Dmrt基因在水生生物中的研究進展
2016-11-12 06:41:17李法君付春鵬羅永巨
水生生物學報 2016年5期
李法君 付春鵬 羅永巨
(1. 濰坊科技學院,壽光 262700; 2. 廣西水產科學研究院,南寧 530021)
Dmrt基因在水生生物中的研究進展
李法君1付春鵬1羅永巨2
(1. 濰坊科技學院,壽光 262700; 2. 廣西水產科學研究院,南寧 530021)
Dmrt(Doble-sex and Mab-3 Relatated Transcription factor)是指同果蠅Dsx基因和線蟲Mab-3基因同源的基因家族,其家族成員的主要特征是所編碼的多肽鏈中都包含一個具有 DNA 結合能力的保守基序-DM結構域。Dmrt基因是從無脊椎動物到脊椎動物都存在的古老基因,功能涉及性別決定與分化、許多組織和器官的形成及相關功能的維持等多方面。文章綜述了近來Dmrt基因在水生生物中的研究進展; 按從低等到高等的順序,梳理了水產動物Dmrt基因的功能; 分析了Dmrt1作為性別決定基因的歷程; 進而對Dmrt基因在水生生物的研究熱點做了展望。
Dmrt基因;Dsx基因;性腺;甲殼動物;魚類;水生生物
Dmrt(Doble-sex and Mab-3 Relatated Transcription factor)是指與果蠅(Drosophila melanogaste)的性別決定基因Doublesex(Dsx)和秀麗隱桿線蟲(Caenorhabditis elegans)的性別決定基因Maleabnormal-3(Mab-3)同源的基因[1—3],最早發現于果蠅中[4]。不同的Dmrt基因構成一個基因家族,該家族成員的主要特征是所編碼的多肽鏈都包含一個能與DNA結合的保守基序-DM(Doublesex and Mab-3)結構域,該結構域在不同進化類型的生物中具有相當的保守性[5]。Dmrt基因是一類轉錄調控因子,該因子以鋅指結構與目的DNA序列結合,通過調節目的基因的轉錄參與發育調節過程。研究表明,在整個脊椎動物和無脊椎動物中,只有性別決定基因Dmrt才具有進化保守性,也是參與性別決定最古老的發育基因家族[6]。 Dmrt基因的功能主要體現在性別決定、性腺分化、許多組織和器官的形成及相關功能的維持等多方面[3,7,8]。近來在水生生物,特別是甲殼動物中,有關Dmrt基因的研究已經取得了眾多成果。而截止目前為止,還缺乏對其進行卻進行全面總結的報道。鑒于此,本文對Dmrt基因在水生生物中的研究進行了綜述,并探討了Dmrt1基因及與性別決定的關系,以期為以后更好地研究水生生物Dmrt基因提供參考。
1 Dmrt基因家族的結構特征及種類
Dmrt基因家族成員的主要特征是其編碼的多肽鏈中含有與DNA結合的保守區域-DM結構域。DM結構域中的鋅指結構含有6個半胱氨酸(C)和2個組氨酸(H),這8個保守的氨基酸形成與Zn離子結合的兩個位點(CCHC和HCCC)。DM結構域的羧基端有一個可識別DNA的α-螺旋,用以穩定DNADM結構域結合體[3]。從無脊椎動物到脊椎動物,Dmrt家族成員編碼的DM結構域多肽序列高度保守,而在DM結構域外的序列相似性卻很低。
不同物種的Dmrt基因家族成員不盡相同,在哺乳類、鳥類、爬行類、兩棲類、魚類中,Dmrt基因家族的種類一般為8種(Dmrt1-8)[9,10]、4種(Dmrt1-4)[11]、6種(Dmrt1-6)[12]、5種(Dmrt1-5)[13]、5種(Dmrt1-5)[14,15]。在無脊椎動物中,Dmrt基因家族成員數目因物種而異。
2 水生生物Dmrt基因的功能
2.1腔腸動物和扁形動物
目前為止,動物界中僅低等的海綿動物沒有發現Dmrt基因[16,17],從腔腸動物開始檢測到Dmrt基因的存在(表1)。鹿角珊瑚(Acropora millepora)為雌雄同體的生物,一般在春季產卵之前分化產生雌雄生殖細胞,在此過程中鹿角珊瑚的AmDM1基因表達水平會明顯升高,暗示AmDM1基因可能參與了雌雄生殖細胞的分化過程[18]; 最新的研究表明,在海葵(Nematostella vectensis)中發現11個Dmrt基因(Nv-DMRT A-Nv-DMRT K),7個基因在雌雄個體間存在表達差異,其中Nv-DMRT B,F,G,H,I,K在雄性中的表達量明顯高于雌性,而Nv-DMRT E在雌性中的表達量要高于雄性。進一步的分析表明,Nv-DMRT基因在海葵雌雄配子的發生過程中發揮作用[19]。雖然扁形動物的真渦蟲(Schmidtea mediterranea)為雌雄同體動物,但Smed-dmd-1基因在雄性生殖系統中特異性表達,而且Smed-dmd-1基因對于精巢和雄性附屬生殖腺的發生、發展、維持及重新生成是必須的[20],被認定為性別決定基因[14]。同屬扁形動物的曼氏血吸蟲(Schistosoma mansoni)(雌雄異體)中,dmd-1基因在雄性個體的表達量要顯著高于雌性,表現為明確的性別特異性表達[20]。
綜上所述,腔腸動物Dmrt基因的功能主要體現在參與雌雄配子的分化過程,而在扁形動物中,Dmrt基因的功能則進一步升級為調控雄性性腺的發育(表1),參與性別決定。
2.2軟體動物
于非非等[21]克隆了馬氏珠母貝(Pinctada matensii)的3個Dmrt基因(Dmrt2,3,4),并預測Dmrt2基因可能在馬氏珠母貝的性別分化過程中起重要作用; 進而,Yu等[22]進一步研究了Dmrt2基因的組織和時空表達,結果顯示,Dmrt2基因主要在馬氏珠母貝雄性生殖腺中表達,其轉錄水平在雄性生殖腺的起始階段較低,在生殖腺的成熟階段最高,此外在鰓中也檢測到微量表達。研究表明,Dmrt2參與了馬氏珠母貝精子的形成過程。馮政夫等[23]克隆了櫛孔扇貝(Chlamys farreri)的Cf-dmrt4-like基因,該基因從受精卵到匍匐幼蟲各發育時期均有表達,其中卵裂期表達量較高; 在不同發育時期的精巢中均有表達,以成熟期的精巢表達量最高; 在卵巢中未見表達。由此推測,櫛孔扇貝Cf-dmrt4-like基因參與了個體的早期發育,并在兩性成體中發揮著不同的作用。周麗青等[24]在蝦夷扇貝(Patinopecten yessoensis)中克隆了PyDmrt3和PyDmrt4兩個基因,并推測PyDmrt4 可能參與了調控雄性性別的形成過程。張娜等[25]在長牡蠣(Crassostrea gigas)中發現了CgDsx,CgDmrtA2兩個基因,CgDsx 基因可能調控長牡蠣早期的胚胎發育,而CgDmrtA2基因可能參與了胚胎中后期的發育過程,與神經的形成相關。太平洋牡蠣(Crassostrea gigas)的Cg-DMl基因在雌雄個體中均有表達,但在雄性生殖腺的表達量明顯高于雌性,表明Cg-DMl參與了太平洋牡蠣雄性性腺的發育過程[26]。黑蝶真珠蛤(Pinctada margaritifera)是雌雄同體雄性先成熟的物種,其中的性轉機制尚不清楚,研究人員通過性腺轉錄組數據分析了其中可能的性別分化和決定基因。研究表明,pmarg-dmrt基因和其他兩個基因可能參與了精卵巢的轉化過程[27]。在耳鮑(Haliotis asinin)中,HADMRT1基因在雄性的精巢中特異性表達,表明HADMRT1在耳鮑的精巢發育過程中發揮作用[28]。

表1 Dmrt基因在水生無脊椎動物的研究匯總表Tab. 1 Summary of the study for Dmrt genes in the aquatic invertebrate
據此,軟體動物的Dmrt基因主要在雄性的生殖腺及精子形成過程中發揮作用(表1)。
2.3甲殼動物
水蚤科動物在環境適宜的條件下多為雌性個體,行孤雌生殖,可以使種群大量繁殖; 在環境惡劣(光照周期縮短、食物短缺、群體密度增大、水質惡劣)的情況下,雌性個體所帶的卵會產生雄性和雌性個體行有性生殖,產生休眠卵渡過惡劣環境,保證物種的持續性。由此可見,環境變化是誘導水蚤科動物生殖策略改變的原因,而此過程中所蘊含的性別決定和生殖策略改變的遺傳機制也成為最近研究的熱點。
