雞卵清蛋白基因調控序列的克隆與載體構建
2016-11-01 00:37:23朱志偉陳曉宇于福先潘建治
浙江農業學報 2016年3期
黃 菁,朱志偉,陳曉宇,于福先,潘建治
(浙江省農業科學院 畜牧獸醫研究所,浙江 杭州 310021)
?
雞卵清蛋白基因調控序列的克隆與載體構建
黃菁,朱志偉,陳曉宇,于福先,潘建治*
(浙江省農業科學院 畜牧獸醫研究所,浙江 杭州310021)
卵清蛋白(ovalbumin)基因在雞基因組中只有一對等位基因,卻能每天合成分泌多達2 g的蛋白,占據卵白蛋白質的50%以上,是外源基因載體表達調控構件的首選。該研究從輸卵管特異性啟動子方面著手,通過對卵清蛋白基因啟動子的篩選優化,找出啟動子增強因子的位置以及組織特異性區域。將卵清蛋白基因上游-922~-2 073和-2 801~-3 100區域平均分成12個長度約為150 bp的序列,分別插入到-921~+38序列的上游,成功構建12個系列表達載體,為進一步篩選短縮版優化啟動子提供材料;卵清蛋白基因第一內含子區域截斷成300 bp左右的迷你內含子序列,成功構建8個迷你內含子系列載體,為篩選優化的迷你內含子提供必要的材料;成功分離雞輸卵管上皮細胞并優化電轉染條件,通過熒光素酶活性檢測初步篩選出具有最強活性重組質粒pGL4-UP-1412和內含子重組質粒pGL4-mini-intron-3,同時推斷出若干包含增強子序列區域。
卵清蛋白基因;調控序列;載體;熒光素酶活性
卵清蛋白(Ovalbumin)在雞基因組中只有一對等位基因,卻能每天合成分泌多達2 g的蛋白,占據卵白蛋白質的50%以上,自然成為外源基因載體表達調控構件的首選。至今,以家禽輸卵管定位表達為目標的基因導入研究,絕大部分采用了卵清蛋白基因的調控序列。 然而,即便采用了卵清蛋白的調控序列,也不能高效誘導出外源蛋白的大量合成和分泌。迄今,在美日韓等國制備的生物反應器轉基因家禽卵中,外源蛋白占卵白蛋白的極少超過1‰。Lillico等[1]的研究中采用了卵清蛋白基因上游2.8 kb的片段,實現了迄今最成功的輸卵管特異表達,卵白中人干擾素含量達到平均38 mg·L-1,最高的個體也僅為426 mg·L-1。相比之哺乳動物細胞培養體系的水平(10 g·L-1左右)仍然存在相當差距。Kwon等[2]進一步縮短啟動子長度至1.35 kb用于轉基因鵪鶉,雖然保持了輸卵管特異性,但在卵白中表達重組人白細胞介素-1受體拮抗蛋白(rhIL1RN)的含量更低,僅為88.7~233.8 μg·L-1。Zhu等[3]采用卵清蛋白基因5′端上游長達7.5 kb和15 kb的調控序列制作嵌合體雞表達單克隆抗體,雖然表達量較高(750 mg·L-1),但輸卵管特異性不強,也超過了病毒載體允許插入的長度。
外源基因表達量的高低,除受到基因組插入位置效應等隨機因素的影響外,主要與轉基因載體中啟動子等元件的活性有關,單純截取卵清蛋白基因上游的調控序列片段,難以獲得根本性的改善。而利用基因敲入的方法向卵清蛋白基因座定點導入外源基因,或者將外源基因插入BAC克隆等包含卵清蛋白基因的長片段中,再導入到基因組,雖然有可能使遠距離存在的潛在調節元件發揮作用,獲得高效率特異性表達,但受到目前家禽轉基因技術的限制,尚難于在個體水平上付諸實施,無法獲得實驗驗證。
因此,在轉基因載體的有限長度內,對表達調控元件,特別是啟動子領域進行優化,是現實有效的途徑,也是國際性的研究熱點。本研究從輸卵管特異性啟動子方面著手,通過對卵清蛋白基因啟動子的篩選優化,找出啟動子增強因子的位置以及組織特異性區域,為下一步對增強區域和特異性區域的整合,制備出具有表達活性高、組織特異性強的優化型啟動子提供基礎材料。
1 材料與方法
1.1儀器和試劑
