上、下游引物由上海吉瑪公司合成;M-MLV逆轉錄試劑盒、SYBR+Tap混合劑購自美國Invitrogen公司;活性氧檢測試劑盒購自江蘇南通碧云天生物技術公司。凝膠圖像分析儀購自美國Bio-RAD公司。
1.2 方法
1.2.1 肺成纖維細胞的培養 肺成纖維細胞在5%血清水平的改良杜氏伊格爾培養基(DMEM),5% CO2,37℃條件下常規體外培養。
1.2.2 實驗分組 MRC-5細胞分為4組:對照組(0.4%血清),TGF-β1組(5μg/L),TGF-β1+陰性轉染組(5μg/L TGF-β1+陰性對照siRNA),TGF-β1+Prx-1siRNA轉染組(5μg/L TGF-β1+Prx-1siRNA),每組樣本數為5。本研究共設計3個不同靶基因位點的特異性Prx-1siRNA,分別是Prx-1siRNA-209、Prx-1siRNA-289、Prx-1siRNA-453,選擇抑制作用最明顯的Prx-1siRNA用于實驗。利用脂質體Lipo2000將陰性對照siRNA、Prx-1siRNA分別轉染到TGF-β1+陰性轉染組和TGF-β1+Prx-1siRNA轉染組,轉染48h后各組細胞用0.4%血清培養12h,使其同步化。
1.2.3 實時熒光定量PCR(RT-PCR)檢測Prx-1siRNA轉染后Prx-1mRNA水平 Trizol法提取對照組、TGF-β1+陰性轉染組和TGF-β1+Prx-1siRNA轉染組的細胞總RNA。逆轉錄后取行RT-PCR,擴增體系如下:SYRB+Taq混合劑10μL,Prx-1或甘油醛-3-磷酸脫氫酶(GAPDH)上、下游引物各0.5μL(Prx-1的上游引物為5′-CCC CAC GGA GAT CAT TGC TT-3′,Prx-1的下游引物為5′-CGA GAT GCC TTC ATC AGC CT-3′;GAPDH的上游引物為5′-ATG AAT GGG CAG CCG TTA GG-3′,GAPDH的下游引物為5′-TGG ATT TGC CAT GGG TGG A-3′,cDNA 1μL,雙蒸水8μL;循環參數為:95℃預變性30s;95℃ 5s,60℃ 30s,72℃ 30s,循環40次。
1.2.4 2,7-二氯熒光素二乙酸檢測細胞內ROS水平 2,7-二氯熒光素二乙酸(無熒光)進入細胞后被酯酶分解,生成還原型二氯熒光素(無熒光),還原型二氯熒光素不但無法穿過細胞膜,還可被細胞內的ROS氧化生成氧化型二氯熒光素,二氯熒光素可發出熒光,因此熒光量可間接反映細胞內ROS水平。本研究中,細胞同步化后,與TGF-β1共孵育20min,收集制備單細胞懸液。加入2,7-二氯熒光素二乙酸(終濃度為10μmol/L),37℃孵育20min,洗滌后計數1×104個細胞,熒光酶標儀(激發波長488nm,發射波長525nm)檢測各組的熒光強度。
1.2.5 蛋白免疫印跡法(Western blot)檢測Ⅰ、Ⅲ型膠原和Akt水平 各組細胞同步化后,與TGF-β1共孵育45min (用于檢測p-Akt)和48h(用于檢測Ⅰ和Ⅲ型膠原蛋白),磷酸鹽緩沖液(PBS)洗滌后細胞裂解液裂解細胞30min,收集細胞,離心后收集上清液,-70℃保存。利用考馬斯亮藍法測定細胞總蛋白濃度,并以每孔20μg總蛋白量進行電泳。轉膜后,經5%牛血清清蛋白室溫孵育1h,T-Akt、p-Akt、Ⅰ型膠原、Ⅲ型膠原和GAPDH抗體(均1∶1000稀釋) 4℃孵育過夜,Ⅱ抗(均1∶3000稀釋)室溫孵育2h,BCIP/NBT(1∶50稀釋)顯色3min。Image J軟件掃描蛋白表達條帶并進行定量分析。p-Akt與T-Akt的比值為p-Akt的表達值,Ⅰ或Ⅲ型膠原與GAPDH的比值為Ⅰ或Ⅲ型膠原的表達值。