Zhang等[29,30]在隆線溞(Daphnia carinata)中相繼研究了DM結構域的兩個基因DapcaDsx1和DapcaDsx2,并且在雌性幼體、孤雌生殖的雌性、有性生殖雌性的早期階段、有性生殖的雌性、休眠卵五個不同生殖階段研究了DapcaDsx1和DapcaDsx2表達情況。結果這兩個基因均顯示出相似的變化規律,在有性生殖雌性的早期階段表達量最高,且有性生殖階段的表達量明顯高于孤雌生殖階段。DapcaDsx1和DapcaDsx2呈現明顯的性別特異性表達,暗示它們可能在隆線溞由孤雌生殖向有性生殖階段過渡的過程中發揮作用。蚤狀溞(Daphnia pulex)中的研究表明,Dpdsx1基因在有性生殖雄性個體的第一觸角、第一胸肢、復眼中的表達量最高,在行孤雌生殖和有性生殖的雌性個體中,上述三個組織中的表達量顯著下降。且Dpdsx1基因在蚤狀溞有性生殖的雄性階段表達量最高,在雌性幼體階段表達量最低。與隆線溞相似,蚤狀溞Dpdsx1基因也主要在生殖模式的轉換、性別分化過程中發揮作用[31]。Kato等[32]首先在大型溞(Daphnia magna)中克隆到三個Dmrt基因:DMRT11E,DMRT93B和DMRT99B,其中DMRT11E和DMRT99B在卵巢中的表達量要高于精巢,而DMRT93B則在雄性的精巢中特異性的表達。由于這三個基因在胚胎階段沒有表達,且不具有性別二態性表達,所以它們不是性別決定基因[33]。進而,Kato等[34]在大型溞中克隆得到兩種Dsx 基因DapmaDsx1和DapmaDsx2,其中DapmaDsx1編碼兩種不同的亞型DapmaDsx1-α和DapmaDsx1-β。DapmaDsx1展現出明顯的性別二態性表達,在雄性胚胎的形成過程中,DapmaDsx1表達量明顯升高,而在雌性胚胎中則沒有這種現象。進一步在雄性胚胎中沉默DapmaDsx1,可誘導其卵巢成熟,產生雌性特征; 相反,在雌性胚胎中異位表達Dapma Dsx1,可使其產生雄性特性。表明DapmaDsx1雄性性別決定中起著關鍵作用。因此,DapmaDsx1被認為是大型溞的性別決定基因[14]。Toyota等[33]進一步在蚤狀溞、盔形溞(Daphnia galeata)、網紋溞(Ceriodaphnia dubia)中分別克隆了兩個Dsx基因,在多刺裸腹水蚤(Moina macrocopa)中克隆了一個Dsx基因,這些基因均表現為明顯的性別二態性表達,表明它們具有相似的功能。
近來,Dmrt基因在十足目(蝦蟹類)中的研究也取得了相應進展。中華絨螯蟹(Eriocheir sinensis)中發現一個EsDmrt-like基因,EsDmrt-like在精巢中特異性表達,進一步的分析表明,EsDmrt-like主要在精巢中的支持細胞中表達,在成熟的精子中則沒有檢測到其表達,而支持細胞是為發育中的精子提供保護和營養的細胞,精子發育的各個階段都是發生在支持細胞的表面。此結果表明在中華絨螯蟹中,EsDmrt-like是雄性精巢發育的關鍵因子[35]。羅氏沼蝦(Macrobrachium rosenbergii)中發現兩個Dmrt基因:MroDmrt11E和MroDmrt99B基因,這兩個基因均在精巢中高度表達。通過RNA干擾分別沉默MroDmrt11E和MroDmrt99B基因,結果表明,MroDmrt11E正調控胰島素樣促雄腺激素(Insulinlike androgenic gland hormone,IAG)基因的表達[36]。
綜上所述,甲殼動物Dmrt基因主要參與雄性精巢的發育過程,且在大型溞中Dsx1基因被認定為性別決定基因(表1)。
2.4魚類
性別決定魚類種類繁多,其性別決定方式和機制也呈現多樣化。而Dmrt1基因作為魚類的性別決定基因的證據來源于青鳉(Oryzias latipe)(XX/XY型,雌性同配、雄性異配)[37—40]。DMY(DM domain gene on the Y chromosome)基因位于青鳉Y染色體上的性別決定區域,是進化上相對年輕的基因,由位于常染色體的Dmrt1基因復制、轉移到Y 染色體上而形成[41—43]。在雌性青鳉中過度表達DMY基因,可使雌性青鳉產生性逆轉[37]; 相反,在雄性青鳉中沉默DMY基因或者使DMY基因產生突變,可使雄性個體逆轉為雌性[44,45]。此外,青鳉的DMY基因只存在于Y染色體上,在X染色體上沒有檢測到它的同源基因,表達也只限于雄性的胚胎和成體精巢中的支持細胞,表明DMY基因對青鳉精巢的分化和功能維持至關重要[46,47]。然而DMY基因也僅見于青鳉,而其他魚類卻沒有發現此基因[48],表明青鳉DMY基因的性別決定功能在魚類中不具有普遍性[49]。Chen等[50]首次構建了比目魚-半滑舌鰨(Cynoglossus semilaevis)(ZZ/ZW型,雄性同配、雌性異配)全基因組精細圖譜,發現半滑舌鰨dmrt1基因是Z染色體連鎖、雄性特異表達、精巢發育必不可少的關鍵基因,表現出性別決定基因的特性。
性腺發育自然界中魚類存在雌雄異體和雌雄同體兩大類,而后者又可分為精巢先成熟和卵巢先成熟兩種類型。而Dmrt1基因在上述三種魚類的性腺發育過程中均發揮重要作用。
雌雄異體的魚類:在革胡子鲇(Clarias gariepinus)[51]、稀有鮈鯽(Gobiocypris rarus)[52]、尼羅羅非魚(Oreochromis niloticus)[53]、牙鲆(Paralichthys olivaceus)[54,55]、紅尾劍魚(Xiphophorus maculatus)[56]、紅鰭東方鲀(Takifugu rubripes)[57]、奧利亞羅非魚(O. aurea)[58]、異育銀鯽(Carassius auratus gibelio)[59]等魚類中,Dmrt1基因在精巢中檢測到特異性表達(表2)。在湖鱘(Acipenser fulvescens)[60]、斑馬魚(Danio rerio)[61]、大西洋鱈(Gadus morhua)[62]、銀漢魚(Odontesthes bonari-ensis)[63]、虹鱒(Oncorhynchus mykiss)[64]、密西西比鏟鱘(Scaphirhynchus platorynchus)[65]、南方大口鯰(Silurus meridionals)[66]、團頭魴(Megalobrama amblycephala)[67]、許氏平鲉(Sebastods schlege-lii)[68]、腋孔蟾魚(Halobatrachus didactylus)[69]中,Dmrt1基因主要在精巢中表達,此外在卵巢中也檢測到微量的表達(表2)。
雌雄同體的魚類:在此類型的魚類中,精巢和卵巢之間存在相關轉化的階段。而誘導精卵巢轉化的遺傳信號分子,很可能出現在胚胎或幼體之后,而Dmrt1基因同樣在其中發揮作用。雌性先成熟的黑鯛(Acanthopagrus schlegeli)[70]、金頭鯛(Sparus auratus)[71]在卵巢向精巢轉化的過程中,Dmrt1基因表達量明顯升高; 而在雄性先成熟的點帶石斑魚(Epinephlus coioides)[72]、細棘海豬魚(Halichoeres tenuispinis)[73]、黃鱔(Monopterus albus)[74]中,在由精巢向卵巢轉的過程中,Dmrt1基因表達量則呈現相反的表達趨勢。
雖然Dmrt1基因對魚類精巢功能的維持起著關鍵作用,但Dmrt家族的其他成員對已經分化的性腺的正常發育和功能維持也有重要作用。在青鳉中,Dmrt3基因在成體的精巢中就有所表達,而Dmrt2和Dmrt4基因在成體精巢和卵巢中均有所表達[75]。最新的研究表明,黃鱔的Dmrt2,3,4,5 基因在發育生殖細胞中均有表達[76]。在尼羅羅非魚[77]、奧利亞羅非魚[78]、斑馬魚[61]中也存在類似的現象。
其他功能Dmrt家族基因除了參與性別決定和性腺分化與維持之外,還在魚類神經系統和感覺器官的發育[79,80]、體節形成(沉默dmrt2b可導致尾部彎曲和U型體節)[81—83]、以及其他器官形成與功能維持方面[67,75,82]發揮作用。
據此,Dmrt基因家族在魚類中作用主要體現在性腺分化(表2)、性別決定、體節發育、各種器官的形成和維持等方面。
3 Dmrt1基因功能的思考