PCR儀(ABI 9902,美國應用生物系統公司);高速冷凍離心機(HITACHI CR21GIII,日本株式會社日立制作所);酶標儀(Molecular Devices SpectraMax M5,美國);倒置顯微鏡(Nikon ELIPSE Ti-U,日本尼康公司);電轉染儀(BTX ECM830,美國哈佛儀器旗下BTX公司);內切酶(TAKARA公司);DNA回收試劑盒[TIANGEN,天根生化科技(北京)有限公司];熒光素酶活性檢測系統(ONE-Glo Luciferase Assay system,Promega公司)。
1.2雞卵清蛋白基因上游調控序列系列變異載體構建
1.2.1雞卵清蛋白基因上游調控序列的克隆
參考GenBank中雞卵清蛋白基因組序列(NC_006089),利用生物軟件Primer Premier 5和DNAMAN設計添加了酶切位點的引物,PCR擴增系列目的片段。引物名稱和序列詳見表1。
將轉錄起始位點作為+1位置,本試驗克隆的序列按照其長度分別命名詳見表2。
以實驗室已有的pGL4-921作為模板,TransTaq High Fidelity(HiFi)PCR SuerMix II PCR Kit擴增目的片段X—Y。用引物(F-3100,R-2981),(F-3010,R-2891),(F-2920,R-2801),(F-2703,R-1916),(F-1937,R-1776),(F-1803,R-1636),(F-1659,R-1522),(F-1547,R-1387),(F-1412,R-1251),(F-1275,R-1122),(F-1148,R-1023),(F-1048,R-922)進行PCR擴增反應。PCR產物在2%瓊脂糖凝膠中進行電泳檢測,用DNA純化回收試劑盒(離心柱型,TIANGEN)進行割膠回收。
1.2.2雞卵清蛋白基因上游調控序列變異熒光素酶報告基因載體構建
表1引物序列及酶切位點
Table 1Sequences of primers and their restriction sites

引物名稱引物序列(5’—3’)Tm/℃F-3100GGGGTACCCAACTATGCAGGTATGCT-GACAA64R-2981GGGGTACCTATACTACACTCTTTC-CCAATGGAAGAF-3010GGGGTACCTGGTCTTCCATTGG-GAAAGAGT64R-2891GGGGTACCTTTCCTCTCCAAGAAG-TAGCCGF-2920GGGGTACCTAAACCACCGGCTACTTCTT-GG64R-2801GGGGTACCCCACATAGTGTCCTGATTC-CAGAF-2073GGGGTACCGACCTCCACTACCACAAC-TACCACAA67R-1916GGGGTACCACAGGCTGCTTCTCACCCT-GTGF-1937GGGGTACCCACAGGGTGAGAAGCAGC-CTGT64R-1776GGGGTACCAGCAACCATGCCATATATA-ATTAAGCATF-1803GGGGTACCATGCTTAATTATATATG-GCATGGTTGCT64R-1636GGGGTACCCTGACTTGTCAGACAGGCTC-CAGAF-1659GGGGTACCGGGGTACCTCTGGAGCCT-GTCTGACAAGTCAG66R-1522GGGGTACCAAGAGAGACCGTTTTG-GCTCTTACTAF-1547GGGGTACCTAGTAAGAGCCAAAACG-GTCTCTCTT64R-1387GGGGTACCTCTGTCATCTGGTGTCAAT-CAAACACF-1412GGGGTACCGTGTTTGATTGACAC-CAGATGACAGA64R-1251GGGGTACCAGAAAGTCTGAAAATCTT-GCCTGCTF-1275GGGGTACCAGCAG-GCAAGATTTTCAGACTTTCT64R-1122GGGGTACCTGAGGACTTAAGACAAGAT-GGTAGTGCF-1148GGGGTACCGCACTACCATCTTGTCTTA-AGTCCTCA64R-1023GGGGTACCCACTAGCTGAAGTTTAGT-CAGTGCCAF-1048GGGGTACCTGGCACTGACTAAACT-TCAGCTAG64R-922GGGGTACCACTGTTTGATTTAAAAAT-GAGGGAA
