不同品種鴨胚胎期骨骼肌成肌細(xì)胞GHR和IGF-1 mRNA表達(dá)差異分析
2016-11-01 00:37:23姬改革陶志云朱春紅束婧婷劉宏祥徐文娟李慧芳
浙江農(nóng)業(yè)學(xué)報(bào) 2016年3期
關(guān)鍵詞:差異
姬改革,陶志云,朱春紅,束婧婷,劉宏祥,徐文娟,胡 艷,李慧芳
(江蘇省家禽科學(xué)研究所,江蘇省家禽遺傳育種重點(diǎn)實(shí)驗(yàn)室,江蘇 揚(yáng)州 225003)
?
不同品種鴨胚胎期骨骼肌成肌細(xì)胞GHR和IGF-1 mRNA表達(dá)差異分析
姬改革,陶志云,朱春紅,束婧婷,劉宏祥,徐文娟,胡艷,李慧芳*
(江蘇省家禽科學(xué)研究所,江蘇省家禽遺傳育種重點(diǎn)實(shí)驗(yàn)室,江蘇 揚(yáng)州 225003)
為探討生長(zhǎng)激素受體(growth hormone receptor,GHR)和胰島素樣生長(zhǎng)因子-1(Insulin-like growth factor-1,IGF-1)在不同品種鴨胚胎期骨骼肌成肌細(xì)胞中的表達(dá)差異。選擇生長(zhǎng)速度不同的金定鴨和高郵鴨為試驗(yàn)動(dòng)物,采用實(shí)時(shí)熒光定量PCR方法檢測(cè)鴨13,15,17,19,21和23胚齡時(shí)胸肌和腿肌成肌細(xì)胞GHR,IGF-1 mRNA的表達(dá)情況,并進(jìn)行了品種間比較。結(jié)果發(fā)現(xiàn):13胚齡時(shí),高郵鴨胸肌和腿肌成肌細(xì)胞IGF-1的表達(dá)均顯著高于金定鴨(P<0.05);17胚齡是兩個(gè)品種鴨GHR和IGF-1共同的表達(dá)高峰期;17胚齡之后,鴨胸肌和腿肌成肌細(xì)胞GHR和IGF-1表達(dá)均顯著降低。在19,21胚齡時(shí),高郵鴨胸肌成肌細(xì)胞2個(gè)基因的表達(dá)均顯著高于金定鴨(P<0.05),而腿肌成肌細(xì)胞GHR和IGF-1表達(dá)在品種間無(wú)顯著差異(P>0.05)。鴨胚胸肌和腿肌成肌細(xì)胞GHR和IGF-1表達(dá)呈現(xiàn)極顯著的正線性相關(guān)(P<0.01)。提示:GHR和IGF-1的表達(dá)譜及各自在品種間的變化規(guī)律基本一致;鴨胚骨骼肌成肌細(xì)胞GHR和IGF-1 mRNA表達(dá)有組織特異性,二者之間可能存在正調(diào)控的作用機(jī)制。
鴨;胚胎;成肌細(xì)胞;GHR;IGF-1
生長(zhǎng)激素受體(GHR)和胰島素樣生長(zhǎng)因子-1(IGF-1)在動(dòng)物骨骼肌發(fā)育過(guò)程中起著重要的調(diào)控作用。動(dòng)物的生長(zhǎng)受到生長(zhǎng)軸“下丘腦—垂體—靶器官”上的激素及受體組成的神經(jīng)內(nèi)分泌系統(tǒng)的調(diào)控,生長(zhǎng)激素(growth hormone,GH)與IGF-1都是生長(zhǎng)軸上的重要因子。GH處于生長(zhǎng)軸的中心環(huán)節(jié),可以通過(guò)與肝臟或肝外組織細(xì)胞膜表面的GHR結(jié)合,介導(dǎo)產(chǎn)生IGF-1,IGF-1以內(nèi)分泌或旁分泌與骨骼肌上的受體結(jié)合,激活下游的信號(hào)因子,啟動(dòng)基因表達(dá),促進(jìn)蛋白質(zhì)合成、細(xì)胞分裂等;或者不通過(guò)IGF-1[1],直接作用于骨骼肌上的GHR發(fā)揮其作用。
肌肉的基本單位是肌纖維,是由成肌細(xì)胞增殖、分化與融合形成。胚胎期成肌細(xì)胞的增殖與分化能力的大小對(duì)肌肉產(chǎn)量的提高具有重要意義[2-3]。關(guān)于GH對(duì)成肌細(xì)胞的作用,目前主要集中在哺乳動(dòng)物。在小鼠中,體內(nèi)和體外的實(shí)驗(yàn)表明,GH可以通過(guò)介導(dǎo)IGF-1促進(jìn)成肌細(xì)胞的增殖與融合[4-6];但也有研究認(rèn)為,GHR可以不通過(guò)IGF-1調(diào)控成肌細(xì)胞的增殖與分化[7]。在雞上,體內(nèi)體外的試驗(yàn)表明[8-9],IGF-1可促進(jìn)成肌細(xì)胞的增殖與分化。鴨作為水禽的代表,其生長(zhǎng)發(fā)育與哺乳動(dòng)物、雞等具有不同的特點(diǎn)。有關(guān)GHR和IGF-1在鴨成肌細(xì)胞的表達(dá)變化規(guī)律尚未見(jiàn)報(bào)道。