在低等(腔腸動物和扁形動物)和較低等(軟體動物)的無脊椎動物中,現有的研究表明,Dmrt1基因“主管”配子分化與性腺發育。眾所周知,性別決定和性腺分化是緊密聯系的,“主管”性腺分化的基因隨著生物的進化,很可能升級為性別決定基因。在渦蟲中,Dmrt1基因作為性別決定基因已初見端倪[20]; 在節肢動物門中其性別決定地位得以鞏固,達到高峰。例如在果蠅中,sxl(sex-lethal)-tra(transformer)-dsx的性別決定通路已經得以論證[39,40,85];甲殼綱的水蚤科中Dsx基因也是其性別決定基因[34];十足目(蝦蟹類)中存在特殊腺體-促雄腺,其轉錄的IAG是十足目的性別決定基因[86—90],而Dmrt則是IAG基因的上游調控基因,正調控IAG的表達[36]。此時,Dmrt基因仍在十足目的性別決定通路中發揮作用,但其性別決定地位的作用已經開始弱化。根據Kopp的理論:在生物的進化過程中,性別決定機制也保持較快的進化速率,以至于性別決定的“主導”基因很少能在這個機制的“頂端”保存較長的時間[40]。例如,sxl和tra這兩個基因僅是昆蟲性別決定基因,在昆蟲之外的物種中雖然存在,但其作用卻不是性別決定; 再如,SRY基因僅在哺乳動物中行使特異的性別決定功能,在非哺乳的脊椎動物中,SRY基因就不存在。因此,(魚類)青鳉的DMY基因作為性別決定基因也僅僅是個別現象,是由于Dmrt1基因復制產生的“副作用”,這可能與青鳉特殊的進化地位有關[15]; 半滑舌鰨中Dmrt1基因除了在Z染色體上,在W染色體上也存在一個假基因化拷貝(Pseudogenized copy),在性逆轉的雄魚(ZW型)中這個假基因化拷貝被激活,通過劑量補償效益達到與正常雄魚相同的表達水平[50]。但是以上現象在魚類不存在普遍性。并且研究表明,在所有發現Dmrt1基因的魚類中,Dmrt1基因無一例外地在魚類的性腺發育中發揮作用[39]。至此,Dmrt1作為性別決定基因的地位進一步削弱。雖然在兩棲類、爬行類、鳥類等非哺乳的脊椎動物中,Dmrt1和Sox9基因決定性別分化[6]。但Dmrt1基因作為性別決定基因也僅見于非洲爪蟾(Xenopus laevis)[91]和原雞(Gallus gallus)[92]兩個物種。而且研究表明,Dmrt1不可能是鳥類性別決定的開關基因[93]。在哺乳類脊椎生物中,SRY基因是其真正的性別決定基因[6]; 進而縱觀無脊椎動物到脊椎動物的各個物種,不難發現,調控性腺(或配子)的分化與發育是Dmrt1基因的主導功能。例如,人類的Dmrt1基因是第一個在脊椎動物中得以鑒定的DM結構域基因,是在人類睪丸中發現的,定位于人類常染色體(9p24.3)上。該區域的缺失會導致睪丸發育異常,然而到目前為止沒有發現該基因的點突變影響人類的性別決定[94]。
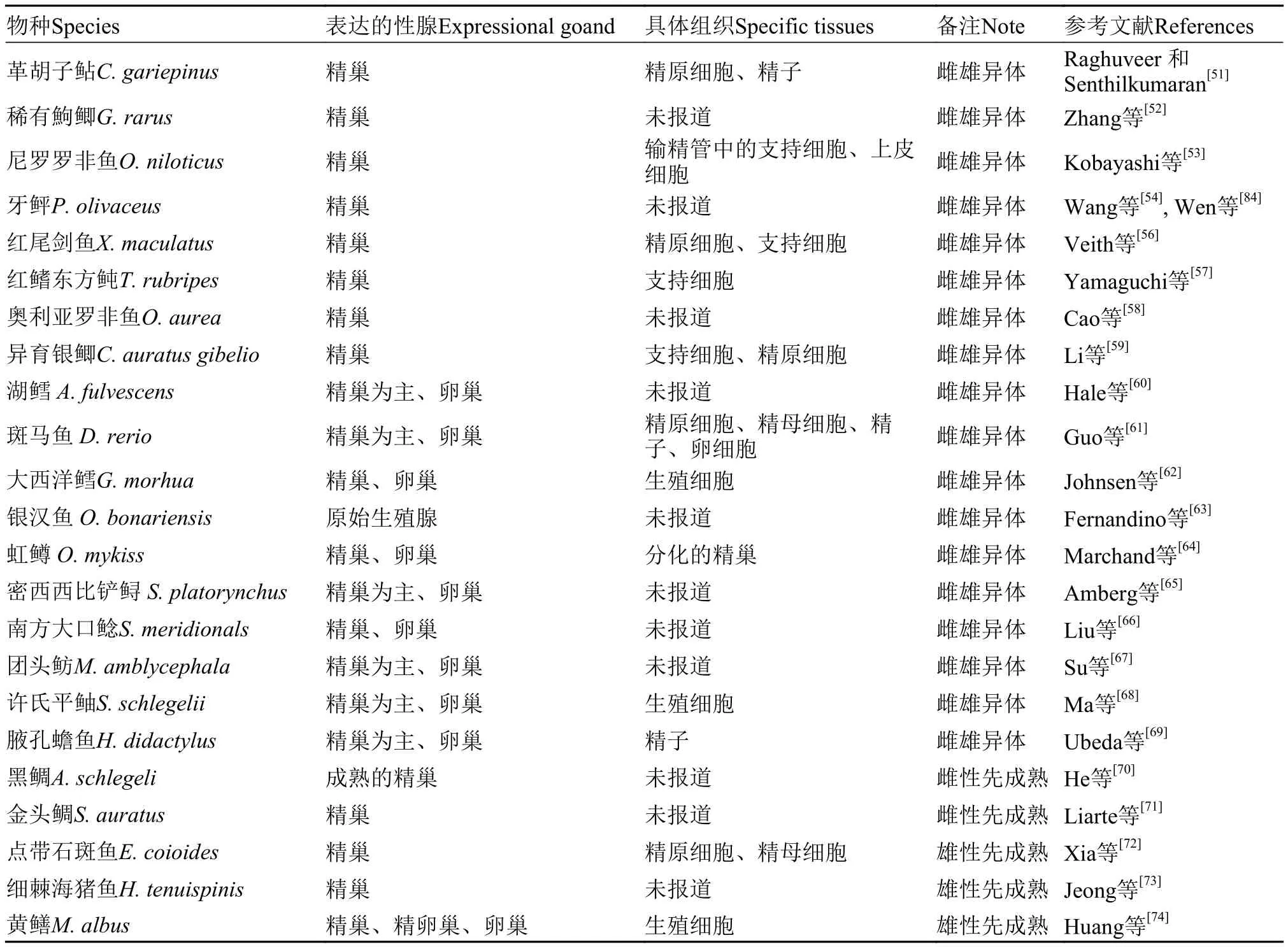
表2 Dmrt1基因在魚類性腺中的表達Tab. 2 Expression of Dmrt1 genes in fish gonad
作為一個在無脊椎動物和脊椎動物中均廣泛存在的古老基因,結構相對保守的Dmrt基因很可能在功能上也具有相當的保守性。如上所述,從低等的腔腸動物到高等的哺乳動物,Dmrt1基因的功能更多地是體現在性腺分化,特別是雄性性腺分化及精子成熟方面。而Dmrt1作為性別決定基因,現有的研究成果表明,最早出現于渦蟲,在節肢動物中其性別決定基因地位達到高峰,并且在甲殼綱中性別決定基因地位已經開始弱化。在眾多的脊椎動物中,Dmrt1作為性別決定基因地位已接近“尾聲”。僅在個別物種-青鳉、半滑舌鰨、非洲爪蟾、原雞中是性別決定基因。
4 展望
雖然有關Dmrt家族基因的研究在水生生物中取得了較大的進展,但尚有許多工作需要深入開展。從低等的腔腸動物到高等的脊椎動物-魚類,幾乎都存在雌雄同體的生物,在這些生物中,Dmrt基因無一例外地都是促進雄性配子或雄性生殖腺的發育,可見其功能的保守性,Dmrt多肽作為轉錄調節因子,眾多的研究也為揭示其蘊含的分子機制提供了堅實的基礎; 但每個物種具體的調控機制,特別是以Dmrt基因作為性別決定基因的物種,它們之間是否存在相似的調控網絡,還需進一步研究; 在蝦蟹類中,有關Dmrt家族基因的研究也僅見于河蟹和羅氏沼蝦,其他十足目物種的Dmrt序列及功能還未見報道; 有趣的是,半滑舌鰨的研究還發現,半滑舌鰨的性染色體并不和已知性染色體的魚類共祖先,而是和雞的ZW染色體共起源,揭示半滑舌鰨和雞性染色體的趨同進化現象[50]。而在半滑舌鰨和雞中,Dmrt1都是性別決定基因。這提醒我們,在性染色體與這兩個物種趨同進化的其他物種中,Dmrt1是否也是性別決定基因,值得深入探索。
[1]Zarkower D. Establishing sexual dimorphism:conservation amidst diversity [J]? Nature Reviews Genetics,2001,2(3):175—185
[2]Burtis K C,Baker B S. Drosophila doublesex gene controls somatic sexual differentiation by producing alternatively spliced mRNAs encoding related sex-specific polypeptides [J]. Cell,1989,56(6):997—1010
[3]Zhu L,Wilken J,Phillips N,U,et al. Sexual dimorphism in diverse metazoans is regulated by a novel class of intertwined zinc fingers [J]. Genes & Development,2000,14(14):1750—1764
[4]Hildreth P E. Doublesex,a recessive gene that tranforms both males and females of drosophila into intersexes [J]. Genetics,1965,51(4):659—678
[5]Erdman S E,Burtis K C. The Drosophila doublesex proteins share a novel zinc finger related DNA binding domain [J]. The EMBO Journal,1993,12(2):527—535
[6]Cheng Z H. The function and trait of Dmrt gene [J]. Journal of Anhui Agricultural Agricultural,2006,34(5):835—836 [程子華. Dmrt基因的功能和特點. 安徽農業科學,2006,34(5):835—836]
[7]Ellegren H. Hens,cocks and avian sex determination. A quest for genes on Z or W [J]? Embo Reports,2001,2(3):192—196
[8]Kim S,Kettlewell J R,Anderson R C,et al. Sexually dimorphic expression of multiple doublesex-related genes in the embryonic mouse gonad [J]. Gene Expression Patterns,2003,3(1):77—82
[9]Ottolenghi C,Fellous M,Barbieri M,et al. Novel paralogy relations among human chromosomes support a link between the phylogeny of doublesex-related genes and the evolution of sex determination [J]. Genomics,2002,79(3):333—343
[10]Hong C S,Park B Y,Saint J P. The function of Dmrt genes in vertebrate development:it is not just about sex[J]. Developmental Biology,2007,310(1):1—9
[11]Wang H,Wang T R,Yuan J,et al. Advances in Dmrt gene family of vertebrates [J]. Guizhou Agricultural Science,2012,40(5):148—152 [汪海,王婷茹,袁靜,等. 脊椎動物Dmrt基因的研究進展. 貴州農業科學,2012,40(5):148—152]