注:下劃線表示確切位點及保護堿基。表3同。
表2序列名稱及序列位置
Table 2Name and position of sequences
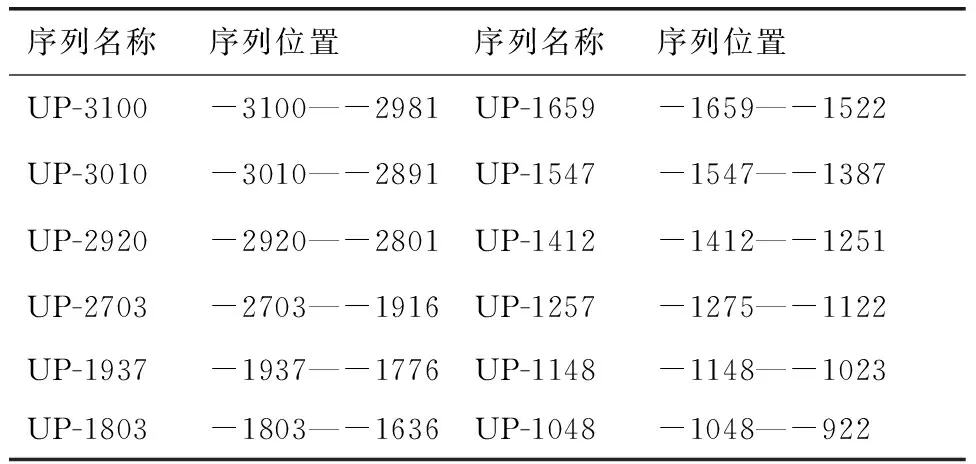
序列名稱序列位置序列名稱序列位置UP-3100-3100—-2981UP-1659-1659—-1522UP-3010-3010—-2891UP-1547-1547—-1387UP-2920-2920—-2801UP-1412-1412—-1251UP-2703-2703—-1916UP-1257-1275—-1122UP-1937-1937—-1776UP-1148-1148—-1023UP-1803-1803—-1636UP-1048-1048—-922
將回收得到的上游DNA片段進行Kpn Ⅰ酶切,酶切后產物用DNA純化回收試劑盒(離心柱型,TIANGEN)進行純化回收。將酶切后回收得到的UPs目的片段與pGL4-921雙酶切產物按照比例進行連接,然后轉化大腸埃希菌。采用菌液PCR法篩選出陽性菌落。挑選正向連接檢測為陽性的菌液送生工生物工程(上海)股份有限公司測序。
1.3迷你內含子系列熒光素酶報告載體構建
1.3.1卵清蛋白基因第一內含子全序列及系列分割序列克隆
參考GenBank中雞卵清蛋白基因組序列(NC_006089),利用生物軟件Primer Premier 5和DNAMAN設計8對添加了酶切位點的引物,PCR擴增系列包含或不包含內含子的目的片段。具體引物序列詳見表3。方法同上。
1.3.2迷你內含子系列載體構建
以純化回收得到的OV基因為模版,PCR擴增目的片段OV+38/OV+1640/OV-合成,膠回收法純化回收PCR產物。方法同上。
將OV+38/OV+1640/OV-合成的PCR產物進行Kpn Ⅰ,Xho Ⅰ雙酶切,同時將質粒pGL4.10 Vector進行Kpn Ⅰ,Xho Ⅰ雙酶切。37℃酶切2 h后,將每個酶切產物進行切膠回收、連接、轉化,挑選陽性克隆子測序鑒定。
1.4載體的表達活性檢測
1.4.1雞輸卵管上皮細胞分離培養與轉染
輸卵管上皮細胞的分離培養步驟如下:
(1)放血處死試驗雞,盡量將血放干凈,體內如果瘀血太多,輸卵管上也會存在大量血絲,分離上皮細胞時會有較多的紅細胞摻雜進來。
(2)將處死的雞置于托盤中,腹部朝上,去除腹部及雞右側腿部羽毛(去除之前可噴灑醫用乙醇,避免雞毛散飛),用滅菌剪刀剪開腹部及雞右側皮膚,換剪刀鑷子,小心剪開腹部肌肉,去除脂肪,取出輸卵管膨大部,置于超凈臺內準備好的PBS中。
(3)選擇較好的膨大部3~5 cm,用小剪刀、小鑷子小心去除膨大部外層的膜和血管。輸卵管內層為褶皺狀,去除外層系膜有利于褶皺部展開,消化時與胰酶充分接觸。此外,毛細血管一般分布在系膜與組織之間,去除系膜有利于去除血管,減少血細胞的污染。
表3PCR引物序列及酶切位點
Table 3Sequence of primers and their restriction sites

