MARCH家族蛋白在免疫、生殖等方面的研究進展*
2016-12-19 05:19:50王美姣綜述于廷和審校
重慶醫學 2016年29期
王美姣 綜述,于廷和 審校
(重慶醫科大學附屬第二醫院婦產科重點實驗室 400010)
?
·綜 述·
MARCH家族蛋白在免疫、生殖等方面的研究進展*
王美姣 綜述,于廷和△審校
(重慶醫科大學附屬第二醫院婦產科重點實驗室 400010)
MARCH;E3泛素連接酶;免疫調節;精子
膜相關RING-CH(membrane-associated RING-CH,MARCH)家族是近年發現的一類屬于E3泛素連接酶的環指結構域家族,迄今已發現的MARCH家族成員有11種,分別是MARCH1~11。MARCH家族的成員分布廣泛,參與多種細胞功能,如免疫調節,蛋白質量控制和膜轉運,內質網相關降解,內體蛋白質運輸,線粒體動力學平衡和精子發生調控等[1-9]。MARCH家族蛋白相關領域的研究正日益受到重視,因此,本文從MARCH家族蛋白的分類、生物學結構、表達及在免疫、生殖等生物學功能方面就MARCH家族最新研究進展作一綜述,旨在為下一步的功能研究和臨床研究提供依據。
1 MARCH家族成員的生物學結構
MARCH家族蛋白結構相對保守,大多數MARCH家族蛋白具有類似結構:一個N端的環指結構域,又稱RING結構域和零個、兩個或多個C端的跨膜結構域(圖1)。RING結構域是此家族具有E3泛素連接酶作用的重要因素。E3泛素連接酶的活性依賴于環指結構域,并通過其與E2泛素結合酶相連。這個環指結構域具有特殊的C3HC4結構,由1個3條鏈組成的β折疊片層、1個α螺旋和兩個大環構成。每一環指結構域連有兩個鋅離子,RING結構域通過與這兩個鋅離子螯合穩定其結構。MARCH家族的11個成員中有9個成員(MARCH1、MARCH2、MARCH3、MARCH4、MARCH5、MARCH6、MARCH8、MARCH9和MARCH11)含有疏水跨膜結構域,并定位于質膜和細胞器膜。典型的MARCH蛋白特征是2個跨膜域,但MARCH5有4個跨膜域,MARCH6甚至有14個跨膜域,而MARCH7和MARCH10結構特征類似,均不含有跨膜域,其序列同源性僅存在于環指結構域[3,8,10]。MARCH1和MARCH8、MARCH2和MARCH3,以及MARCH4和MARCH9之間高度序列同源,表明這些兩兩高度同源蛋白具有部分相似功能[1]。
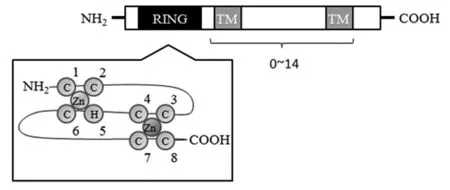
圖1 MARCH結構示意圖
2 MARCH家族各成員的表達與生物學功能
MARCH家族各成員各具特點和功能,其若干成員還能協同參與多種生理病理功能,見表1。
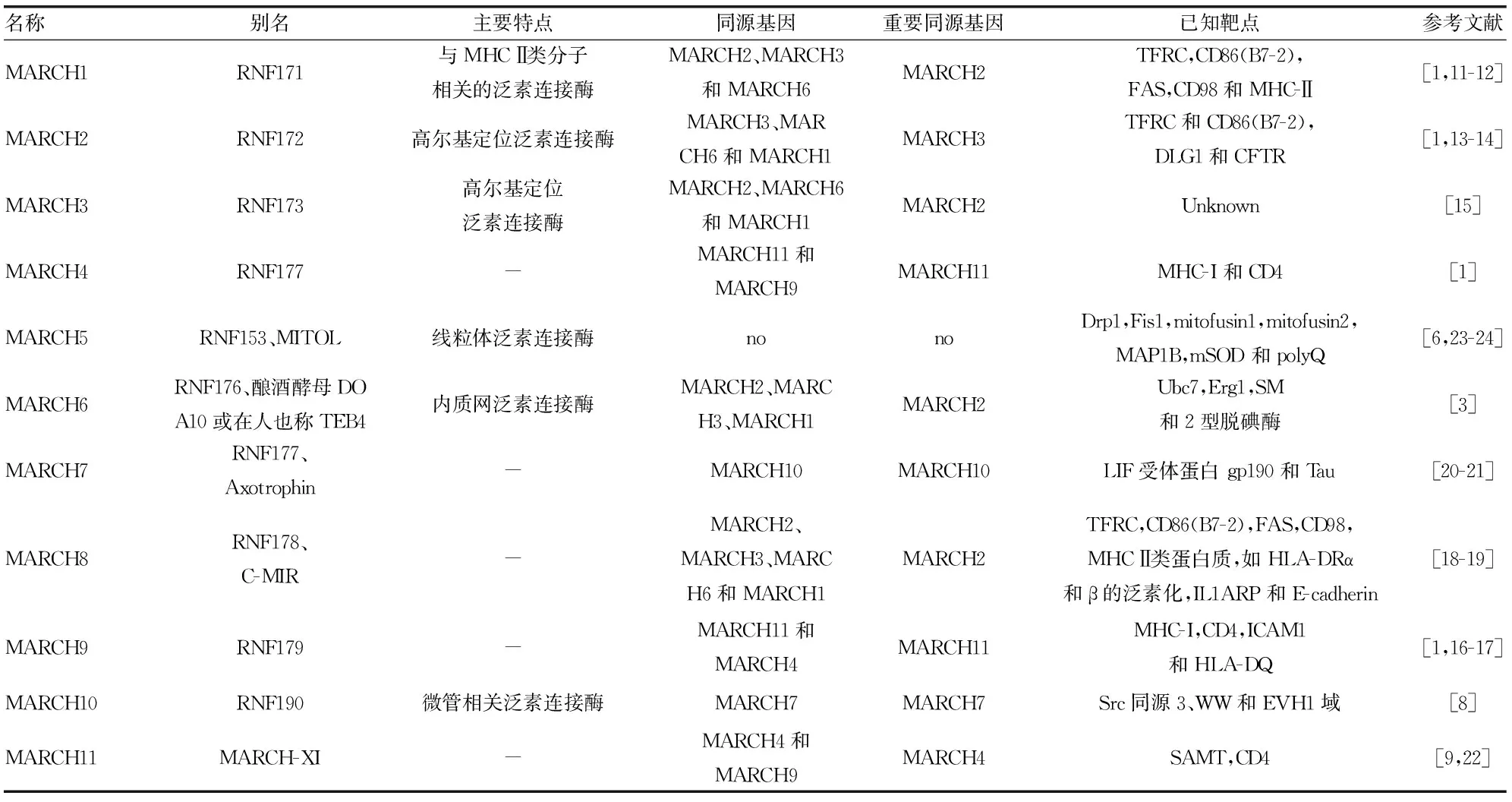
表1 MARCH各成員概要
2.1 MARCH1、MARCH2、MARCH3、MARCH4、MARCH9和MARCH8及其與免疫功能