[12]Wang Z,Miyake T,Edwards S V,et al. Tuatara(Sphenodon) genomics:BAC library construction,sequence survey,and application to the DMRT gene family [J]. Journal of Heredity,2006,97(6):541—548
[13]Bewick A J,Anderson D W,Evans B J. Evolution of the closely related,sex-related genes dmw and dmrt1 in african clawed frogs(Xenopus) [J]. Evolution,2011,65(3):698—712
[14]Picard A L,Cosseau C,Mouahid G,et al. The roles of Dmrt(Double sex/Male-abnormal-3 Related Transcription factor) genes in sex determination and differentiation mechanisms:Ubiquity and diversity across the animal kingdom [J]. Comptes Rendus Biologies,2015,338(7):451—462
[15]Cao J L,Chen J J,Gan X,et al. Advances of Dmrt gene for fish [J]. Journal of Guangdong Ocean University,2011,31(1):94—98 [曹謹玲,陳劍杰,甘西,等. DMRT基因的研究進展. 廣東海洋大學學報,2011,31(1):94—98]
[16]Bellefroid E J,Leclère L,Saulnier A,et al. Expanding roles for the evolutionarily conserved Dmrt sex transcriptional regulators during embryogenesis [J]. Cellular & Molecular Life Sciences,2013,70(20):3829—3845
[17]Wexler J R,Plachetzki D C,Kopp A. Pan-metazoan phylogeny of the DMRT gene family:a framework forfunctional studies [J]. Development Genes and Evolution,2014,224(3):175—181
[18]Miller S W,Hayward D C,Bunch T A,et al. A DM domain protein from a coral,Acropora millepora,homologous to proteins important for sex determination [J]. Evolution & Development,2003,5(3):251—258
[19]Traylor N G,Kane E G,Sombatsaphay V,et al. Sex-specific and developmental expression of Dmrt genes in the starlet sea anemone,Nematostella vectensis [J]. Evolutionary Developmental Biology,2015,6(1):13—19
[20]Chong T,Collins J J,Brubacher J L,et al. A sex-specific transcription factor controls male identity in a simultaneous hermaphrodite [J]. Nature Communications,2013,4:1814—1826
[21]Yu F F,Zhou L,Wang M F,et al. Cloning and sequence analysis of three DM domain in Pinctada matensii [J]. Journal of Agricultural Biotechnology,2007,15(5):905—906 [于非非,周莉,王梅芳,等. 馬氏珠母貝(Pinctada matensii)3個DM結構域的克隆及序列分析.農業生物技術學報,2007,15(5):905—906]
[22]Yu F F,Wang M F,Zhou L,et al. Molecular cloning and expression characterization of dmrt2 in akoya pearl oysters,Pinctada martensii [J]. Journal of Shellfish Research,2011,30(2):247—254
[23]Feng Z F,Shao M Y,Sun D P,et al. Cloning,characterization and expression analysis of Cf-dmrt4-like gene in Chlamys farreri [J]. Journal of Fishery Scicnece of China,2010,17(5):930—940 [馮政夫,邵明瑜,孫大鵬,等. 櫛孔扇貝Cf-dmrt4-like基因的克隆、序列特征及表達分析. 中國水產科學,2010,17(5):930—940]
[24]Zhou L Q,Yang A G,Wang Y Q,et al. Sequence analysis of DM domain in two Dmrt genes of three sex types of Patinopecten yessoensis [J]. Marine Sciences,2015,39(3):19—25 [周麗青,楊愛國,王清印,等. 蝦夷扇貝不同性別類型2個Dmrt基因DM結構域分析. 海洋科學,2015,39(3):19—25]
[25]Zhang N,Huang W,Xu F,et al. Expression of two Dmrt family genes in the pacific oyster Crassostrea gigas [J]. Oceanologia et Limnologia Sinica,2015,46(3):717—724 [張娜,黃雯,許飛,等. 長牡蠣(Crassostrea gigas)兩個Dmrt基因家族的時空表達. 海洋與湖沼,2015,46(3):717—724]
[26]Naimi A,Martinez A S,Specq M L,et al. Identification and expression of a factor of the DM family in the oyster Crassostrea gigas [J]. Comparative Biochemistry and Physiology Part A Molecular & Integrative Physiology,2009,152(2):189—196
[27]Teaniniuraitemoana V,Huvet A,Levy P,et al. Gonad transcriptome analysis of pearl oyster Pinctada margaritifera:identification of potential sex differentiation and sex determining genes [J]. BMC Genomics,2014,15(1):1—20
[28]Klinbunga S,Amparyup P,Khamnamtong B,et al. Isolation and characterization of testis-specific DMRT1 in the tropical abalone(Haliotis asinina) [J]. Biochemical Genetics,2009,47(1—2):66—79