引物名稱產物名稱引物序列(5’—3’)Tm/℃OVFOVROV1GACTGACATGCATTTCATAGGTAGACTGTCTTTTGCACCCAGGT55OVR+38OVF-1338(1)OVR+1640intron-lessintron-containCCGCTCGAGCTTGAGTGCTAAAGGCAATACAGGGGGTACCGACTGACATGCATTTCATAGGTAGACCGCTCGAGGTTGTCTAGAGCAAACAGCAGA60OVF-1338(2)OVR-Sintron-deletionGGGGTACCGACTGACATGCATTTCATAGGTAGAGATA-ACATTTACTGGGAAGCACCCGCTCGAGGTTGTCTAGAGCAGAGCTCCTAGCTAGCAGTTGCTTAC-CTTTTGAGCTTGAGTGCT60OVF+57OVR+387miniintron-1CTAGCTAGCCTCTGGAATTACCTTCTCTCTATTAGCCGAGCTCACTGAACACCATGATTTTGCTC60OVF+368OVR+697miniintron-2CTAGCTAGCGCAAAATCATGGTGTTCAGTGACGAGCTCTTTATATAATCTGTGAAAGTCAGAAGTCCT60OVF+678OVR+1007miniintron-3CTAGCTAGCACTTTCACAGATTATATAAATCTCAGGAAAGCCGAGCTCGCACTTTCAGGTATGGTTTGC60OVF+987OVR+1317miniintron-4CTAGCTAGCAAACCATACCTGAAAGTGCAATAGAGCCGAGCTCGTACCTCTGATTAAAACACAATCTGATAGGC60OVF+1297OVR+1627miniintron-5CTAGCTAGCCAGATTGTGTTTTAATCAGAGGTACTGACGAGCTCAACAGCAGAACAGTGAAAATGTAAGG60

圖1 第一內含子分區示意圖Fig.1 The partition schematic of the first intron
(4)將血管、系膜去除干凈的輸卵管部分翻折,內層朝外。利用小剪刀的把手進行刮除附著的黏液成分,在PBS中進行徹底的清洗。(去除膜以后的輸卵管組織特別薄,不能用尖銳利器進行刮除)。
(5)將清洗干凈的組織整塊至于適量0.2%膠原酶中,水浴鍋中預熱5 min,然后于37℃搖床85 r·min-1消化5~8 min,250 g離心4 min,去除上清,PBS清洗2遍,離心去上清。
(6)加入3倍體積的胰酶37℃搖床85 r·min-1消化10~15 min,超凈臺內吹打數次,取少量消化液顯微鏡下觀察細胞數量。若細胞數較少,可適當延長消化時間,若能夠觀察到大量密布細胞時即可終止消化。
(7)將吹打數次的消化液于200目篩過濾,將濾液收集入新的50 mL離心管中,加入適量血清終止胰酶的消化,250 g離心4 min,去除上清。PBS清洗2次,離心去上清。
(8)加入適量PBS進行洗滌,250 g離心4 min后去除上清,加入適量含10%胎牛血清、1% 雙抗的DMEM重懸,混勻后置于10 cm培養板中,37℃,5%二氧化碳培養,24 h后顯微鏡下觀察細胞貼壁情況。
細胞轉染參照lipofecatmine 2000(Thermo Fisher)試劑使用說明書。
1.4.2熒光素酶活性測定
利用ONE-Glo Luciferase Assay system(Promega)對不同重組表達載體在細胞中的熒光素酶表達進行測定。試驗步驟如下:
(1)提前將光柵式多功能酶標儀開啟,設定化學發光測定程序。
(2)準備進行熒光素酶活性測定的細胞去除培養液,PBS清洗2~3遍。
(3)向每個孔內加入100 μL PBS+150 μL ONE-Glo反應液,避光,室溫條件下輕輕晃動培養板將細胞充分裂解(至少3 min)。
(4)將24孔板中的細胞裂解液吹打數次,每孔取200 μL裂解液吸取至白色96孔酶標板。
(5)將加入裂解液的96孔酶標板放入多功能酶標儀,測定熒光素酶活性。
2 結果與分析