2.1.1 MARCH1 MARCH1,又稱RNF171,含約289個氨基酸,相對分子質量約為3.2×104,是一種與主要組織相容性復合體(MHC)-Ⅱ類分子相關的E3泛素連接酶,主要表達在次級淋巴組織、淋巴結和脾臟的濾泡B細胞上,并高表達于抗原提呈細胞、B細胞和樹突細胞(dendritic cells,DCs),在免疫系統中具有重要的調節作用[11]。MARCH1能泛素化轉鐵蛋白受體TFRC、CD86(B7-2)、FAS和MHC-Ⅱ類蛋白(如HLA-DRα和β),并通過多泡體(multivesicular body,MVB)分選到溶酶體,促使其降解[12]。Ohmura-Hoshino等[11]研究表明,在多器官障礙綜合征(MODS)病程中,MARCH1可能通過泛素化MHC-Ⅱ類分子來調控其表達水平,進而調控DCs的成熟狀態及啟動免疫應答反應,從而起著關鍵性作用,MARCH1可作為一個治療靶點,以降低MODS臨床病死率。MODS病程中MARCH1通過泛素化MHC-Ⅱ類分子來調控其表達并進一步調控DCs抗原提呈作用的具體機制尚需進一步深入研究。Toll作用蛋白(Tollip)會導致MARCH1表達下降而恢復MHC-Ⅱ類分子水平,因此,MARCH1可能是一種新的Tollip靶點。
盡管MARCH1表達僅限于次級淋巴器官[1],但它能被不同刺激物所誘導或抑制。如白細胞介素10(IL-10)在人初級單核細胞和小鼠B細胞中可調節其表達。MARCH1可作為IL-10誘導的對MHC-Ⅱ抗原呈遞途徑的免疫抑制效應的調控者。通過反向IL-10誘導MARCH1介導的泛素化和降解,CD83能增加樹突狀細胞MHC-Ⅱ和CD86(B7-2)。LPS誘導的DCs成熟可抑制MARCH1表達,允許新合成的MHC-Ⅱ類分子在細胞表面表達[12]。
2.1.2 MARCH2和MARCH3 在MARCH家族中,MARCH2和MARCH3是相關度最高的蛋白,有63%的蛋白序列相同[5],是兩個高爾基定位的E3泛素連接酶。MARCH2,含約246個氨基酸,相對分子質量約為2.7×104,廣泛表達于各種組織,定位于內質網膜、溶酶體膜和內涵體膜。MARCH2可介導TFRC和CD86(B7.2)的泛素化,并促進其隨后的內吞作用和通過MVB分選到溶酶體,從而調節細胞表面β腎上腺素受體的表達[13]。MARCH2能與DLG1相互作用。MARCH2是一種參與胞內轉運的突觸6結合蛋白[4]。Cheng等[14]報道,MARCH2通過其聯合適配蛋白CAL和syntaxin 6(STX6)泛素化并降解CFTR。 MARCH3,含約253個氨基酸,相對分子質量約為2.9×104,定位于早期內涵體膜和多通道膜,可能參與了內涵體運輸通路[5]。Koebis等[15]的研究顯示,MARCH3在虹鱒魚低表達,可能參與免疫系統、囊泡運輸和蛋白泛素化。
2.1.3 MARCH4和MARCH9 MARCH4,含約410個氨基酸,相對分子質量約為4.6×104,定位于高爾基體膜和多通道膜,可能介導MHC-Ⅰ和CD4的泛素化,并促進其后續內吞作用和通過MVB分選到溶酶體,從而參與免疫系統的調節[1]。MARCH9,又稱RNF179,含約346個氨基酸,相對分子質量約為3.8×104,廣泛表達于人組織,定位于高爾基體膜、多通道膜和溶酶體膜,可能介導MHC-Ⅰ、CD4和關鍵細黏附分子ICAM-1的泛素化,并促進其隨后的內吞作用和通過MVB分選到溶酶體,從而參與免疫系統的調節[16]。過表達MARCH9導致細胞表面MHC-Ⅰ、CD4和ICAM-1下調。此外,MARCH9僅特異性與HLA-DQ相互作用[17]。