[29]Zhang M,Li H,Liu A,et al. Cloning,expression and cellular localization of the Doublesex gene in the water flea,Daphnia carinata,during different developmental stages[J]. Gene,2014,550(2):185—192
[30]Zhang M,Ma C,Lv W,et al. Molecular cloning,characterization and expression analysis of a Doublesex gene from Daphnia carinata(Crustacea:Cladocera) during different reproductive stages [J]. Genetics and Molecular Research,2015,14(2):5930—5942
[31]Xu S L,Zhou W,Chen P,et al. Identification and expression analysis of a doublesex1 gene in Daphnia pulex during different reproductive stages [J]. Development Genes and Evolution,2014,224(3):147—157
[32]Kato Y,Kobayashi K,Oda S,et al. Molecular cloning and sexually dimorphic expression of DM-domain genes in Daphnia magna [J]. Genomics,2008,91(1):94—101
[33]Toyota K,Kato Y,Sato M,et al. Molecular cloning of doublesex genes of four cladocera(water flea) species [J]. BMC Genomics,2013,14(1):239—252
[34]Kato Y,Kobayashi K,Watanabe H,et al. Environmental sex determination in the branchiopod crustacean Daphnia magna:deep conservation of a Doublesex gene in the sexdetermining pathway [J]. PLoS Genetics,2011,7(3):e1001345
[35]Zhang E F,Qiu G F. A novel Dmrt gene is specifically expressed in the testis of Chinese mitten crab,Eriocheir sinensis [J]. Development Genes and Evolution,2010,220(5—6):151—159
[36]Yu Y Q,Ma W M,Zeng Q G,et al. Molecular cloning and sexually dimorphic expression of two dmrt genes in the giant freshwater prawn,Macrobra-chium rosenbergii[J]. Agricultural Research,2014,3(2):181—191
[37]Matsuda M,Shinomiya A,Kinoshita M,et al. DMY gene induces male development in genetically female(XX)medaka fish [J]. Proceedings of the National Academy of Sciences,2007,104(10):3865—3870
[38]Matsuda M. Sex determination in the teleost medaka,Oryzias latipes [J]. Annual Review of Genetics,2005,39:293—307
[39]Matson C K,Zarkower D. Sex and the singular DM domain:insights into sexual regulation,evolution and plasticity [J]. Nature Reviews Genetics,2012,13(3):163—174
[40]Kopp A. Dmrt genes in the development and evolution of sexual dimorphism [J]. Trends in Genetics,2012,28(4):175—184
[41]Matsuda M,Nagahama Y,Shinomiya A,et al. DMY is a Y-specific DM-domain gene required for male development in the medaka fish [J]. Nature,2002,417(6888):559—563
[42]Kondo M,Hornung U,Nanda I,et al. Genomic organiza-tion of the sex-determining and adjacent regions of the sex chromosomes of medaka [J]. Genome Research,2006,16(7):815—826
[43]Indrajit N,Mariko K,Ute H,et al. A duplicated copy of DMRT1 in the sex-determining region of the Y chromosome of the medaka,Oryzias latipes [J]. Proceedings of the National Academy of Science,2002,99(18):11778—11783
[44]Masuyama H,Yamada M,Kamei Y,et al. Dmrt1 mutation causes a male-to-female sex reversal after the sex determination by Dmy in the medaka [J]. Chromosome Research,2012,20(1):163—176
[45]Paul B,Matsuda M,Lau E L,et al. Knock-down of DMY initiates female pathway in the genetic male medaka,Oryzias latipes [J]. Biochemical and Biophysical Research Communications,2006,351(4):815—819
[46]Kobayashi T,Matsuda M,Kajiura H,et al. Two DM domain genes,DMY and DMRT1,involved in testicular differentiation and development in the medaka,Oryzias latipes [J]. Developmental Dynamics,2004,231(3):518—526
[47]Matsuda M,Sato T,Toyazaki Y,et al. Oryzias curvinotus has DMY,a gene that is required for male development in the medaka,O. latipes [J]. Zoological Science,2003,20(2):159—161
[48]Kondo M,Nanda I,Hornung U,et al. Absence of the candidate male sex-determining gene dmrt1b(Y) of medaka from other fish species [J]. Current Biology,2003,13(5):416—420
