冷藏條件下帶蓬鮮蓮優勢腐敗菌鑒定及其消長規律研究
2016-09-28 01:36:24王建輝王發祥李向紅劉永樂
食品與機械 2016年8期
關鍵詞:優勢
王建輝 王 秀 王發祥 李向紅 俞 健 靳 娜 劉永樂
(長沙理工大學湖南省水生資源食品加工工程技術研究中心,湖南 長沙 410114)
?
冷藏條件下帶蓬鮮蓮優勢腐敗菌鑒定及其消長規律研究
王建輝 王秀 王發祥 李向紅 俞健 靳娜 劉永樂
(長沙理工大學湖南省水生資源食品加工工程技術研究中心,湖南 長沙410114)
通過稀釋平板法分離4 ℃條件貯藏18 d的帶蓬鮮蓮腐敗菌,提取篩選菌株DNA后進行PCR擴增,采用細菌16S rDNA菌種鑒定法對優勢腐敗菌進行鑒定,并分析其4 ℃冷藏過程中的消長規律。結果表明,帶蓬鮮蓮4 ℃冷藏過程中的優勢腐敗菌主要為分散泛菌(Pantoeadispersa)、表皮葡萄球菌(Staphylococcusepidermidis)、巴氏葡萄球菌(Staphylococcuspasteuri)、成團泛菌(Pantoeaagglomerans)。且4 ℃貯藏期間,分散泛菌、表皮葡萄球菌、巴氏葡萄球菌的變化趨勢大體相同,適應期后呈增長趨勢,成團泛菌變化規律不明顯。
鮮蓮;冷藏;優勢腐敗菌;分離鑒定;消長規律
鮮蓮營養豐富,味道鮮美,口感甜脆[1],具有益心、補腎、健脾、止瀉和安神等多重功效[2-4],但常溫下易軟化皺縮,貨架期3~6 d,采后低溫4 ℃帶蓬貯藏也僅能維持15 d[5]。鮮蓮在采摘、運輸、加工及貯藏過程中極易腐敗變質,喪失其原有的鮮食風味,直接影響其感官品質及食用價值,極大限制了鮮食蓮子的銷售和消費者的需求。研究[6]表明,微生物活動是引起果蔬腐敗變質的主要原因之一,果蔬常見腐敗菌有:沙門氏菌(Salmonella)、假單胞菌(Pseudomonasspp)、李斯特菌(Listeria)、黃單胞菌(Xanthomonas)等[7],對蓮子危害最大的微生物主要是曲霉屬和青霉屬霉菌[8]。目前國內外鮮蓮研究處于起步階段,有關鮮蓮優勢腐敗菌分離鑒定研究尚無報道。隨著分子生物學的快速發展,細菌的分類鑒定從傳統的表型、生理生化分類深入到各種基因型分類水平,如質粒圖譜、限制性片段長度多態性分析、PCR 指紋圖、16S rRNA 序列分析等技術[9-10]。16S rDNA 序列分析技術在一定程度上克服了傳統表型、生理生化指標鑒定微生物的局限性,能簡便、快速準確地鑒定微生物[11-12]。
本研究以帶蓬鮮蓮為研究對象,通過16S rDNA 序列分析技術初步鑒定低溫貯藏下帶蓬鮮蓮的主要優勢腐敗菌,并分析其優勢菌的消長變化規律,對進一步監控和靶向抑制鮮蓮品質變化以及鮮蓮低溫貯藏新技術的開發,提高其貯藏品質,延長貨架期均具有很好的理論價值。
1 材料與方法
1.1材料與試劑
新鮮蓮蓬:由湖南省湘潭縣湘蓮種植基地提供;
PCA培養基、營養瓊脂培養基:廣東環凱微生物科技有限公司;
Folin酚試劑、NaCl、異戊醇:分析純,國藥集團化學有限公司;
瓊脂糖:電泳級,生工生物工程上海股份有限公司;
Tris-乙酸-EDTA:電泳級,北京索萊寶生物科技有限公司;
溴化乙錠(EB)、PCR試劑等:電泳級,美國sigma公司。
1.2儀器與設備
電子分析天平:AUY120型,日本島津公司;
立式壓力蒸汽滅菌器:DY01-13-44-00型,上海東亞力容器制造有限公司;