2.1.4 MARCH8 MARCH8,也稱C-MIR,含約291個氨基酸,相對分子質量約為3.3×104,廣泛表達于各種組織,高表達于肺和胰腺,存在于未成熟樹突狀細胞,定位于細胞質泡膜、溶酶體膜和早期內涵體膜,能介導CD86(B7-2)、TFRC、FAS和MHC-Ⅱ類蛋白質,如HLA-DP、-DQ和-DR的泛素化,并促進其隨后的內吞作用和通過MVB分選到溶酶體。對乳腺癌細胞的研究發現,MARCH8能調控腫瘤壞死因子相關的凋亡誘導配體(TRAIL)受體的穩態細胞表面表達,是一種腫瘤細胞對TRAIL受體的潛在治療靶點[18]。MARCH8泛素化降解IL-1受體輔助蛋白(IL1ARP)抑制IL-1β對NF-κB和MAPK的激活,即MARCH8為IL-1β誘導的NF-κB激活途徑的特異性抑制劑,為解析天然免疫信號轉導的調控機制提供了更多的研究證據,同時也為將來的藥物設計提供可能的靶標。Kim等[19]研究發現,MARCH8在斑馬魚卵、斑馬魚和非洲爪蟾的早期胚胎表達,表明其可能在胚胎發育中發揮作用。MARCH8過表達和敲除均導致畸形發育。E-cadherin能被MARCH8泛素化,過表達MARCH8導致E-cadherin降低,表明MARCH8能通過調節E-cadherin的定位來調節細胞黏附強度而對胚胎起作用。因此,適當的MARCH8水平對胚胎生存和細胞黏附維持至關重要。
2.2 MARCH7、MARCH10和MARCH11及其與雄性生殖的關系。
2.2.1 MARCH7 MARCH7,又稱Axotrophin,含約690個氨基酸,相對分子質量約為7.8×104,是一種帶有單一RING結構域的蛋白質,其重要相似基因是MARCH10。MARCH7的RING結構域與C末端靠近,其N末端為富含絲氨酸/脯氨酸的高度無序結構[10]。MARCH7在脊椎動物中高度保守,尤其在哺乳動物。人類和鼠約85%的氨基酸序列同源,且有一個相同的RING-CH結構域,基因表達譜也相似。最近有研究者對MARCH7在人細胞和組織的表達譜進行篩選,發現MARCH7可在不同類型的人細胞和組織中表達:在人上皮細胞高表達,海馬和小腦的神經元和神經元祖細胞、脾血竇內皮細胞、骨髓巨核細胞、小腸隱窩肝細胞和肺泡巨噬細胞中選擇性表達,也能在睪丸早期生精細胞、附睪腺上皮表達。Zhao等[7]研究證實,MARCH7在大鼠精子細胞發育的中后期高表達,定位于頂體和鞭毛,可能參與精子頭部和鞭毛的形成。Lys48多泛素化鏈存在于大鼠精子細胞的頂體/acroplaxome、頸部、尾部中段和胞漿,MARCH7介導Lys48多泛素化鏈進入蛋白酶體被降解。