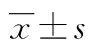
2 結 果
2.1 Prx-1siRNA轉染對肺成纖維細胞Prx-1mRNA表達的影響 RT-PCR結果顯示,對照組和TGF-β1+陰性轉染組的Prx-1mRNA的表達水平差異無統計學意義(P>0.05)。與對照組和TGF-β1+陰性轉染組相比,轉染的3個特異性Prx-1siRNA均可不同程度地降低肺成纖維細胞Prx-1mRNA水平,其中,以Prx-1siRNA-453抑制作用最明顯,遂將Prx-1siRNA-453用于后續實驗,見表1。

表1 Prx-1siRNA轉染對MRC-5細胞Prx-1mRNA表達的影響
2.2 Ⅰ、Ⅲ型膠原蛋白表達 Western blot結果顯示,對照組Ⅰ和Ⅲ型膠原的表達值分別為0.34±0.06和0.42±0.05,TGF-β1組Ⅰ和Ⅲ型膠原的表達值分別為0.59±0.07和0.57±0.07,TGF-β1組Ⅰ和Ⅲ型膠原的表達明顯高于對照組,差異有統計學意義(P<0.01)。TGF-β1+陰性轉染組Ⅰ和Ⅲ型膠原的表達值分別為0.52±0.07和0.58±0.09,與TGF-β1組比較,差異無統計學意義(P>0.05)。但TGF-β1+Prx-1siRNA轉染組Ⅰ和Ⅲ型膠原的表達值分別為0.79±0.09和0.77±0.08,明顯高于TGF-β1組的表達,差異有統計學意義(P<0.05),見圖1。

1:對照組;2:TGF-β1組;3:TGF-β1+陰性轉染組;4:TGF-β1+Prx-1siRNA轉染組。
2.3 ROS水平 對照組、TGF-β1組、TGF-β1+陰性轉染組和TGF-β1+Prx-1siRNA轉染組的熒光強度分別為2988±379、4315±580、4850±572、5841±782。TGF-β1組的熒光強度顯著高于對照組,差異有統計學意義(P<0.01)。TGF-β1+陰性轉染組和TGF-β1組的熒光強度差異無統計學意義(P>0.05),但TGF-β1+Prx-1siRNA轉染組的熒光強度明顯高于TGF-β1組,差異有統計學意義(P<0.05)。
2.4 T-Akt及p-Akt的表達 各組T-Akt水平差異無統計學意義(P>0.05)。對照組、TGF-β1組、TGF-β1+陰性轉染組和TGF-β1+Prx-1siRNA轉染組的p-Akt表達值分別為0.29±0.05、0.65±0.07、0.70±0.10和0.93±0.15。TGF-β1組p-Akt表達明顯高于對照組,差異有統計學意義(P<0.01)。TGF-β1+陰性轉染組和TGF-β1組p-Akt表達差異無統計學意義(P>0.05),但TGF-β1+Prx-1siRNA轉染組p-Akt表達明顯高于TGF-β1組,差異有統計學意義(P<0.01,圖2)。

1:對照組;2:TGF-β1組;3:TGF-β1+陰性轉染組;4:TGF-β1+Prx-1siRNA轉染組。
3 討 論