在前期研究中我們也已經(jīng)建立一套完善的分離與培養(yǎng)體系[10],本試驗(yàn)以高郵鴨和金定鴨為研究對(duì)象,提取不同時(shí)間點(diǎn)鴨胚骨骼肌成肌細(xì)胞,采用實(shí)時(shí)熒光絕對(duì)定量分析技術(shù),對(duì)GHR和IGF-1的mRNA表達(dá)量進(jìn)行比較分析,初步揭示鴨胚胎期骨骼肌成肌細(xì)胞中GHR和IGF-1基因表達(dá)的品種差異和時(shí)空特點(diǎn),為提高鴨的肌肉產(chǎn)量,和品種選育等提供可借鑒的遺傳學(xué)資料。
1 材料與方法
1.1實(shí)驗(yàn)動(dòng)物
在相同的日糧水平下,收集高郵鴨和金定鴨種蛋。種蛋孵化前,進(jìn)行稱量、消毒和編號(hào),品種內(nèi)蛋質(zhì)量變異系數(shù)在2%以內(nèi)。2個(gè)品種種蛋隨機(jī)置入同一孵化箱,在相同條件下同時(shí)孵化。種蛋入孵后的24 h設(shè)定為1胚齡,分別于6個(gè)時(shí)間點(diǎn)(13,15,17,19,21和23胚齡)取5~8個(gè)鴨胚,用于胸肌和腿肌成肌細(xì)胞的提取。
1.2主要試劑和儀器
DMEM和FBS購(gòu)于Hyclone公司;胰蛋白酶,Ⅰ型膠原酶購(gòu)自Sigma公司;青霉素-鏈霉素溶液購(gòu)自碧云天生物技術(shù)研究所,Percoll細(xì)胞分離液購(gòu)自北京索萊寶科技有限公司。TRNzol-A+總RNA提取試劑、SuperReal PreMix、Quant cDNA 第一鏈合成試劑盒、pGM-T克隆試劑盒、質(zhì)粒小提試劑盒、DNA產(chǎn)物純化回收試劑盒購(gòu)自TIANGEN公司;DNA Marker DL2000為T(mén)aKaRa公司產(chǎn)品。倒置顯微鏡(日本尼康公司);9700PCR儀和Max3000P熒光定量PCR儀(愛(ài)普拜斯應(yīng)用生物系統(tǒng)上海有限公司);凝膠成像系統(tǒng)(Tanon 2500);紫外分光光度計(jì)(GeneQuant Ⅱ,Pharmacia Biotech)。
1.3鴨胚胎期骨骼肌原代成肌細(xì)胞的提取
無(wú)菌分離鴨胚胸肌和腿肌,用PBS清洗,去除脂肪及筋膜后,將肌肉剪為肉糜,加入1倍體積的混合酶(膠原酶和胰蛋白酶)消化5~10 min,用FBS終止消化。被消化下來(lái)的細(xì)胞經(jīng)不同孔徑的濾膜(50目~400目)過(guò)濾后,500 g離心10 min,去上清,細(xì)胞團(tuán)重懸在DMEM培養(yǎng)基中。將重懸的細(xì)胞液加入到含60%,20% Percoll的不連續(xù)密度梯度Percoll柱中,800 g,離心25 min。小心取出60%和20% Percoll兩層之間的云霧狀細(xì)胞至新的離心管。添加少量DMEM并混勻,500 g離心10 min,棄上清,用適量DMEM液懸浮,吸管吹吸,將細(xì)胞團(tuán)塊吹散,計(jì)數(shù)后將所得成肌細(xì)胞調(diào)整單細(xì)胞懸液密度為106cells·mL-1。
1.4總RNA提取和cDNA合成
將所得的單細(xì)胞懸液分裝至2 mL離心管中,每管1 mL,每個(gè)時(shí)間點(diǎn)分裝6~8管。500 g離心5 min后,去上清。將所得的細(xì)胞沉淀加入1 mL Trizol裂解液,保存在-20℃,用于總RNA提取。成肌細(xì)胞總RNA 提取,按總RNA提取試劑盒的說(shuō)明書(shū)進(jìn)行。所得的RNA用DEPC水溶解后,用1.4%瓊脂糖凝膠電泳和紫外分光光度計(jì)檢測(cè)純度和含量,保證RNA樣品質(zhì)量可靠,計(jì)算樣品總RNA濃度。經(jīng)檢測(cè)合格的總RNA樣品,取1 μg按照反轉(zhuǎn)錄試劑盒的使用說(shuō)明書(shū)進(jìn)行cDNA 第1鏈的合成。用內(nèi)參基因β-actin檢測(cè)cDNA合成質(zhì)量以及是否有基因組DNA污染。RT產(chǎn)物保存在-20℃用于PCR檢測(cè)。