此外,MARCH7也與神經退行性疾病和免疫耐受相關。MARCH7為正常大腦發育所必需,March7敲除小鼠能存活并具有生育能力,但顯示過早神經退行性變和胼胝體的發育缺陷。MARCH7的功能與Foxp3和白血病抑制因子(LIF)相關,在T細胞的增殖、免疫耐受和神經細胞的發育中具有重要作用。Gao等[20]使用MARCH7缺失小鼠,發現其對T淋巴細胞應答具有極大影響,包括了干細胞的細胞因子LIF 8倍過度增殖和5倍過量釋放。他們進一步發現:MARCH7是LIF受體亞單位gp190靶向降解所必須的,這就意味著MARCH7在LIF信號調節中起直接作用。Nathan等[10]研究發現,MARCH7趨向于自身泛素化,并與兩個去泛素化酶相關-去泛素化酶ubiquitin-specific protease(USP)9X(定位在胞質)和USP7(定位在胞核)。Flach等[21]表明,MARCH7與Tau蛋白相互作用,當體外微管結合受損時,在微管結合區域能單泛素化Tau。
2.2.2 MARCH10 MARCH10,含約808個氨基酸,相對分子質量約為9.0×104,其重要相似基因是MARCH7。Iyengar等[8]研究發現MARCH10定位于大鼠長形精子細胞的主段,是一種精子形成期調控精子細胞鞭毛形成和穩定微管的泛素連接酶。March10基因產物的兩種同種型March10a和March10b主要在睪丸表達。MARCH10a與MARCH10b的功能不同:March10a編碼一種具有泛素連接酶活性的長環指蛋白,是一種微管相關蛋白,在微管結構中發揮作用;而March10b定位于胞質,編碼一種缺少環指結構域的短富含脯氨酸的蛋白質,即MARCH10b缺少泛素連接酶活性。MARCH10b在C末端區富含脯氨酸序列,表明其介導蛋白質與蛋白質相互作用,例如,與Src同源3、WW和EVH1域發生相互作用,因此,MARCH10b可充當適配器或支架蛋白的作用。
2.2.3 MARCH11 MARCH11含約402個氨基酸,相對分子質量約為4.4×104,高表達于睪丸,可能在精子形成期和精子鞭毛形成中發揮重要作用[9]。MARCH11主要表達于發育中的精子細胞,在腦和垂體也有少量表達,定位于胞質泡膜、早期內涵體膜。在早期圓形精子細胞中,MARCH11定位于反面高爾基體管網狀結構(TGN)和MVB,能泛素化精子形成期相關多拷貝跨膜(SAMT)蛋白,使其傳遞到晚期胞內體/溶酶體區室。SAMT家族蛋白是MARCH11的底物,MARCH11在小鼠精子細胞通過泛素連接酶活性將其經TGN-MVB通路遞送至溶酶體,可能參與哺乳動物精子發生[9,22]。此外,CD4是MARCH11的另一底物[9]。
2.3 MARCH5及其與線粒體相關 MARCH5,是一種線粒體泛素連接酶,又稱MITOL,含約273個氨基酸,相對分子質量約為3.1×104。MITOL擁有一個負責泛素連接酶活性的植物同源域(PHD)基序和4個跨膜域,可在腦、心臟、肝、肺、脾、胃、睪丸、骨和肌肉表達,定位于線粒體外膜和內質網膜,能分別通過降解線粒體分裂蛋白hFis1和Drp1、線粒體融合蛋白mitofusin1(Mfn1)和mitofusin2(Mfn2),以及微管相關蛋白1B(MAP1B)而對包括線粒體形態、運輸、與內質網的相互作用等線粒體動力學起關鍵性作用[6]。MARCH5作為Mfn1上游調控者,通過調控乙酰化Mfn1,阻止在應激狀態下過量Mfn1集聚,對維持線粒體穩態和細胞存活至關重要。MARCH5沉默細胞會表現出線粒體形態畸形。