2.1雞卵清蛋白基因上游調控序列系列變異載體構建成功
2.1.1pGL4-UPS表達載體菌液PCR陽性結果
上游調控序列各片段PCR產物回收之后,12個片段均介于120~200 bp之間,符合預期大小。所有重組質粒的菌液PCR檢測以pGL4.10載體上的pGL3+(TAGCAAAATAGGCTGTCCC)為上游引物,OVsR為下游引物,12個重組載體中的目的片段大小存在一定差異,所以經PCR擴增后,正確的連接將會分別擴增出介于200~300 bp之間,約為220 bp左右的片段。圖2顯示為其中一個表達載體PGL4-UP-2073菌液PCR呈陽性,所有測序驗證結果表明該載體構建成功。
2.1.2pGL4-UPS表達載體構建的酶切鑒定成功
菌液鑒定為陽性的質粒經過測序驗證,選取測序無突變克隆子提取質粒,Kpn Ⅰ,EcoRⅤ雙酶切鑒定。所有質粒總長度均為5 000 bp左右,經Kpn I,EcoRⅤ雙酶切后,將質粒切成長度分別為4 020,921和120~160 bp的3部分。由圖3可見,所有質粒的酶切檢測均獲得預期片段,質粒構建成功。泳道1,4,7,10,13,16,19,22,25,28,31,34為構建的12個重組質粒。每個質粒后的2個泳道分別為相應質粒的限制性內切酶EcoRⅤ的單酶切和Kpn Ⅰ,EcoRⅤ雙酶切電泳圖。

圖2 pGL4-UP-2073正連接菌液PCR檢測結果Fig.2 PCR result of the positive ligation of pGL4-UP-2073 plasmid
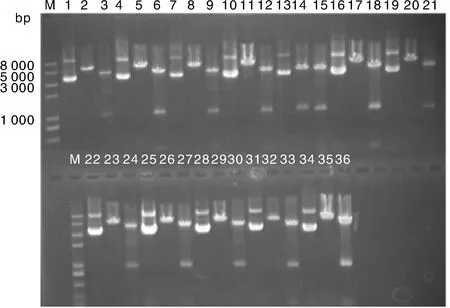
泳道1,4,7,10,13,16,19,22,25,28,31,34為構建的12個重組質粒;泳道2,5,8,11,14,17,20,23,26,29,32,35為各質粒經由EcoR Ⅴ的單酶切;泳道3,6,9,12,15,18,21,24,27,30,33,36為各質粒經由Kpn Ⅰ,EcoR Ⅴ雙酶切。圖3 pGL4-UPs質粒酶切鑒定結果Fig.3 Results of restriction enzyme digestion for pGL4-UPs plasmids
2.2迷你內含子系列載體成功構建
2.2.1卵清蛋白基因第一內含子序列成功克隆
利用PCR方法克隆雞卵清蛋白基因第一內含子-1 338~+1 798位置的目的片段,純化回收后電泳圖。由圖4可見,目的基因長度為3 136 bp,目的條帶的位置符合預期,克隆成功。
2.2.2基礎載體及迷你內含子系列載體成功構建
經過菌液PCR陽性篩選和測序驗證之后,將無序列突變質粒進行酶切鑒定,如圖5所示,所有質粒的酶切檢測均獲得預期片段,質粒構建成功。泳道1,4,7,10,13,16為構建成功的質粒,每個質粒后的2個泳道分別為相應質粒的限制性內切酶EcoRⅤ單酶切和Kpn Ⅰ,EcoRⅤ雙酶切電泳圖。
2.3載體的表達活性效果驗證
2.3.1雞輸卵管上皮細胞培養狀態良好
由圖6可見,本試驗成功地從產蛋母雞的輸卵管分離到上皮細胞,呈現出良好的上皮細胞形態,成纖維細胞較少。
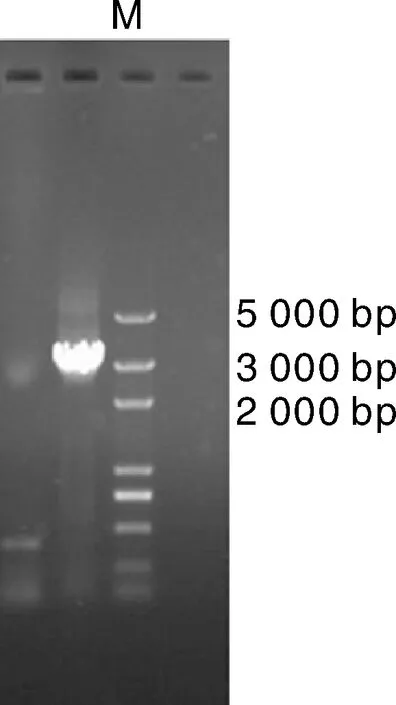
圖4 第一內含子序列克隆回收后凝膠電泳圖Fig.4 Gel electrophoresis of the first-intron sequence after cloning and purification
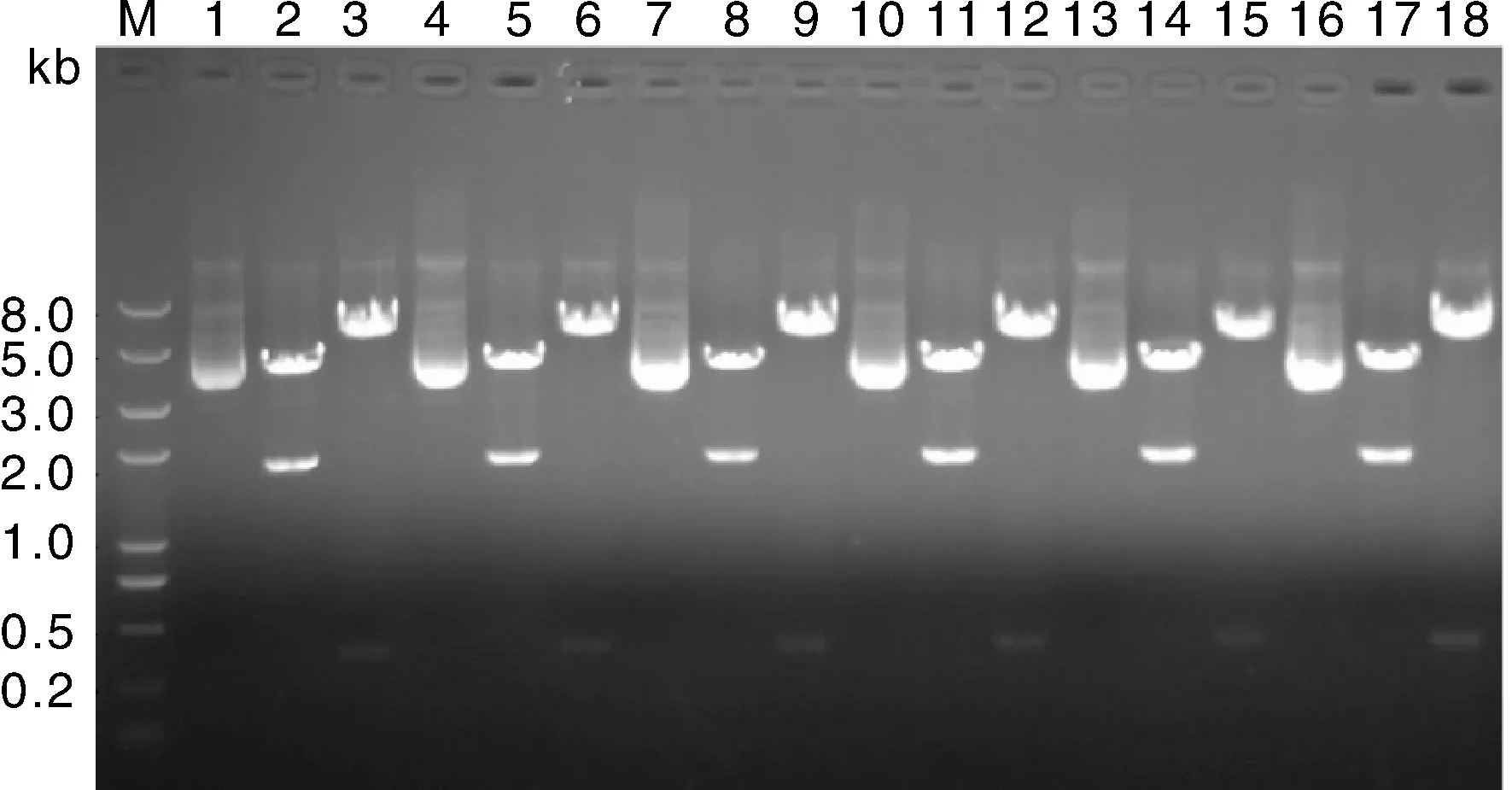
泳道1,4,7,10,13,16為構建成功的質粒;泳道2,5,8,11,14,17為各質粒經由EcoRV單酶切;泳道3,6,9,12,15,18為個質粒經由KpnI,EcoRV雙酶切。圖5 pGL4-mini-introns質粒酶切鑒定結果Fig.5 Results of restriction enzyme digestion for pGL4-mini-introns plasmids