超聲波清洗器:KQ-500B型,昆山市超聲儀器有限責任公司;
搖床:ZHWY-100C型,上海智城公司;
生化培養箱:DZX180型,上海艾測電子科技有限公司;
顯微鏡:BH-2型,日本Olympus公司;
PCR儀:T100型,美國Thermo公司。
1.3方法
1.3.1鮮蓮原料處理新鮮蓮蓬在低溫條件下快速運回實驗室。選取成熟度一致、大小均勻、無病蟲害和機械損傷的帶蓬鮮蓮樣品于RH(相對濕度)65%~70%,溫度4 ℃恒溫條件下貯藏。
1.3.2優勢菌落的確定根據王建輝等[5]的研究,帶蓬鮮蓮貨架期為15 d,18 d時蓮蓬表面菌斑明顯增多,質地明顯變軟,感官品質明顯下降。故取貯藏18 d的帶蓬鮮蓮樣品,在超凈環境下,于無菌的組織搗碎機中搗碎,準確稱取搗碎后的蓮肉樣品10.00 g于250 mL無菌的錐形瓶中,注入90 mL無菌生理鹽水,振蕩30 min。然后取1 mL進行10倍梯度稀釋,在預試驗的基礎上,選擇3個適合的稀釋度傾注平板,平行3份,36 ℃培養48 h后進行菌落計數。并通過肉眼觀察,記錄各個稀釋度的菌落特征,并將數量最多的4~5種菌落視為優勢菌。
1.3.3優勢菌的分離純化挑選1.3.2中各優勢菌于PCA平板上進行多次劃線分離,得到單菌落后繼續傳代培養,對各代的單菌落進行革蘭氏染色,顯微鏡觀察細胞形態、大小等,直至盡可能得到純的單菌落,將純化好的菌株保存于營養瓊脂斜面試管,并于4 ℃低溫保存備用。
1.3.4優勢腐敗菌的鑒定
(1) 形態觀察:將經1.3.3純化后菌株接種于平板上,36 ℃下培養48 h,期間結合肉眼與革蘭氏染色后顯微鏡觀察各優勢菌的菌落特征、細胞形態[13-14]以及相對比例等。
(2) 細菌16S rDNA菌種鑒定:經純化的菌株采用細菌16S rDNA 菌種鑒定方法進行鑒定[15]。將純化的5 種優勢腐敗菌接種于10 mL LB 培養基中,37 ℃振蕩過夜培養,分別提取1#~5#腐敗細菌的基因組DNA,以通用引物:16S(F) 5'-AGAGTTTGATCCTGGCTCAG-3';16S(R) 5'-GGTTACCTTGTTACGACTT-3' 。
PCR擴增其16S rDNA,回收1 500 bp 長度左右的PCR 產物,送至上海生工股份有限公司進行DNA測序,將測定的序列通過Blast 程序與GenBank 數據庫中已有的16S rDNA序列進行相似性比較分析,根據16S rDNA序列相似性構建系統發育樹,判定細菌種類。
1.3.5鮮蓮貯藏過程中腐敗菌的生長規律參考Chang Shu-chen等[16]的方法,并稍做修改。將于4 ℃冷藏條件下0~24 d的鮮蓮每3 d取樣,在無菌條件下進行稀釋平板計數,分別統計各優勢腐敗菌個數,并繪制其生長規律曲線。
2 結果與分析
2.1腐敗菌的分離和優勢腐敗菌的確定
通過傾注平板、培養、觀察、計數,根據菌落形態特征將所有菌落大致分為8組,分別編號為1#~8#,將數量最多5種菌視為優勢腐敗菌,鮮蓮低溫貯藏條件下5種優勢腐敗菌菌落形態及相對比例見表1。

表1 優勢腐敗菌形態及組成
2.2優勢腐敗菌的鑒定
2.2.1優勢腐敗菌16S rDNA的擴增結果分別提取腐敗菌1#~5#號的基因組DNA,進行PCR擴增,擴增出的PCR產物進行0.8%瓊脂糖凝膠電泳。由圖1可知,1#~5#號腐敗細菌基因經PCR擴增后,擴增產物經電泳分離獲得比較清晰的條帶,可見PCR擴增產物主要位于1 500 bp左右,且獲得的目的基因經凝膠電泳可很好地純化分離。