MARCH5缺失可導致線粒體伸長,通過阻斷Drp1活性和(或)促進Mfn1在線粒體的積聚而誘導細胞衰老,因此,MARCH5作為線粒體質量控制的調節器,可能在預防細胞衰老中發揮作用[23]。Yonashiro等[24]證實MITOL能阻斷S-亞硝基MAP1B-輕鏈1(LC1)介導的線粒體功能障礙與神經元細胞死亡。MARCH5能與線粒體的Mfn2相互作用,但不能與內質網的Mfn2相互作用。Mfn2的GTP酶域的K192是MARCH5主要的泛素化位點,MITOL通過使該位點泛素化來激活Mfn2,從而調控內質網-線粒體的相互作用。由此推測MARCH5可能通過調控Mfn2而參與神經退行性疾病的發病機制。MITOL泛素化SOD1突變體并減少SOD1突變所致活性氧產生,從而對線粒體起保護作用。MITOL能直接泛素化,并通過泛素-蛋白酶體通路促進線粒體多聚谷氨酰胺polyQ的降解,從而控制polyQ聚集體形成和細胞毒性,由此推測MITOL可以作為治療polyQ疾病的潛在靶點。
2.4 MARCH6及其與降脂相關 內質網E3泛素連接酶MARCH6,又稱釀酒酵母Doa10或在人也稱TEB4,含約910個氨基酸,相對分子摶量約為1.03×104,定位于內質網膜和多通道膜,參與內質網相關蛋白的降解。Doa10能通過RING結構域與泛素結合酶Ubc7相互作用,調控E2的降解。Doa10/Teb4能通過調控角鯊烯單加氧酶(SM)的降解,以達到甾醇體內平衡。研究發現Erg1(SM的同源物)是酵母中Doa10的底物,Doa10依賴Erg1的降解受羊毛甾醇所調控,高水平的lanosterol促使Erg1的降解。但在哺乳動物細胞中,是膽固醇刺激TEB4介導的SM降解,影響3-羥基-3-甲基戊二輔酶A還原酶和膽固醇合成途徑,從而為TEB4靶向治療高脂血癥提供了可能[3]。
3 展 望
MARCH家族作為E3泛素連接酶新近發現的一類,在調節許多生物過程中起著重要作用,其異常表達會導致某些疾病(如免疫功能異常、雄性不育、線粒體功能異常、高脂血癥等)的發生。MARCH家族決定了泛素-蛋白酶體通路底物的特異性,因此,高通量識別底物將是今后研究的主要方向。隨著MARCH家族蛋白相關領域研究的不斷深入,其關鍵性的新功能不斷被發現,其重要性將日益受到更為廣泛的關注。總之,深入研究MARCH家族,無論對于相關疾病發病機制的研究,還是對于這些疾病臨床治療和藥物的開發皆具有重要意義。
[1]Bartee E,Mansouri M,Hovey Nerenberg BT,et al.Downregulation of major histocompatibility complex class I by human ubiquitin ligases related to viral immune evasion proteins[J].J Virol,2004,78(3):1109-1120.
[2]Nakamura N.The role of the transmembrane RING finger proteins in cellular and organelle function[J].Membranes (Basel),2011,1(4):354-393.