圖6 雞輸卵管上皮細胞形態Fig.6 Cell morphology of chicken oviduct epithelial cell
2.3.2確定最優轉染條件
根據有關文獻,將電轉染電壓初步設定為250 V,利用293T細胞進行電轉染時間的優化,將電擊時間設1,3,5,7,9,11 ms等6個實驗組,結果顯示在同等電壓下,7 ms以上細胞死亡率達30%以上,1 ms時,轉染效率很低,在保證細胞成活率的前提下,選擇5 ms作為最佳電擊時間。根據此結果,在輸卵管上皮細胞上進行電轉染條件的進一步優化。以250 V,5 ms為基礎,固定電擊時間,設150,200,250,300,350 V等5個實驗組進行探究。實驗結果顯示:300 V,5 ms條件時,相較于低電壓電轉條件,死亡細胞數稍多,懸浮于培養液中,轉染效率高,可達20%左右,如圖7所示,而電壓為350 V時細胞死亡數過多,不利于后續試驗,因此確定最佳電轉染條件為300 V,5 ms。

圖7 雞輸卵管上皮細胞電轉染效果Fig.7 Electric transfection efficiency of chicken oviduct epithelial cell
2.3.3卵清蛋白基因上游調控序列表達活性
熒光素酶檢測結果顯示,重組質粒pGL4-UP-1412表達活性最強(圖8),表明在雞卵清蛋白基因上游-1 412~-1 251 bp序列部分可能存在有效調控元件。
2.3.4迷你內含子系列載體表達活性
熒光素酶檢測結果顯示,內含子重組質粒pGL4-mini-intron-3表達活性最強(圖9),說明在內含子+678~+1 008位置存在對轉錄活性有增強作用的區域。整體比較pGL4-mini-intron-1,pGL4-mini-intron-2,pGL4-mini-intron-3,pGL4-mini-intron-4和pGL4-mini-intron-5,結果顯示,表達活性呈先降后升的趨勢,表明在+1 298~+1 628區域可能存在表達抑制性因子。同時推斷出若干包含增強子序列區域。
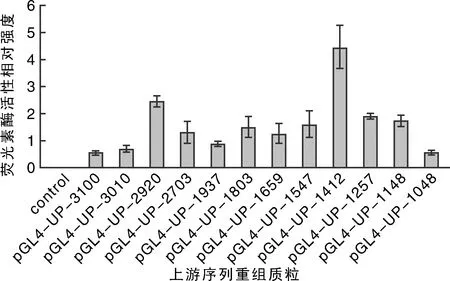
圖8 pGL4-Ups質粒熒光素酶活性檢測結果Fig.8 Luciferase activity of pGL4-UPs plasmids
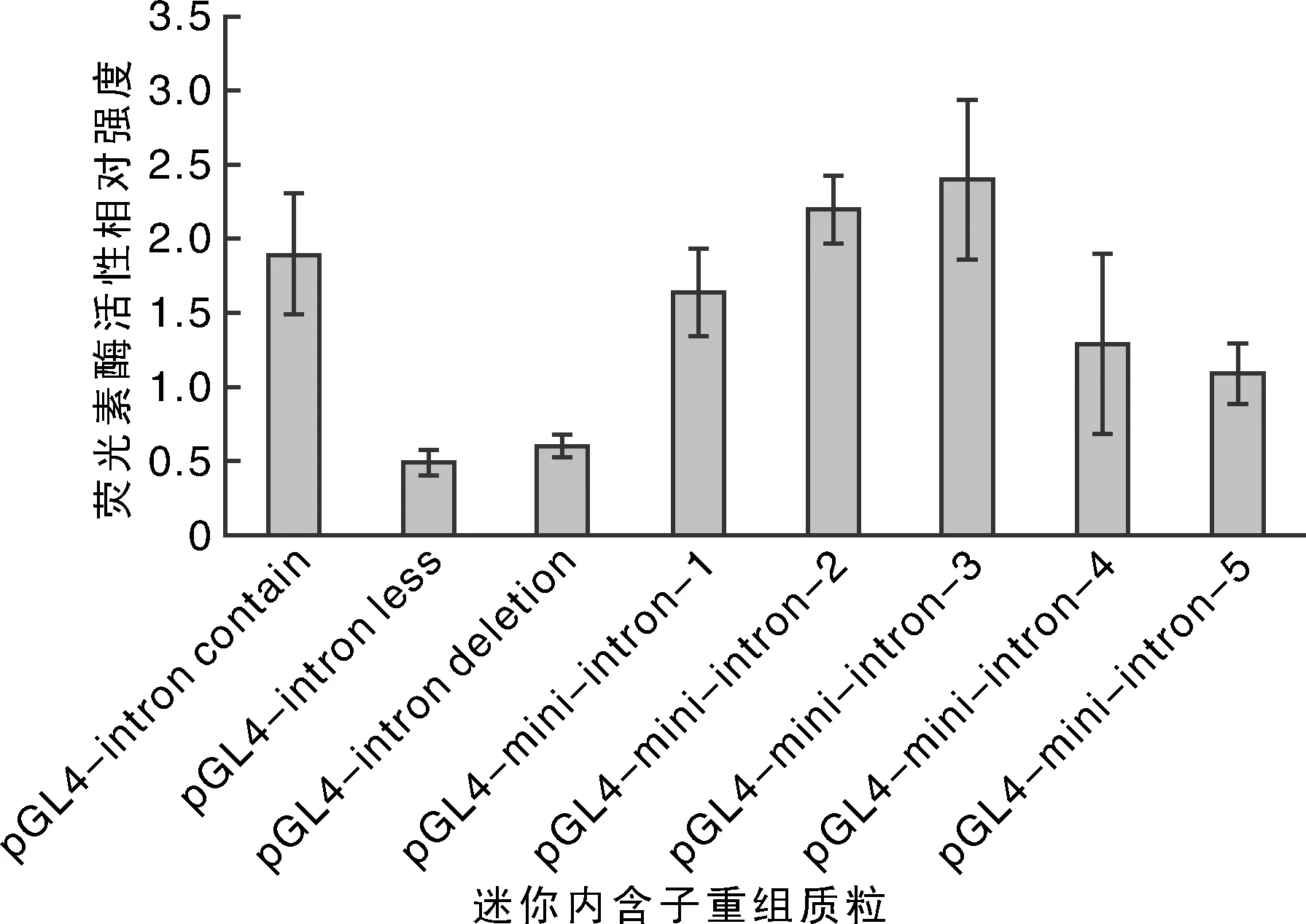
圖9 pGL4-mini intron質粒熒光素酶活性檢測結果Fig.9 Luciferase activity of pGL4-mini intron plasmids
3 結論與討論
與哺乳動物相比,禽蛋作為表達重組蛋白的載體,具有多方面顯著優勢:家禽繁殖周期短,能在較短周期內建群;家禽飼養成本低、產蛋量高,一段濕質量50 g的產卵期輸卵管膨大部每天能產生4 g蛋白,蛋白合成分泌的效率驚人;卵白中的蛋白不會漏入血液循環,危害動物本身的健康;禽蛋蛋白成分簡單,具有天然無菌環境,穩定性好,便于提取純化;輸卵管細胞能表達糖蛋白和磷蛋白等復雜結構的蛋白質,結構較牛羊乳腺更接近于人[4];SPF飼養技術成熟,產品安全性高等。以上特點使家禽被視為生產重組蛋白的理想生物反應器體系,其生產成本有可能降至細胞培養法的1%以下,也大大低于轉基因牛羊。蛋白中的蛋白質穩定,藥物蛋白在雞蛋中有比較長的半衰期,因此禽類輸卵管生物反應器以其卓越的優勢必將成為生物科學領域研究的熱點之一,必將成為研究和投資的重點,成為醫療性蛋白生產的理想途徑。