1.5引物設(shè)計(jì)、目的片段標(biāo)準(zhǔn)品的制備
根據(jù)GenBank中相關(guān)序列設(shè)計(jì)引物,由上海英駿生物工程有限公司合成(表1)。PCR產(chǎn)物經(jīng)2%瓊脂糖凝膠鑒定后,用DNA產(chǎn)物純化回收試劑盒純化回收目的片段,與pGM-T載體相連接,然后轉(zhuǎn)化Escherichia coli TOP10感受態(tài)細(xì)胞,挑取轉(zhuǎn)化子于含氨芐抗性的LB液體培養(yǎng)基中,37℃,200 r·min-1振搖培養(yǎng)過(guò)夜,用PCR鑒定。將鑒定正確的質(zhì)粒送上海生工生物工程技術(shù)服務(wù)有限公司進(jìn)行序列測(cè)定,所獲序列結(jié)果在NCBI網(wǎng)站中用Blast程序與數(shù)據(jù)庫(kù)中公布的已知基因進(jìn)行序列同源性比較。比對(duì)正確后,用質(zhì)粒小提試劑盒提取陽(yáng)性克隆的質(zhì)粒,用分光光度計(jì)測(cè)其濃度后作為標(biāo)準(zhǔn)品備用。
1.6熒光實(shí)時(shí)定量PCR
所用熒光定量PCR采用SYBR Green Ⅰ法。將上述經(jīng)測(cè)序驗(yàn)證后正確的含GHR,IGF-1基因的標(biāo)準(zhǔn)質(zhì)粒,分別做10n梯度稀釋。將每個(gè)待測(cè)樣品RT產(chǎn)物取等體積混合,用混合樣(cDNA mix)和梯度稀釋的標(biāo)準(zhǔn)品進(jìn)行反應(yīng)條件的優(yōu)化,包括標(biāo)準(zhǔn)品稀釋梯度范圍、目的基因和內(nèi)參基因引物設(shè)計(jì)和合成、最佳退火溫度、引物濃度、模板濃度等,確定好最佳反應(yīng)條件。根據(jù)最佳反應(yīng)條件,將待測(cè)樣品進(jìn)行稀釋。將稀釋后的樣品與梯度稀釋的標(biāo)準(zhǔn)品在同一個(gè)試驗(yàn)中進(jìn)行定量PCR,每次反應(yīng)均設(shè)陰性對(duì)照,每個(gè)樣品設(shè)置3個(gè)重復(fù)。根據(jù)標(biāo)準(zhǔn)品構(gòu)建的標(biāo)準(zhǔn)曲線(標(biāo)準(zhǔn)曲線由系統(tǒng)軟件自動(dòng)分析獲得)計(jì)算出待測(cè)樣品目的基因的拷貝數(shù)。
表1基因引物序列
Table 1The primer sequences of the target genes

目的基因PCR產(chǎn)物/bp引物序列PCR條件GHR170F:5'-TGGAAATTTGTATA-ACCTCACTGCT-3'R:5'-TGGCAAGCTTAACA-CAGTATGGT-3'55℃,30sIGF-I182F:5'-CTGGTTGATGCTCT-TCAGTTCGTAT-3'R:5'-GCAGACTTAGGTG-GCTTTATTGGA-3'60℃,20s
1.7統(tǒng)計(jì)分析
運(yùn)用SPSS 20.0軟件中One-way ANOVA,t 檢驗(yàn),Univarinate,Bivariate Correlation進(jìn)行差異顯著性檢驗(yàn)和相關(guān)性分析。所有數(shù)據(jù)以Mean ± SE表示,P<0.05,表示差異顯著;P<0.01,表示差異極顯著。
2 結(jié)果與分析
2.1鴨胚胸肌成肌細(xì)胞GHR,IGF-1表達(dá)