[49]Zheng Y,Wang Z Z,Chen J Z. Progresses on the study of sex differentiation genes in fish [J]. Acta Hydrobiologica Sinica,2015,39(4):798—810 [鄭堯,王在照,陳家長. 調控魚類性腺分化基因的研究進展. 水生生物學報,2015,39(4):798—810]
[50]Chen S,Zhang G,Shao C,et al. Whole-genome sequence of a flatfish provides insights into ZW sex chromosome evolution and adaptation to a benthic lifestyle[J]. Nature Genetics,2014,46(3):253—260
[51]Raghuveer K,Senthilkumaran B. Identification of multiple dmrt1s in catfish:localization,dimorphic expression pattern,changes during testicular cycle and after methyltestosterone treatment [J]. Journal of Molecular Endocrinology,2009,42(5):437—448
[52]Zhang X,Zha J,Wang Z. Influences of 4-nonylphenol on doublesex and mab-3-related transcription factor 1 gene expression and vitellogenin mRNA induction of adult rare minnow(Gobiocypris rarus) [J]. Environmental Toxicology and Chemistry,2008,27(1):196—205
[53]Kobayashi T,Kajiura H,Guan G,et al. Sexual dimorphic expression of DMRT1 and Sox9 a during gonadal differentiation and hormone-induced sex reversal in the teleost fish Nile tilapia(Oreochromis niloticus) [J]. Developmental Dynamics,2008,237(1):297—306
[54]Wang D S,Zhou L Y,Kobayashi T,et al. Doublesex-and Mab-3-related transcription factor-1 repression of aromatase transcription,a possible mechanism favoring the male pathway in tilapia [J]. Endocrinology,2010,151(3):1331—1340
[55]Wen A,You F,Sun P,et al. Sexually dimorphic gene expression patterns during gonadal differentiation in olive flounder,Paralichthys olivaceus [J]. 2015,DOI:10.1163/15707563—00002470
[56]Veith A M,Sch?fer M,Klüver N,et al. Tissue-specific expression of dmrt genes in embryos and adults of the platyfish Xiphophorus maculatus [J]. Zebrafish,2006,3(3):325—337
[57]Yamaguchi A,Lee K H,Fujimoto H,et al. Expression of the DMRT gene and its roles in early gonadal development of the Japanese pufferfish Takifugu rubripes [J]. Comparative Biochemistry and Physiology Part D∶ Genomics & Proteomics,2006,1(1):59—68
[58]Cao J,Cao Z,Wu T. Generation of antibodies against DMRT1 and DMRT4 of Oreochromis aurea and analysis of their expression profile in Oreochromis aurea tissues[J]. Journal of Genetics and Genomics,2007,34(6):497—509
[59]Li X Y,Li Z,Zhang X J,et al. Expression characterization of testicular DMRT1 in both Sertoli cells and spermatogenic cells of polyploid gibel carp [J]. Gene,2014,548(1):119—125
[60]Hale M C,Jackson J R,DeWoody J A. Discovery and evaluation of candidate sex-determining genes and xenobiotics in the gonads of lake sturgeon(Acipenser fulvescens) [J]. Genetica,2010,138(7):745—756
[61]Guo Y,Cheng H,Huang X,et al. Gene structure,multiple alternative splicing,and expression in gonads of zebrafish Dmrt1 [J]. Biochemical and Biophysical Research Communications,2005,330(3):950—957
[62]Johnsen H,Seppola M,Torgersen J S,et al. Sexually dimorphic expression of dmrt1 in immature and mature Atlantic cod(Gadus morhua L.) [J]. Comparative Biochemistry and Physiology Part B:Biochemistry and Molecular Biology,2010,156(3):197—205
[63]Fernandino J,Hattori R,Shinoda T,et al. Dimorphic expression of dmrt1 and cyp19a1(ovarian aromatase) during early gonadal development in pejerrey,Odontesthes bonariensis [J]. Sexual Development,2008,2(6):316—324
[64]Marchand O,Govoroun M,D'Cotta H,et al.DMRT1 expression during gonadal differentiation and spermatogenesis in the rainbow trout,Oncorhynchus mykiss [J]. Biochimica et Biophysica Acta(BBA)-Gene Structure and Expression,2000,1493(1):180—187
[65]Amberg J J,Goforth R,Stefanavage T,et al. Sexually dimorphic gene expression in the gonad and liver of shovelnose sturgeon(Scaphirhynchus platorynchus) [J]. FishPhysiology and Biochemistry,2010,36(4):923—932