商業化的載體所容納外源基因具有一定的長度限制,因此,在表達載體的有限長度內,對表達調控元件進行優化,是一種現實有效的途徑,也是國際性的研究熱點。此外,外源基因的表達量高低,除受到基因組插入位置效應的影響外,主要與轉基因在體內啟動子等元件的活性有關,單純截取卵清蛋白基因上游的調控序列片段,難以獲得根本性的改善。在本試驗結果的基礎上,制備出具有表達活性高、組織特異性強的優化型啟動子,調控蛋白基因的高效表達,從而解決載體容納量低和外源基因在體內表達量低的問題。
生物反應器可以生產種類繁多的有用蛋白,包括單克隆抗體、疫苗、激素、生長因子、酶、血清蛋白等,這些大多難于從自然界原料中分離提取,或者自然界根本不存在。微生物(大腸埃希菌和酵母)發酵表達系統是目前最為常見的基因工程蛋白生產手段,但對于眾多具有復雜結構的蛋白而言,難于從微生物獲得正確表達,立體構造和翻譯后修飾狀態的不同,影響到蛋白質的功能。因此,需要借助高等動物細胞培養系統或動物個體來進行重組蛋白生產。
本研究成功獲得pGL4-UP-2703,pGL4-UP-1937,pGL4-UP-1803,pGL4-UP-1659,pGL4-UP-1547,pGL4-UP-1412,pGL4-UP-1257,pGL4-UP-1148,pGL4-UP-1048,pGL4-UP-3100,pGL4-UP-3010,pGL4-UP-2920等12種雞卵清蛋白上游調控序列重組報告基因載體;及pGL4-intron-contain,pGL4-intron-less,pGL4-intron-delation,pGL4-mini-intron-57,pGL4- mini-intron-368,pGL4- mini-intron-678,pGL4- mini-intron-988,pGL4- mini-intron-198等8種雞卵清蛋白第一內含子迷你化重組報告基因載體;成功分離培養了雞輸卵管原代上皮細胞,并優化電轉染條件,為后期實驗研究提供了有用的實驗數據;所構建的載體均具有調控熒光素酶基因表達的活性。通過比較UPs等20種序列的啟動子活性,為找出存在于雞卵清蛋白基因上游調控序列中及第一內含子中的增強子、抑制子等功能性區域提供了新的數據,但要制作出高活性、高特異性的啟動子還須進一步探索研究。
[1]LILLICO S G,SHERMAN A,MCGREW M J,et al.Oviduct-specific expression of two therapeutic proteins in transgenic hens [J].Proceedings of the National Academy of Sciences of the United States of America,2007,104(6):1771-1776.
[2]KWON S C,CHOI J W,JANG H J,et al.Production of biofunctional recombinant human interleukin 1 receptor antagonist (rhIL1RN) from transgenic quail egg white [J].Biology of Reproduction,2010,82(6):1057-1064.
[3]ZHU L,VAN DE LAVOIR M C,ALBANESE J,et al.Production of human monoclonal antibody in eggs of chimeric chickens [J].Nature Biotechnology,2005,23(9):1159-1169.
[4]PALEYANDA R K,VELANDER W H,LEE T K,et al.Transgenic pigs produce functional human factor Ⅷ in milk [J].Nature Biotechnology,1997,15(10):971-975.