鴨胚胸肌成肌細(xì)胞GHR表達(dá)情況如圖1-A所示,GHR mRNA表達(dá)在2個(gè)品種的變化規(guī)律基本一致,都是13胚齡表達(dá)較高,15胚齡時(shí)表達(dá)下降,17胚齡時(shí)表達(dá)上升到最高,顯著高于其他胚齡(P<0.05),隨后表達(dá)開(kāi)始下降,至23胚齡時(shí),表達(dá)量又有所上升。不同的是,高郵鴨19,21胚齡時(shí)的基因表達(dá)量與23胚齡無(wú)顯著差異,金定鴨的則是顯著低于23胚齡(P<0.05)。品種間比較,17胚齡時(shí),金定鴨基因表達(dá)量極顯著高于高郵鴨(P<0.01),而19~21胚齡時(shí),則是高郵鴨的顯著高于金定鴨(P=0.009,P=0.002)。胸肌成肌細(xì)胞GHR mRNA 表達(dá)存在極顯著的日齡(P<0.01)、日齡與品種的交互效應(yīng)(P=0.002),品種效應(yīng)不顯著(P=0.689)。
鴨胚胸肌成肌細(xì)胞IGF-1的表達(dá)如圖1-B所示,2個(gè)品種的IGF-1 mRNA變化趨勢(shì)基本一致,都是在13,15胚齡時(shí)表達(dá)較高,17胚齡的表達(dá)量顯著高于其他胚齡(P<0.05),19胚齡時(shí),表達(dá)顯著下調(diào),之后至21~23胚齡時(shí),又逐漸回升;但高郵鴨13胚齡時(shí)的表達(dá)量與17胚齡無(wú)顯著差異(P=0.769),金定鴨的則是13胚齡表達(dá)顯著低于17胚齡(P<0.05)。品種間比較,13,19,21胚齡時(shí),高郵鴨IGF-1基因表達(dá)顯著高于金定鴨(P<0.01),與之相反,17胚齡時(shí)金定鴨的顯著高于高郵鴨(P<0.01)。胸肌成肌細(xì)胞IGF-1 mRNA 表達(dá)存在極顯著的日齡(P<0.01)、日齡與品種的交互效應(yīng)(P<0.01),品種效應(yīng)不顯著(P=0.662)。
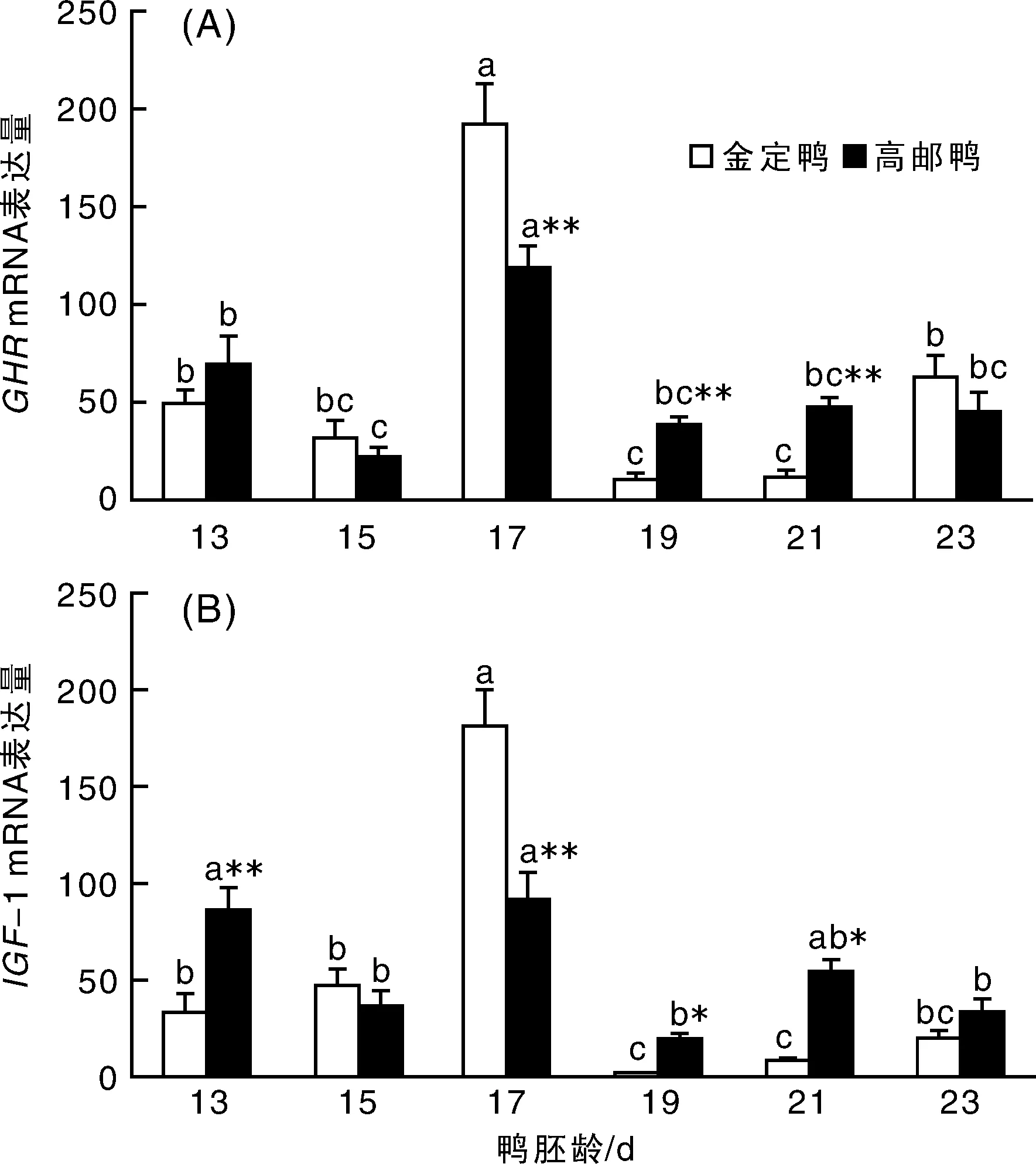
A:GHR表達(dá)量變化;B:IGF-1表達(dá)量變化。*表示同胚齡品種間差異顯著(P<0.05);**表示同胚齡的種間差異極顯著(P<0.01)。柱形圖上字母不同者表示同一品種不同胚齡之間差異顯著(P<0.05)。圖2同。圖1 鴨胚胸肌成肌細(xì)胞GHR,IGF-1 mRNA表達(dá)量變化Fig.1 The profiles of GHR and IGF-I mRNA expression in breast myoblast of different duck breeds
2.2鴨胚腿肌成肌細(xì)胞GHR,IGF-1表達(dá)
鴨胚腿肌成肌細(xì)胞GHR表達(dá)情況如圖2-A所示,GHR mRNA表達(dá)在2個(gè)品種的變化規(guī)律基本一致,17胚齡是表達(dá)的高峰期,顯著高于其他胚齡(P<0.05);其他胚齡的表達(dá)趨于平緩,相互之間無(wú)顯著差異(P>0.05)。品種間比較,17胚齡時(shí),金定鴨基因表達(dá)量顯著高于高郵鴨(P<0.01)。GHR mRNA表達(dá)存在顯著的日齡(P<0.01)、品種效應(yīng)(P<0.05)及日齡與品種的交互效應(yīng)(P<0.01)。
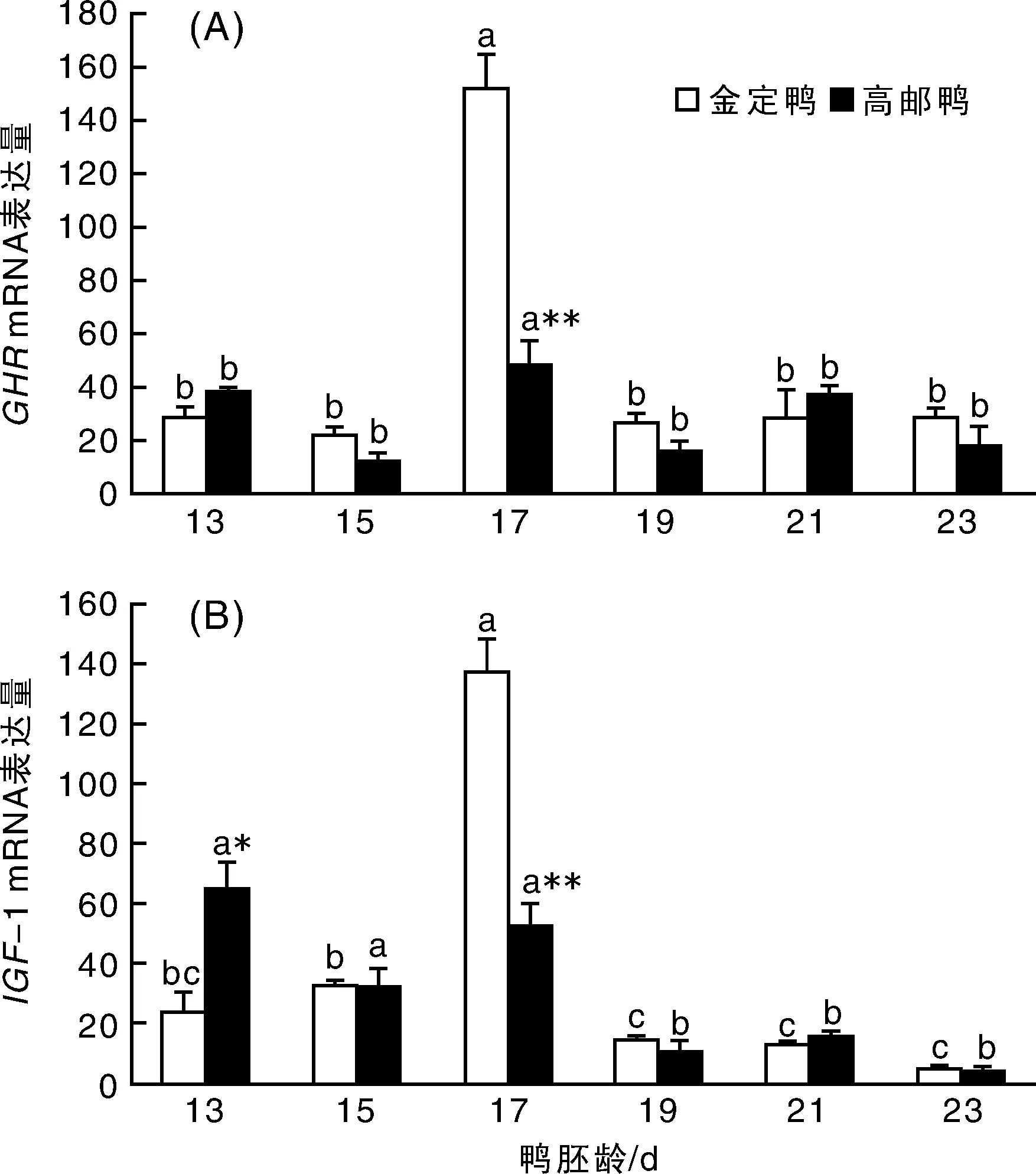
圖2 鴨胚腿肌成肌細(xì)胞GHR,IGF-1 mRNA表達(dá)量變化Fig.2 The profiles of GHR and IGF-I mRNA expression in leg myoblast of different duck breeds
鴨胚腿肌成肌細(xì)胞IGF-1的表達(dá)如圖2-B所示,2個(gè)品種的IGF-1 mRNA表達(dá)變化規(guī)律基本一致,都是13,15胚齡時(shí)表達(dá)較高,至17胚齡時(shí),上升至最高(P<0.05),19胚齡時(shí)則顯著下調(diào),并持續(xù)至23胚齡;但高郵鴨13胚齡的基因表達(dá)量與17胚齡無(wú)顯著差異(P=0.506)。品種間比較,13胚齡時(shí),高郵鴨的表達(dá)量高于金定鴨的2倍之多(P<0.05),17胚齡則相反(P<0.01)。IGF-1 mRNA 表達(dá)存在極顯著的日齡效應(yīng)(P<0.01)、日齡與品種的交互效應(yīng)(P<0.01),品種效應(yīng)不顯著(P=0.356)。
2.3鴨不同品種成肌細(xì)胞GHR與IGF-1表達(dá)的相關(guān)性