[66]Liu Z,Zhang Y,Wang D. Studies on feminization,sex determination,and differentiation of the Southern catfish,Silurus meridionalis-a review [J]. Fish Physiology and Biochemistry,2010,36(2):223—235
[67]Su L,Zhou F,Ding Z,et al. Transcriptional variants of Dmrt1 and expression of four Dmrt genes in the blunt snout bream,Megalobrama amblycephala [J]. Gene,2015,573(2):205—215
[68]Ma L,Wang W,Yang X,et al. Characterization of the Dmrt1 gene in the black rockfish Sebastes schlegeli revealed a remarkable sex-dimorphic expression [J]. Fish Physiology and Biochemistry,2014,40(4):1263—1274
[69]úbeda M,Merlo M A,Ortiz J B,et al. Expression profiling of the sex-related gene Dmrt1 in adults of the Lusitanian toadfish Halobatrachus didactylus(Bloch and Schneider,1801) [J]. Gene,2014,535(2):255—265
[70]He C L,Du J L,Wu G C,et al. Differential Dmrt1 transcripts in gonads of the protandrous black porgy,Acanthopagrus schlegeli [J]. Cytogenetic and Genome Research,2003,101(3—4):309—313
[71]Liarte S,Chaves E,García A,et al. Testicular involution prior to sex change in gilthead seabream is characterized by a decrease in DMRT1 gene expression and by massive leukocyte infiltration [J]. Reproductive Biology and Endocrinoogyl,2007,5:20—35
[72]Xia W,Zhou L,Yao B,et al. Differential and spermatogenic cell-specific expression of DMRT1 during sex reversal in protogynous hermaphroditic groupers [J]. Molecular and Cellular Endocrinology,2007,263(1):156—172
[73]Jeong H B,Park J G,Park Y J,et al. Isolation and characterization of DMRT1 and its putative regulatory region in the protogynous wrasse,Halichoeres tenuispinis [J]. Gene,2009,438(1):8—16
[74]Huang X,Guo Y,Shui Y,et al. Multiple alternative splicing and differential expression of dmrt1 during gonad transformation of the rice field eel [J]. Biology of Reproduction,2005,73(5):1017—1024
[75]Kondo M,Froschauer A,Kitano A,et al. Molecular cloning and characterization of DMRT genes from the medaka Oryzias latipes and the platyfish Xiphophorus maculatus [J]. Gene,2002,295(2):213—222
[76]Sheng Y,Chen B,Zhang L,et al. Identification of Dmrt genes and their up-regulation during gonad transformation in the swamp eel(Monopterus albus) [J]. Molecular Biology Reports,2014,41(3):1237—1245
[77]Guan G,Kobayashi T,Nagahama Y. Sexually dimorphic expression of two types of DM(Doublesex/Mab-3)-domain genes in a teleost fish,the Tilapia(Oreochromis niloticus) [J]. Biochemical and Biophysical Research Communications,2000,272(3):662—666
[78]Cao J L,Yu J H,Cao Z M,et al. Temporal and spatial expression of DMO and DMT gene in Oreochromis aurea[J]. Journal of Fisheries of China,2007,31(2):129—136[曹謹玲,俞菊華,曹哲民,等. 奧利亞羅非魚DMO和DMT基因的時空表達特征分析. 水產學報,2007,31(2):129—136]
[79]Wen A,You F,Tan X,et al. Expression pattern of dmrt4 from olive flounder(Paralichthys olivaceus) in adult gonads and during embryogenesis [J]. Fish Physiology and Biochemistry,2009,35(3):421—433
[80]Li Q,Zhou X,Guo Y,et al. Nuclear localization,DNA binding and restricted expression in neural and germ cells of zebrafish Dmrt3 [J]. Biology of the Cell,2008,100(8):453—463
[81]Winkler C,Hornung U,Kondo M,et al. Developmentally regulated and non-sex-specific expression of autosomal dmrt genes in embryos of the Medaka fish(Oryzias latipes) [J]. Mechanisms of Development,2004,121(7):997—1005
[82]Zhou X,Li Q,Lu H,et al. Fish specific duplication of Dmrt2:characterization of zebrafish Dmrt2b [J]. Biochimie,2008,90(6):878—887
[83]Liu S,Li Z,Gui J F. Fish-specific duplicated dmrt2b contributes to a divergent function through Hedgehog pathway and maintains left-right asymmetry establishment function [J]. PloS One,2009,4(9):e7261