ROS是在人體有氧代謝過程中產生的一類含有活性氧功能基團的化合物,包括氧自由基、過氧化物和激發態氧等。正常時ROS的生成與清除保持動態平衡,濃度很低,參與機體免疫過程、抵抗損傷等,對機體有保護作用。但病理狀態下ROS生成增多或清除減少,濃度增高,通過攻擊細胞脂質、蛋白質和DNA引起細胞損傷[6]。另外,ROS還參與細胞內信號轉導通路,作為TGF-β1等多種細胞因子的第二信使,調節ERK1/2、JNK等信號轉導途徑,介導細胞增殖和膠原合成等[2,7]。Akt又稱蛋白激酶B,其激酶活性區的氨基酸組成與蛋白激酶A和蛋白激酶C具有高度的同源性。Akt屬于一種絲氨酸/蘇氨酸激酶,當其第308位蘇氨酸和第473位絲氨酸發生磷酸化時,該激酶即被激活,并從細胞膜轉移到細胞內,通過調控下游靶蛋白,進而調節細胞生物學活動。最近研究顯示,ROS激活的Akt途徑在膠原的合成過程中發揮重要作用。如Liu等[8]報道在卵清蛋白誘導的哮喘和氣道重塑過程中,高良姜素通過抑制ROS/Akt途徑抑制膠原蛋白的合成,說明ROS/Akt通路介導了卵清蛋白誘導的膠原合成增加。Pérez de Obanos等[9]也報道谷胱甘肽通過降低ROS水平抑制了亮氨酸激活的Akt途徑,從而減少肝星形細胞合成Ⅰ型膠原。
利用Western blot,本研究發現TGF-β1刺激肺成纖維細胞48h后,Ⅰ和Ⅲ型膠原表達水平較對照組分別增高了1.71和1.33倍,說明TGF-β1可誘導肺成纖維細胞合成Ⅰ和Ⅲ型膠原;同時TGF-β1組的ROS及p-Akt水平也明顯增高,提示ROS/Akt信號通路的激活在TGF-β1誘導的肺成纖維細胞合成Ⅰ和Ⅲ型膠原過程中發揮著重要作用。
Prx-1是一種新型過氧化物酶,具有強大的抗氧化能力,其含有的過氧化半胱氨酸和可溶性半胱氨酸是消除H2O2的主要功能基團。首先過氧化半胱氨酸與H2O2作用,失去質子,轉變為半胱氨酸次磺酸。然后半胱氨酸次磺酸再通過二硫鍵與可溶性半胱氨酸縮合,并在細胞內的二硫鍵還原酶作用下還原為氧化半胱氨酸,回到初始狀態。這個循環過程可有效降低H2O2水平[10]。現已發現在多種疾病中Prx-1均可通過降低ROS來調節細胞的生物學活性,如Prx-1通過降低ROS抑制內皮細胞線粒體的損傷及線粒體介導的細胞凋亡[11]。Madrigal-Matute等[12]最新研究發現Prx-1與NADPH氧化酶在人動脈粥樣硬化斑塊巨噬細胞中表達位置相同,表達水平也呈正相關,認為不但Prx-1可在ROS產生的第一時間原位將其清除,而且對ROS敏感度高,是氧化應激的“感受器”。另外,更重要的是Prx-1還可抑制ROS介導的細胞內信號傳導通路激活,如在β-拉帕醌(一種抗癌藥)誘導的人子宮頸癌細胞凋亡過程中,凋亡信號調節激酶(ASK1)可通過激活JNK途徑促進細胞凋亡,而Prx-1可通過促進TRX與ASK1的結合而抑制ASK1的活性和細胞凋亡[13]。
利用體內外實驗,筆者前期研究發現Prx-1在矽肺大鼠肺組織中表達增高, Prx-1可以通過抑制ROS來抑制JNK和ERK1/2通路激活,抑制TGF-β1誘導的肺成纖維細胞增殖及膠原合成增加,從而抑制矽肺纖維化[3,7,14]。本實驗利用Prx-1siRNA轉染肺成纖維細胞使肺成纖維細胞Prx-1基因沉默,發現沉默Prx-1基因后可明顯上調TGF-β1刺激的ROS水平,同時p-Akt 和Ⅰ、Ⅲ型膠原合成增強,說明沉默Prx-1基因可激活ROS/Akt信號傳導通路,從而有助于TGF-β1促進肺成纖維細胞合成膠原。也進一步證實TGF-β1誘導的肺成纖維細胞合成Ⅰ和Ⅲ型膠原過程中,ROS/Akt信號傳導通路的激活具有重要的促進作用。
[1]Xu H,Yang F,Sun Y,et al.A new antifibrotic target of Ac-SDKP:inhibition of myofibroblast differentiation in rat lung with silicosis[J].PLoS One,2012,7(7):e40301.