2.2.2優勢腐敗菌16S rDNA 鑒定結果通過細菌16S rDNA可變區的差異對菌種類別進行區分。回收2.2.1中的目的基因片段,并送至上海生工股份有限公司測序,將測序獲得的1#~5#腐敗菌的16S rDNA 序列與NCBI數據庫中已有的序列進行Blast相似檢索,從Genbank核苷酸數據庫中挑選同源性較高的16S rDNA序列進行比較,利用MAGE 5軟件構建系統發育樹[17],結果見圖2。1#(L1)、2#(L2)菌株分別與分散泛菌(Pantoeadispersa)、表皮葡萄球菌(Staphylococcusepidermidis)親緣關系最近,驗證可信度達99%;3#(L3)和5#(L5)菌株為同一菌屬,且與巴氏葡萄球菌(Staphylococcuspasteuri)親緣關系最近,驗證可信度達100%;4#(L4)菌株與成團泛菌(Pantoeaagglomerans)親緣關系最近,驗證可信度達100%,鑒定結果見表2。

圖1 腐敗細菌的擴增結果
2.3鮮蓮低溫貯藏過程中優勢腐敗菌的消長規律
由圖3可知,隨著貯藏時間的延長,4種優勢腐敗菌:分散泛菌、表皮葡萄球菌、巴氏葡萄球菌的變化趨勢大體相同,呈現先降后升的趨勢,貯藏第3天時,優勢腐敗菌菌落數都大幅度降低,研究現象與王滿生等[18]對草魚低溫貯藏過程中優勢腐敗菌前期先大幅降低的結果相似,可能是鮮蓮采后貯藏溫度突然降低至冷藏4 ℃,導致部分細胞生長受抑或遇冷死亡引起。其中,巴氏葡萄球菌變化最為明顯,第3天后菌落數上升迅速,表皮葡萄球菌上升較分散泛菌變化略微明顯,第3天時菌落數低于分散泛菌,但第9天其菌落數超過了分散泛菌。在貯藏期間成團泛菌菌落數變化無規律,可能是此菌本身的特點,易成片聚集,菌落數變化規律不明顯。
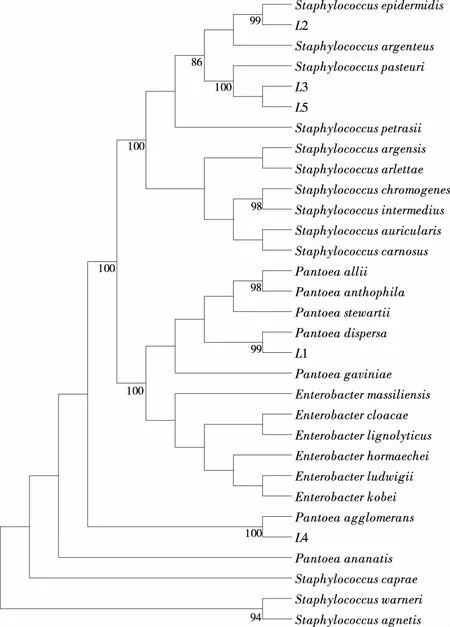
圖2 1#~5# 鮮蓮腐敗菌16S rDNA 的系統發育樹

編號Blast相似性最高菌株種群1#分散泛菌(Pantoeadispersa)2#表皮葡萄球菌(Staphylococcusepidermidis)3#和5#巴氏葡萄球菌(Staphylococcuspasteuri)4#成團泛菌(Pantoeaagglomerans)

因3# 和5#屬于同一菌屬,故合并統計
3 結論
本研究分離鑒定得到了鮮蓮4 ℃貯藏過程中的4種優勢腐敗菌,即分散泛菌(Pantoeadispersa)、表皮葡萄球菌(Staphylococcusepidermidis)、巴氏葡萄球菌(Staphylococcuspasteuri)和成團泛菌(Pantoeaagglomerans);明確了其在貯藏期間的消長規律,隨著貯藏時間的延長,分散泛菌、表皮葡萄球菌、巴氏葡萄球菌的生長趨勢大體相同,呈現先降后升的趨勢,巴氏葡萄球菌變化最為突出,第3天后菌落數上升迅速,表皮葡萄球菌上升較分散泛菌變化略微明顯,貯藏期間成團泛菌菌落數變化無規律。當前研究分離鑒定了鮮蓮4 ℃貯藏過程中的優勢腐敗菌種類,初步掌握了其各自的消長規律,然而,對其優勢腐敗菌的致腐能力及其致腐機理尚需進一步深入探討,對鮮蓮低溫貯藏新技術的開發及其產品的研發有待進一步開展集成研究。