表2表明,2個(gè)品種的胸肌成肌細(xì)胞GHR與IGF-1 mRNA的表達(dá)呈極顯著的正線性相關(guān)(r=0.899,P<0.01;r=0.862,P<0.01);金定鴨腿肌成肌細(xì)胞GHR與IGF-1 mRNA表達(dá)的相關(guān)性(r=0.951,P<0.01)略高于高郵鴨(r=0.784,P<0.01)。
表2成肌細(xì)胞GHR,IGF-1 mRNA表達(dá)量的相關(guān)性分析
Table 2Correlation analysis between GHR and IGF-1 mRNA expression in myoblast

組織品種相關(guān)系數(shù)P值胸肌金定鴨0.899<0.01高郵鴨0.862<0.01腿肌金定鴨0.951<0.01高郵鴨0.784<0.01
3 討論
GH要與骨骼肌上GHR結(jié)合,才能發(fā)揮生物學(xué)作用。因此,動(dòng)物的生長(zhǎng)速度及各器官發(fā)育的優(yōu)先順序,很大程度上要受骨骼肌上的GHR及與之相關(guān)的基因表達(dá)的時(shí)空特異性影響。為了揭示成肌細(xì)胞GHR和IGF-1 mRNA的變化規(guī)律,選擇生長(zhǎng)發(fā)育存在明顯表型差異的金定鴨和高郵鴨為試驗(yàn)動(dòng)物,檢測(cè)了鴨胚骨骼肌成肌細(xì)胞GHR和IGF-1 mRNA的表達(dá),并進(jìn)行了品種間比較。結(jié)果發(fā)現(xiàn),成肌細(xì)胞GHR和IGF-1 mRNA各自的發(fā)育性表達(dá)變化特征在品種間基本一致,與之前肌肉中的研究結(jié)果相似[11-12]。17胚齡是GHR和IGF-1 mRNA共同的表達(dá)高峰期,基因的表達(dá)升高[7,13],會(huì)促進(jìn)成肌細(xì)胞向分化的方向轉(zhuǎn)變[7,13],這個(gè)時(shí)期可能是成肌細(xì)胞分化的高峰期,也是肌纖維數(shù)量形成的高峰期。17胚齡之后,高郵鴨胸肌成肌細(xì)胞GHR mRNA的表達(dá)趨于平穩(wěn),且在19,21胚齡時(shí),顯著高于金定鴨;IGF-1的表達(dá)與之類似,推測(cè)在17胚齡之后,高郵鴨胸肌成肌細(xì)胞增殖與分化活性高于金定鴨,最終導(dǎo)致在25胚齡時(shí),高郵鴨的胸肌質(zhì)量顯著高于金定鴨[14]。腿肌成肌細(xì)胞GHR,IGF-1 mRNA表達(dá)則在各胚齡間無(wú)顯著差異,與胚胎發(fā)育后期,腿肌重[14]、肌纖維直徑、面積等[15]在品種間無(wú)顯著差異相對(duì)應(yīng)。
Mavalli等[5]在小鼠上研究認(rèn)為,GH主要是通過(guò)與靶器官上的GHR結(jié)合,介導(dǎo)成肌細(xì)胞產(chǎn)生IGF-1,促進(jìn)成肌細(xì)胞分化為肌管。也有報(bào)道指出,GH促進(jìn)成肌細(xì)胞的分化,不是通過(guò)介導(dǎo)IGF-1來(lái)實(shí)現(xiàn)的[7]。本試驗(yàn)結(jié)果顯示,胸肌和腿肌成肌細(xì)胞GHR表達(dá)與各自的IGF-1表達(dá)規(guī)律基本一致,呈正相關(guān),提示GHR的表達(dá)可能發(fā)揮著正調(diào)控作用,通過(guò)調(diào)節(jié)IGF-1的表達(dá)來(lái)調(diào)節(jié)成肌細(xì)胞向增殖或分化的方向轉(zhuǎn)變。GHR表達(dá)量低時(shí),IGF-1的表達(dá)量維持在低水平,大量成肌細(xì)胞更多地進(jìn)入到G2/M期,使該時(shí)期所占時(shí)相增加,向增殖方向轉(zhuǎn)變;GHR表達(dá)升高,介導(dǎo)IGF-1表達(dá)升高,促進(jìn)成肌細(xì)胞向分化轉(zhuǎn)變,基因表達(dá)量的變化,對(duì)應(yīng)于成肌細(xì)胞增殖與分化轉(zhuǎn)變[15]。因此,17胚齡之后,隨著基因的表達(dá)顯著下調(diào),成肌細(xì)胞更多地向增殖方向轉(zhuǎn)變,相應(yīng)的肌纖維密度下降[16]。