[2]趙立雙,魏中秋,楊方,等.活性氧參與TGF-β1的促肺成纖維細胞增殖和膠原合成作用[J].中華勞動衛生與職業病雜志,2015,33(1):15-19.
[3]孫影,魏中秋,胡亞萍,等.新型過氧化物酶Peroxiredoxin-1對大鼠肺成纖維細胞JNK介導的Ⅰ和Ⅲ型膠原合成的影響[J].解剖學雜志,2014,37(5):590-593.
[4]Lu Y,Azad N,Wang L,et al.Phosphatidylinositol-3-kinase/Akt regulates bleomycin-induced fibroblast proliferation and collagen production[J].Am J Respir Cell Mol Biol,2010,42(4):432-441.
[5]Bae JY,Ahn SJ,Han W,et al.Peroxiredoxin Ⅰ and Ⅱ inhibit H2O2-induced cell death in MCF-7cell lines[J].J Cell Biochem,2007,101(4):1038-1045.
[6]李丹,李蓓,石亞楠,等.心力衰竭與活性氧關系研究進展[J].中華實用診斷與治療雜志,2013,27(1):8-10.
[7]孫影,劉寶欣,魏中秋,等.ERK1/2通路在Peroxiredoxin-1抑制TGF-β1誘導的肺成纖維細胞增殖中的作用[J].廣東醫學,2015,36(14):2139-2141.
[8]Liu YN,Zha WJ,Ma Y,et al.Galangin attenuates airway remodelling by inhibiting TGF-β1-mediated ROS Generation and MAPK/Akt phosphorylation in asthma[J].Sci Rep,2015,5(6):11758.
[9]Pérez De Obanos MP,López-Zabalza MJ,Arriazu E,et al.Reactive oxygen species (ROS) mediate the effects of leucine on translation regulation and type Ⅰ collagen production in hepatic stellate cells[J].Biochim Biophys Acta,2007,1773(11):1681-1688.
[10]Rhee SG,Woo HA,Kil IS,et al.Peroxiredoxin functions as a peroxidase and a regulator and sensor of local peroxides[J].J Biol Chem,2012,287(7):4403-4410.
[11]Fiuza B,Subelzú N,Calcerrada P,et al.Impact of SIN-1-derived peroxynitrite flux on endothelial cell redox homeostasis and bioenergetics:protective role of diphenyl diselenide via induction of peroxiredoxins[J].Free Radic Res,2015,49(2):122-132.
[12]Madrigal-Matute J,Fernandez-Garcia CE,Blanco-Colio LM,et al.Thioredoxin-1/peroxiredoxin-1as sensors of oxidative stress mediated by NADPH oxidase activity in atherosclerosis[J].Free Radic Biol Med,2015,86(17):352-361.
[13]He T,Banach-Latapy A,Vernis L,et al.Peroxiredoxin 1 knockdown potentiates β-lapachone cytotoxicity through modulation of reactive oxygen species and mitogen-activated protein kinase signals[J].Carcinogenesis,2013,34(4):760-769.
[14]劉佳麒,鄭素琴,桑銀洲,等.染矽塵大鼠肺組織過氧化物還原酶Ⅰ蛋白表達的情況[J].中華勞動衛生職業病雜志,2013,31(7):531-533.