(責任編輯盧福莊)
本刊目前僅刊載黑白圖,請勿使用彩圖,圖片的分辨率不能低于600 dpi。圖中縱橫軸標值要求圓整化且成等差,標值線一律朝內。柱狀圖先用不同灰度加以區別;線圖圖例優先使用○●△▲◇◆。半欄圖的寬度宜控制在6.0~7.5 cm,且圖寬度和高度比以3∶2為宜;通欄圖的寬度宜控制在15.0 cm以內。相近內容的圖最好組合在一起作為一圖出現,各小圖可分別用大寫英文字母A,B,C等標識。
4.10參考文獻
參考文獻應只列出與本研究切實相關,在文稿中實際引用的文獻。參考文獻采用順序編碼制,即按在文中引用的先后順序編輯和著錄。請盡量選用新近發表在權威學術期刊上的文獻。所有參考文獻均須按本刊格式譯成英文(中文文獻須英漢對照)。無論中英文署名,一律姓先名后,對于外文作者,姓全部大寫,名用縮寫、大寫,但不加縮寫點。其他參考文獻格式要求同GB/T 7714—2015《信息與文獻 參考文獻著錄規則》。
例:
[1]CHANEY R L,MALIK M,YIN M L,et al.Phytoremediation of soil metals[J].Current Opinion in Biotechnology,1997,8(3):279-284.
(外文期刊)
[2]蔣明金,馬均,孫永健,等.播種量和氮肥運籌對直播雜交稻光合生產力及氮素利用的影響[J].浙江大學學報(農業與生命科學版),2015,41(5):516-526.DOI:10.3875/j.issn.1008-9209.2015.07.101.(in Chinese with English abstract)
JIANG M J,MA J,SUN Y J,et al.Effects of seeding rates and nitrogen fertilizer managements on photosynthetic productivity and nitrogen utilization in direct-seeded rice[J].Journal of Zhejiang University (Agric.&Life Sci.),2015,41(5):516-526.DOI:10.3875/j.issn.1008-9209.2015.07.101.(in Chinese with English abstract)
(帶英文摘要的中文期刊)
[3]朱月清.有機肥施用量對瓠瓜浙蒲6號生長的影響[J].浙江農業科學,2015 (10):1561-1562.
ZHU Y Q.Effects of organic fertilizer on growth of bottle gourd cv.Zhepu No.6[J].Journal of Zhejiang Agricultural Sciences,2015 (10):1561-1562.(in Chinese)
(中文期刊)
[4]畢艷蘭.油脂化學[M].北京:化學工業出版社,2005:70,78.
BI Y L.Oil chemistry[M].Beijing:Chemical Industry Press,2005:70,78.(in Chinese)
(中文圖書)
[5]田生科.超積累東南景天(Sedum alfredii Hance)對重金屬(Zn/Cd/Pb)的解毒機制[D].杭州:浙江大學,2010.
TIAN S K.Mechanisms behind detoxification of heavy metals (Zn/Cd/Pb) in the hyperaccumulator Sedum alfredii Hance[D].Hangzhou:Zhejiang University,2010.(in Chinese with English abstract)
(學位論文)
4.11謝辭
如有需要致謝的內容,可在正文與參考文獻之間另起一段,不要寫在首頁的題注中。謝辭要簡明扼要,僅對研究工作或論文撰寫有實際幫助(資助)者致謝。
Cloning and vector construction of chicken ovalbumin gene regulatory sequences
HUANG Jing,ZHU Zhi-wei,CHEN Xiao-yu,YU Fu-xian,PAN Jian-zhi*
(Animal Husbandry and Veterinary Research Institute,Zhejiang Academy of Agricultural Sciences,Hangzhou 310021,China)
There was only one allele of ovalbumin gene in the chicken genome,but it could synthesize and secrete 2 g protein per day,which were accounting for more than 50% of the albumin protein and became the preferred choice in the regulation of exogenous gene expression.This study aimed to identify promoter enhancer and tissue-specific regional location factor by screening the optimization of ovalbumin gene promoter.The upstream -922~-2 073 and -2 801~-3 100 of ovalbumin promoter were divided into 12 regional which average sequence length were about 150 bp,and inserted into the upper reaches of -921~+38 sequences,those 12 series successfully constructed expression vectors provided materials for further optimization promoter with a shortened version.The first intron region of ovalbumin promoter was truncated around 300 bp of mini intron sequences,and successfully constructed 8 mini intron series of vectors.We also successfully separated chicken oviduct epithelial cells and optimized the electricity transfection conditions.In this study,the promoter region with strongest activity pGL4-UP-1412 and pGL4-mini-intron-3 were screened through detected luciferase activity of the initial screening recombinant plasmid and intron recombinant,while inferred several regions containing enhancer sequence.
ovalbumin gene;regulation sequences;vector;luciferase activity
浙江農業學報Acta Agriculturae Zhejiangensis,2016,28(3):412-419http://www.zjnyxb.cn
黃菁,朱志偉,陳曉宇,等.雞卵清蛋白基因調控序列的克隆與載體構建[J].浙江農業學報,2016,28(3):412-419.
10.3969/j.issn.1004-1524.2016.03.09
2015-09-15
浙江省自然科學基金資助項目(LQ12C17002)
黃菁(1983—),女,浙江舟山人,博士,助理研究員,從事動物生物工程研究。E-mail:xiaojingyu1102@gmail.com
,潘建治,E-mail:jzpan9@126.com
S831,Q785
A
1004-1524(2016)03-0412-08
2016-01-08)