[1] 鄭天聞, 徐婷婷, 范亞葦, 等. 蓮子多酚氧化酶的酶學性質[J]. 食品工業科技, 2014, 35(2): 162-165.
[2] BHAT R, SRIDHAR K R. Nutritional quality evaluation of electron beam-irradiated lotus (Nelumbonucifera) seeds[J]. Food Chemistry, 2008, 107(1): 174-184.
[3] WU Jin-zhong, ZHENG Yuan-bin, CHEN Ti-qiang, et al. Evaluation of the quality of lotus seed ofNelumbonuciferaGaertn from outer space mutation[J]. Food Chemistry, 2007, 105(2): 540-547.
[4] 王亮亮, 夏延斌, 任美. 湘蓮的營養和保健功效及其在食品加工中的應用[J]. 食品與機械, 2015, 31(2): 262-266.
[5] 王建輝, 靳娜, 劉永樂, 等. 低溫處理對湘蓮采后生理變化的影響[J]. 食品科學, 2014, 35(18): 209-213.
[6] TOURNAS V H. Moulds in fresh and minimally processed vegetables and sprouts[J]. International Journal of Food Microbiology, 2005, 99(1): 71-77.
[7] 范賢賢, 田密霞, 姜愛麗, 等. 鮮切果蔬表面微生物侵染途徑及控制[J]. 保鮮與加工, 2009(2): 14-16.
[8] 易俊敖. 蓮子的貯藏技術[J]. 長江蔬菜, 1992(2): 38-39.
[9] MIGNARD S, FLANDROIS J P. 16S rRNA sequencing in routine bacterial identification: a 30-month experiment[J]. Journal of Microbiological Methods, 2006, 67(3): 574-581.
[10] 楊靜靜, 孟鎮, 鐘其頂, 等. 分子生物學技術在酵母菌多相分類鑒定中的應用[J]. 中國釀造, 2011(4): 16-19.
[11] LEI Zhang-yu, ZHANG Xiao. Application of 16S rDNA sequence analysis technique in microbial detection[J]. Agricultural Science&Technology, 2014, 15(4): 520-523.
[12] PHILIPP S, HUEMER H P, IRSCHICK E U, et al. Obstacles of multiplex real-time PCR for bacterial 16S rDNA: primer specifity and DNA decontamination of Taq polymerase[J]. Transfusion Medicine & Hemotherapy, 2010, 37(1): 21-28.