Effect of silencing peroxiredoxin-1on the expressions of collagen type Ⅰ and Ⅲ in TGF-β1induced pulmonary fibroblasts*
Liu Baoxin1,Liu Yingyu1,Wei Zhongqiu2,Liang Tingting2,Fan Yulei2,Yang Fang2,Sun Ying2△
(1.Department of Respiratory Medicine,Tangshan Works Hospital Affiliated to North China University ofScience and Technology,Tangshan,Hebei 063000,China;2.Department of Pathology,Primary MedicineCollege,North China University of Science and Technology,Tangshan,Hebei 063000,China)
Objective To investigate the effect of silencing peroxiredoxin-1(Prx-1) on the expressions of collagen type Ⅰ and Ⅲ in TGF-β1-induced pulmonary fibroblasts and the possible mechanism.Methods Cultured pulmonary fibroblasts were randomly divided into four groups:control group(0.4% serum),TGF-β1group(5μg/L),TGF-β1+negative transfection group(TGF-β1+scramble siRNA) and TGF-β1+Prx-1siRNA transfection group(TGF-β1+Prx-1siRNA).The negative control siRNA and siRNA Prx-1were transfected into the lung fibroblast cells transfected with TGF-β1+negative transfection group and TGF-β1+Prx-1siRNA transfection group by liposome Lipo2000respectively.After 48h,the cells were was used for subsequent experiments.Real-time PCR was used to evaluate Prx-1mRNA.Western blot was employed to detect the expressions of collagen type Ⅰ and Ⅲ,phosphorylated Akt(p-Akt),and total Akt.Reactive oxygen species (ROS) were measured by DCFH-DA.Results (1)After transfection of siRNA Prx-1into the lung fibroblasts,Prx-1mRNA expression was significantly decreased in the TGF-β1+ Prx-1siRNA group.(2) Compared with control group,expressions of collagen type Ⅰ and Ⅲ,ROS and p-Akt in TGF-β1group were all increased 0.34±0.06vs. 0.58±0.06,0.42±0.05vs. 0.56±0.06,2988±379vs. 4315±580,0.29±0.05vs. 0.66±0.07,(P<0.01).There were no differences in the levels of collagen,ROS and p-Akt between TGF-β1group and TGF-β1+negative transfection group(P>0.05).However,the levels of collagen type Ⅰ and Ⅲ,ROS and p-Akt in TGF-β1+Prx-1siRNA transfection group were further higher than TGF-β1group 0.58±0.06vs. 0.79±0.09,0.56±0.06vs. 0.77±0.08,4315±580vs. 5841±782and 0.66±0.07vs. 0.93±0.15,(P<0.05).Conclusion TGF-β1induces pulmonary fibroblasts to generate ROS,which contributes to Akt activation and collagen type Ⅰ and Ⅲ synthesis;these changes become more obvious with the treatment of Prx-1siRNA.
reactive oxygen species;transforming growth factro-β1;collagen;Peroxiredoxin-1
10.3969/j.issn.1671-8348.2016.29.022
國家自然科學基金資助項目(81072254);唐山市科學技術研究與發展基金資助項目(14130275B)。作者簡介:劉寶欣(1974-),副主任醫師,本科,主要從事肺癌研究。△
R563.9
A
1671-8348(2016)29-4099-04
2016-03-02
2016-04-19)
猜你喜歡
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
火花(2019年12期)2019-12-26 01:00:28
人大建設(2019年6期)2019-10-08 08:55:48
人大建設(2019年12期)2019-05-21 02:55:32
雜文月刊(2018年21期)2019-01-05 05:55:28
人大建設(2017年6期)2017-09-26 11:50:44
學苑創造·A版(2015年11期)2016-01-14 09:03:27
俄羅斯問題研究(2012年1期)2012-03-25 09:54:45
中國火炬(2010年12期)2010-07-25 13:26:22
中國火炬(2010年8期)2010-07-25 11:34:30