[3]Zelcer N,Sharpe LJ,Loregger A,et al.The E3ubiquitin ligase March6degrades squalene monooxygenase and affects 3-Hydroxy-3-Methyl-Glutaryl coenzyme a reductase and the cholesterol synthesis pathway[J].Mol Cell Biol,2014,34(7):1262-1270.
[4]Nakamura N,Fukuda H,Kato A,et al.MARCH-Ⅱ is a syntaxin-6-binding protein involved in endosomal trafficking[J].Mol Biol Cell,2005,16(4):1696-1710.
[5]Fukuda H,Nakamura N,Hirose S.MARCH-Ⅲ is a novel component of endosomes with properties similar to those of MARCH-Ⅱ[J].J Biochem,2006,139(1):137-145.
[6]Nakamura N,Kimura Y,Tokuda M,et al.MARCH-V is a novel mitofusin 2-and Drp1-binding protein able to change mitochondrial morphology[J].EMBO Rep,2006,7(10):1019-1022.
[7]Zhao B,Ito K,Iyengar PV,et al.March7E3ubiquitin ligase is highly expressed in developing spermatids of rats and its possible involvement in head and tail formation[J].Histochem Cell Biol,2013,139(3):447-460.
[8]Iyengar PV,Hirota T,Hirose S,et al.Membrane-associated RING-CH 10(March10Protein) Is a Microtubule-associated E3Ubiquitin Ligase of the Spermatid Flagella[J].J Biol Chem,2011,286(45):39082-39090.
[9]Morokuma Y,Nakamura N,Kato A,et al.MARCH-XI,a novel transmembrane ubiquitin ligase implicated in ubiquitin-dependent protein sorting in developing spermatids[J].J Biol Chem,2007,282(34):24806-24815.