13胚齡時(shí),胸肌成肌細(xì)胞GHR表達(dá)在品種間無(wú)顯著差異,但I(xiàn)GF-1表達(dá)是高郵鴨顯著高于金定鴨,且與17胚齡時(shí)的高郵鴨IGF-1表達(dá)無(wú)顯著差異。同樣的情況也出現(xiàn)在腿肌成肌細(xì)胞IGF-1的表達(dá)上,說(shuō)明高郵鴨骨骼肌成肌細(xì)胞比金定鴨多了一個(gè)13胚齡IGF-1的表達(dá)高峰期。由于IGF-1可以促進(jìn)成肌細(xì)胞的分化,可能高郵鴨比金定鴨多了一個(gè)成肌細(xì)胞分化的高峰期。盡管高郵鴨胸肌和腿肌成肌細(xì)胞都存在13胚齡的表達(dá)高峰期,但13胚齡時(shí)僅有胸肌質(zhì)量表現(xiàn)出品種差異[14],這可能與胸肌和腿肌成肌細(xì)胞表面分布的IGF-1受體數(shù)量不同有關(guān)[17]。
在雞上的研究認(rèn)為,禽類胚胎期血液中IGF-1主要來(lái)源于肝外其他組織[18-19]。在鴨上,13胚齡時(shí),已檢測(cè)到肝臟中IGF-1 mRNA表達(dá),并且呈現(xiàn)極顯著的品種差異[20],與13胚齡時(shí),成肌細(xì)胞IGF-1 mRNA表達(dá)出現(xiàn)品種差異相一致。IGF-1作為一種內(nèi)分泌和旁分泌激素,鴨胚胎期骨骼肌成肌細(xì)胞IGF-1的分泌,可能受到肝源性IGF-1的影響,可能是使骨骼肌發(fā)育表現(xiàn)出品種差異的因素之一。Ge等[21]在牛成肌細(xì)胞上的研究認(rèn)為,GH對(duì)成肌細(xì)胞的影響具有物種依賴性。在鴨上,GH是否通過(guò)介導(dǎo)IGF-1調(diào)節(jié)成肌細(xì)胞的增殖與分化,是課題組下一步的研究重點(diǎn)。
本研究通過(guò)檢測(cè)鴨胚成肌細(xì)胞GHR和IGF-1 mRNA表達(dá),初步證實(shí)二者表達(dá)品種效應(yīng)不顯著,在胸肌和腿肌成肌細(xì)胞存在不同的表達(dá)模式;GHR可能通過(guò)正調(diào)控IGF-1的表達(dá)來(lái)調(diào)節(jié)成肌細(xì)胞向增殖或分化的方向轉(zhuǎn)變。
[1]LUPU F,TERWILLIGER J D,LEE K,et al.Roles of growth hormone and insulin-like growth factor 1 in mouse postnatal growth [J].Developmental Biology,2001,229(1):141-162.
[2]DU M,YAN X,TONG J F,et al.Maternal obesity,inflammation,and fetal skeletal muscle development [J].Biology of Reproduction,2010,82(1):4-12.
[3]PICARD B,LEFAUCHEUR L,BERRI C E C,et al.Muscle fibre ontogenesis in farm animal species [J].Reproduction Nutrition Development,2002,42(5):415-432.
[4]SADOWSKI C L,WHEELER T T,WANG L H,et al.GH regulation of IGF-I and suppressor of cytokine signaling gene expression in C2C12 skeletal muscle cells [J].Endocrinology,2001,142(9):3890-3900.
[5]MAVALLI M D,DIGIROLAMO D J,F(xiàn)AN Y,et al.Distinct growth hormone receptor signaling modes regulate skeletal muscle development and insulin sensitivity in mice [J].Journal of Clinical Investigation,2010,120(11):4007-4020.