[13] KIERZKOWSKA M, MAJEWSKA A, KUTHAN R T, et al. A comparison of Api 20A vs MALDI-TOF MS for routine identification of clinically significant anaerobic bacterial strains to the species level[J]. Journal of Microbiological Methods, 2013, 92(2): 209-212.
[14] 郭全友, 楊憲時, 許鐘. 冷藏羅非魚優勢腐敗菌的鑒定及其特征[J]. 食品與機械, 2009, 25(3): 87-90.
[15] SUN Shan, FAN Xin-xing, PHARMACY D O, et al. RAPD-PCR and 16S rDNA phylogenetic analysis of alkaline protease producing bacteria isolated from soil of India: Identification and detection of genetic variability[J]. Journal of Genetic Engineering & Biotechnology, 2014, 12(1): 27-35.
[16] CHANG Shu-chen, KUND H F, CHEN H C, et al. Determination of histamine and bacterial isolation in swordfish fillets (Xiphiasgladius) implicated in a food borne poisoning[J]. Food Control, 2008, 19(1): 16-21.
[17] 韓永和, 章文賢, 莊志剛, 等. 耐鹽好氧反硝化菌A-13菌株的分離鑒定及其反硝化特性[J]. 微生物學報, 2013, 53(1): 47-58.
[18] 王滿生. 多靶柵欄技術在草魚貯藏過程中對微生物腐敗的控制研究[D]. 長沙: 長沙理工大學, 2012: 21-34.
The dominant spoilage bacteria and their growth and decline law in fresh lotus seeds storedat the cold storage
WANG Jian-huiWANGXiuWANGFa-xiangLIXiang-hongYUjianJINNaLIUYong-le
(HunanProvincialEngineeringResearchCenterforFoodProcessingofAquaticBioticResources,ChangshaUniversityofScienceandTechnology,Changsha,Hunan410114,China)
In this study, spoilage bacteria in fresh Xiang lotus seeds preserved at 4 ℃ temperature for 18 d were separated by dilution-plate method. Total genomic DNA was extracted from these strains and then subjected to PCR amplification. Dominant spoilage bacteria were identified using 16S rDNA sequencing for the identification and the law of their growth and declination was drawn. The results showed the four dominant spoilage bacteria in fresh Xiang lotus seeds preserved at the temperature of 4 ℃ werePantoeadispersa,Staphylococcusepidermidis,Staphylococcuswarneri, andPantoeaagglomerans. At the storage temperature of 4 ℃, the growing trends ofPantoeadispersa,Staphylococcusepidermidis,Staphylococcuswarneriof fresh lotus seeds were similar, which all needed an adaptive phase before their increase. The amount change ofPantoeaagglomeranswas irregular.
Fresh Lotus; Cold storage; dominant spoilage bacteria; identification; growth and decline law
國家自然科學基金青年科學基金項目(編號:31301564,31201427);湖南省自然科學基金項目(編號:2015JJ2011);湖湘青年英才支持計劃項目(編號:2015RS4051)
王建輝,男,長沙理工大學教授,博士。
劉永樂(1962-),男,長沙理工大學教授,博士。
E-mail:LYLE19@163.com
2016-06-07
10.13652/j.issn.1003-5788.2016.08.026
猜你喜歡
我愛學·笑話與口才(2025年3期)2025-02-24 00:00:00
華人時刊(2020年13期)2020-09-25 08:21:30
趣味(語文)(2020年3期)2020-07-27 01:42:46
意林·全彩Color(2019年8期)2019-09-23 02:12:36
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
中國衛生(2015年2期)2015-11-12 13:13:54
中國火炬(2014年11期)2014-07-25 10:31:58
中國體育(2004年3期)2004-11-11 08:53:02
棋藝(2001年19期)2001-11-25 19:55:34
棋藝(2001年23期)2001-01-06 19:08:36