[10] Nathan JA,Sengupta S,Wood SA,et al.The ubiquitin E3ligase March7is differentially regulated by the deubiquitylating enzymes USP7and USP9X[J].Traffic,2008,9(7):1130-1145.
[11]Ohmura-Hoshino M,Matsuki Y,Mito-Yoshida M,et al.Cutting edge:requirement of MARCH-I-Mediated MHC Ⅱ ubiquitination for the maintenance of conventional dendritic cells[J].J Immunol,2009,183(11):6893-6897.
[12]Tze LE,Horikawa K,Domaschenz H,et al.CD83increases MHC Ⅱ and CD86on dendritic cells by opposing IL-10-driven March1-mediated ubiquitination and degradation[J].J Exp Med,2011,208(1):149-165.
[13]Han SO,Xiao K,Kim J,et al.March2promotes endocytosis and lysosomal sorting of carvedilol-bound β(2)-adrenergic receptors[J].J Cell Biol,2012,199(5):817-830.
[14]Cheng J,Guggino W.Ubiquitination and degradation of CFTR by the E3ubiquitin ligase March2through its association with adaptor proteins CAL and STX6[J].PLoS One,2013,8(6):e68001.
[15]Koebis JM,Rebl A,Kuehn C,et al.Comparison of splenic transcriptome activity of two rainbow trout strains differing in robustness under regional aquaculture conditions[J].Mol Biol Rep,2013,40(2):1955-1966.
[16]Hoer S,Smith L,Lehner PJ.MARCH-IX mediates ubiquitination and downregulation of ICAM-1[J].FEBS Lett,2007,581(1):45-51.
[17]Hor S,Ziv T,Admon A,et al.Stable isotope labeling by amino acids in cell culture and differential plasma membrane proteome quantitation identify new substrates for the March9transmembrane E3ligase[J].Mol Cell Proteomics,2009,8(8):1959-1971.
[18]Van De Kooij B,Verbrugge I,De Vries E,et al.Ubiquitination by the membrane-associated RING-CH-8(MARCH-8) ligase controls steady-state cell surface expression of tumor necrosis factor-related apoptosis inducing ligand (TRAIL) receptor 1[J].J Biol Chem,2013,288(9):6617-6628.
[19]Kim MH,Rebbert ML,Ro H,et al.Cell adhesion in zebrafish embryos is modulated by March8[J].PLoS One,2014,9(4):e94873.
[20]Gao W,Thompson L,Zhou Q,et al.Treg versus Th17lymphocyte lineages are cross-regulated by LIF versus IL-6[J].Cell Cycle,2009,8(9):1444-1450.
[21]Flach K,Ramminger E,Hilbrich I,et al.Axotrophin/March7acts as an E3ubiquitin ligase and ubiquitinates tau protein in vitro impairing microtubule binding[J].Biochim Biophys Acta,2014,1842(9):1527-1538.
[22]Yogo K,Tojima H,Ohno JY,et al.Identification of SAMT family proteins as substrates of March11in mouse spermatids[J].Histochem Cell Biol,2012,137(1):53-65.
[23]Nagashima S,Tokuyama T,Yonashiro R,et al.Roles of mitochondrial ubiquitin ligase MITOL/March5in mitochondrial dynamics and diseases[J].J Biochem,2014,155(5):273-279.
[24]Yonashiro R,Kimijima Y,Shimura T,et al.Mitochondrial ubiquitin ligase MITOL blocks S-nitrosylated MAP1B-light chain 1-mediated mitochondrial dysfunction and neuronal cell death[J].Proc Natl Acad Sci U S A,2012,109(7):2382-2387.
10.3969/j.issn.1671-8348.2016.29.039
國家自然科學基金資助項目(81300535);重慶市科委自然科學基金資助項目(cstc2013jcyjA10075)。 作者簡介:王美姣(1983-),醫師,碩士,主要從事生殖醫學與生殖生物學研究。△
R715.9
A
1671-8348(2016)29-4144-04
2016-02-18
2016-04-06)