[6]FROST R A,NYSTROM G J,LANG C H.Regulation of IGF-I mRNA and signal transducers and activators of transcription-3 and-5 (Stat-3 and-5) by GH in C2C12 myoblasts [J].Endocrinology,2002,143(2):492-503.
[7]SOTIROPOULOS A,OHANNA M,KEDZIA C,et al.Growth hormone promotes skeletal muscle cell fusion independent of insulin-like growth factor 1 up-regulation [J].Proceedings of the National Academy of Sciences of the United States of America,2006,103(19):7315-7320.
[8]SATO K,AOKI M,KONDO R,et al.Administration of insulin to newly hatched chicks improves growth performance via impairment of MyoD gene expression and enhancement of cell proliferation in chicken myoblasts [J].General and Comparative Endocrinology,2012,175(3):457-463.
[9]SCHMID C,STEINER T,F(xiàn)ROESCH E R.Preferential enhancement of myoblast differentiation by insulin-like growth factors (IGF Ⅰ and IGF Ⅱ) in primary cultures of chicken embryonic cells [J].FEBS Letters,1983,161(1):117-121.
[10]SHAN Y J,SHU J T,SONG C,et al.Culture and Identification of Myoblasts Isolated from Duck Embryos [J].Agricultural Science &Technology,2014,15(8):1281-1284.
[11]徐文娟,單艷菊,束婧婷,等.鴨胚骨骼肌GHR基因表達(dá)與生長(zhǎng)發(fā)育的關(guān)系[J].江蘇農(nóng)業(yè)學(xué)報(bào),2013,29(1):126-130.
[12]單艷菊,顧華兵,宋衛(wèi)濤,等.不同鴨種胚胎期和岀雛早期肌肉組織中IGF-Ⅰ基因mRNA的差異表達(dá)分析[J].西北農(nóng)林科技大學(xué)學(xué)報(bào)(自然科學(xué)版),2013(3):1-6.
[13]TOLLEFSEN S E,LAJARA R,MCCUSKER R H,et al.Insulin-like growth factors (IGF) in muscle development.Expression of IGF-I,the IGF-I receptor,and an IGF binding protein during myoblast differentiation [J].Journal of Biological Chemistry,1989,264(23):13810-13817.
[14]HU Y,LIU H,SHAN Y J,et al.The relative expression levels of insulin-like growth factor 1 and myostatin mRNA in the asynchronous development of skeletal muscle in ducks during early development [J].Gene,2015,567(2):235-243.
[15]岑石強(qiáng),張峻梅,黃富國(guó),等.IGF-1 促原代人胚骨骼肌成肌細(xì)胞體外增殖與分化的研究[J].中國(guó)修復(fù)重建外科雜志,2008,22(1):84-87.