[84]Bradley K M,Breyer J P,Melville D B,et al. An SNP-based linkage map for zebrafish reveals sex determination loci [J]. G3:Genes,Genomes,Genetics,2011,1(1):3—9
[85]Ma W J,Vavre F,Beukeboom L. Manipulation of arthropod sex determination by endosymbionts:diversity and molecular mechanisms [J]. Sexual Development,2014,8(1—3):59—73
[86]Lezer Y,Aflalo E D,Manor R,et al. On the safety of RNAi usage in aquaculture:The case of all-male prawn stocks generated through manipulation of the insulin-like androgenic gland hormone [J]. Aquaculture,2015,435:157—166
[87]Ventura T,Manor R,Aflalo E,et al. Timing sexual differentiation:full functional sex reversal achieved in Macrobrachium rosenbergii through silencing of a single insulin-like gene [J]. Biology of Reproduction,2012,86(3):90,1—6
[88]Ventura T,Rosen O,Sagi A. From the discovery of the crustacean androgenic gland to the insulin-like hormone in six decades [J]. General and comparative endocrinology,2011,173(3):381—388
[89]Rosen O,Manor R,Weil S,et al. A sexual shift induced by silencing of a single insulin-like gene in crayfish:ovarian upregulation and testicular degeneration [J]. PloS One,2010,5(12):e15281
[90]Ventura T,Sagi A. The insulin-like androgenic gland hormone in crustaceans:From a single gene silencing to a wide array of sexual manipulation-based biotechnologies[J]. Biotechnology Advances,2012,30(6):1543—1550
[91]Yoshimoto S,Okada E,Umemoto H,et al. A W-linked DM-domain gene,DM-W,participates in primary ovary development in Xenopus laevis [J]. Proceedings of the National Academy of Sciences,2008,105(7):2469—2474
[92]Smith C A,Roeszler K N,Ohnesorg T,et al. The avian Z-linked gene DMRT1 is required for male sex determination in the chicken [J]. Nature,2009,461(7261):267—271
[93]Yang X L,Jiang H S,Yang N. Mechanism of avian sex determination and differentiation [J]. Hereditas,2012,34(4):407—411 [楊秀榮,蔣和生,楊寧. 鳥類性別決定與性別分化機制. 遺傳,2012,34(4):407—411]
[94]Qiu X H. Research progress of sex determination [J]. Biological Technology World,2014,11:114 [丘曉花. 性別決定的研究進展. 生物技術世界,2014,11:114]
RESEARCH PROGRESS OF DMRT GENES IN HYDROBIONTES
LI Fa-Jun1,FU Chun-Peng1and LUO Yong-Ju2
(1. Weifang University of Science and Technology,Shouguang 262700,China; 2. Guangxi Academy of Fishery Sciences,Nanning 530021,China)
Dmrt is a gene family that is homologous to Doublesex in Drosophila melanogaste and Mab-3 in Caenorhabditis elegans. Dmrt genes encode a large family of transcription factors including a characteristic conservative zinc finger DNA-binding domain(i.e.,DM domain). This ancient gene family has been identified in animal groups ranging from invertebrates to vertebrates. Their biological functions include sex determination and differentiation,construction of tissues and organs,and maintenance related functions. This paper reviews the recent progress of Dmrt genes in hydrobionts with theirs functions in line of evolution from invertebrates to vertebrates. We analyzed the course of sex determining gene for Dmrt genes.We further presented prospects of their future research in hydrobionts.
Dmrt gene; Dsx gene; Gonad; Crustacean; Fish; Hydrobiontes
Q173
A
1000-3207(2016)05-1068-10
10.7541/2016.138
2015-10-08;
2016-01-05
國家現代農業產業技術體系專項(CARS-49); 廣西十二五重大科技專項(桂科重14121004-2-2)資助 [Supported by China Agriculture Research System(No. CARS-49); Guangxi Science and Technology Research Program(No. 14121004-2-2)]
李法君(1976—),男,山東壽光人; 博士; 主要從事水產動物遺傳育種研究。E-mail:lifajun1976@163.com
羅永巨(1967—),男,廣西博白人,博士,研究員; 主要從事水產動物遺傳育種研究。E-mail:lfylzc123@163.com