[16]單艷菊,束婧婷,胡艷,等.不同鴨種肌纖維早期發(fā)育特性及其與骨骼肌生長(zhǎng)發(fā)育的相關(guān)性研究[J].農(nóng)業(yè)生物技術(shù)學(xué)報(bào),2014,22(2):225-231.
[17]OUDIN A,CHEVALIER B,SIMON J,et al.Muscle insulin-like growth factor-I (IGF-I) receptors in chickens with high or low body weight:effects of age and muscle fibre type [J].Growth Hormone &IGF Research,1998,8(3):243-250.
[18]LU F Z,WANG X X,PAN Q X,et al.Expression of genes involved in the somatotropic,thyrotropic,and corticotropic axes during development of Langshan and Arbor Acres chickens [J].Poultry Science,2008,87(10):2087-2097.
[19]PABLO F D,ROBICS H L,CALDES T,et al.Insulin-like growth factor-I and insulin as growth and differentiation factors in chicken embryogenesis [J].Poultry Science,1991,70(8):1790-1796.
[20]胡艷,宋遲,宋衛(wèi)濤,等.鴨發(fā)育早期肝臟IGF-1 mRNA的表達(dá)特異性及其與體質(zhì)量的相關(guān)性分析[J].南京農(nóng)業(yè)大學(xué)學(xué)報(bào),2013,36(6):95-99.
[21]GE X,YU J,JIANG H.Growth hormone stimulates protein synthesis in bovine skeletal muscle cells without altering insulin-like growth factor-Ⅰ mRNA expression [J].Journal of Animal Science,2012,90(4):1126-1133.
(責(zé)任編輯盧福莊)
The expression analysis of GHR and IGF-1 mRNA in embryonic skeletal muscle myoblasts of different duck breeds
JI Gai-ge,TAO Zhi-yun,ZHU Chun-hong,SHU Jing-ting,LIU Hong-xiang,XU Wen-juan,HU Yan,LI Hui-fang*
(Jiangsu Provincial Key Laboratory of Poultry Heredity &Breeding,Jiangsu Institute of Poultry Science,Yangzhou 225003,China)
To investigate the expression patterns of growth hormone receptor (GHR) and insulin-like growth factor-1 (IGF-1) in duck skeletal muscle myoblasts,fluorescent real-time quantitative PCR was used to detect the distribution profile of GHR and IGF-I mRNA in skeletal muscle myoblasts of Jinding and Gaoyou ducks at the embryonic day 13,15,17,19,21 and 23.The results showed that the expressions of IGF-1 of Gaoyou ducks in breast and leg muscle myoblasts were significantly higher than that of Jinding ducks at embryonic day 13 (P<0.05);the embryonic day 17 was the common expression peak of GHR and IGF-1 both in Gaoyou ducks and Jinding ducks;and then,significantly decreased.But at the embryonic day 19 and 21,the expressions of GHR and IGF-1 in Gaoyou ducks breast muscle myoblasts were significantly higher than those of Jinding ducks (P<0.05),while there was no significant difference between varieties in the leg muscle myoblasts (P>0.05).The expression of GHR had a significantly linear positive correlation with the expression of IGF-1 in breast and leg muscle myoblasts(P<0.01).The results showed that the expression pattern of GHR was consistent with the expression of IGF-1 between breeds;the expressions of GHR and IGF-1 mRNA in the skeletal muscle myoblasts of duck embryo showed tissue specificity,and there might be a positive regulation mechanism between them.
duck;embryo;myoblasts;GHR;IGF-1
浙江農(nóng)業(yè)學(xué)報(bào)Acta Agriculturae Zhejiangensis,2016,28(3):406-411http://www.zjnyxb.cn
姬改革,陶志云,朱春紅,等.不同品種鴨胚胎期骨骼肌成肌細(xì)胞GHR和IGF-1 mRNA表達(dá)差異分析[J].浙江農(nóng)業(yè)學(xué)報(bào),2016,28(3):406-411.
10.3969/j.issn.1004-1524.2016.03.08
2015-08-28
國(guó)家自然科學(xué)基金資助項(xiàng)目(31172194);江蘇省科技支撐計(jì)劃項(xiàng)目(BE2014362);揚(yáng)州市農(nóng)業(yè)前瞻性研究資助項(xiàng)目(YZ2014142)
姬改革(1985—),女,河南洛陽(yáng)人,碩士,助理研究員,從事家禽遺傳育種與資源保護(hù)研究。E-mail:jigaige@126.com
,李慧芳,E-mail∶lhfxf_002@aliyun.com
S834+.8
A
1004-1524(2016)03-0406-06
猜你喜歡
英語(yǔ)世界(2023年10期)2023-11-17 09:19:16
汽車(chē)實(shí)用技術(shù)(2022年10期)2022-06-09 11:16:58
音樂(lè)探索(2022年2期)2022-05-30 21:01:37
收藏界(2019年3期)2019-10-10 03:16:40
小天使·一年級(jí)語(yǔ)數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國(guó)特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
中國(guó)非營(yíng)利評(píng)論(2017年1期)2017-11-09 03:09:10
海外華文教育(2017年8期)2017-11-07 04:42:02
現(xiàn)代語(yǔ)文(2016年21期)2016-05-25 13:13:50
