沙生植物倒披針葉蟲實在古爾班通古特沙漠的分布
2016-08-29 02:39:13陳正霞劉尊馳何文琴湯景光郝曉冉孫園園
生態學報 2016年13期
關鍵詞:植物
陳正霞,劉 彤,劉尊馳,何文琴,湯景光,郝曉冉,孫園園,曾 勇
石河子大學生命科學學院,石河子 832003
?
沙生植物倒披針葉蟲實在古爾班通古特沙漠的分布
陳正霞,劉彤*,劉尊馳,何文琴,湯景光,郝曉冉,孫園園,曾勇
石河子大學生命科學學院,石河子832003
倒披針葉蟲實(Corispermumlehmannianum)在古爾班通古特沙漠廣泛分布,重要值僅次于該沙漠建群種白梭梭(Haloxylonpersicum),對沙面的穩定起著非常重要的作用。采用固定樣方法對自然條件下該植物的分布特征和種群數量動態進行了研究。結果表明:1)倒披針葉蟲實在沙丘各坡位均占有較高重要值,尤其活化程度較高的中部和上部優勢度更明顯;2)生長季節的不同階段,除坡頂外,其他各坡位種子主要分3批萌發出苗,不同時期種子萌發出苗數量存在差異,3月份有大量種子萌發出苗,4月份種子萌發出苗數最多,5月份萌發出苗量逐漸減少。而坡頂3月份種子萌發出苗量最多,4月至5月均無種子萌發出苗。因此,除坡頂外,種子萌發出苗數量和種群密度在其他各坡位呈先增后減的趨勢。生長季結束時各坡位種群密度表現為:中部和上部>底部和丘間>頂部;3)成熟植株單位面積種子產量為:中部和上部>丘間和底部>頂部,且中部和上部單位面積種子數量極顯著多于丘間、坡底和坡頂(P<0.01);4)通過對0—5 cm和6—10 cm土壤種子庫密度研究發現,沙丘中部和上部種子庫密度顯著高于其他坡位。綜合以上研究表明,倒披針葉蟲實在沙面較活化的中部和上部呈優勢分布,對沙漠環境具有良好適應性。因此,該植物是一種極具開發潛力的沙生植物。
短命植物;倒披針葉蟲實;種群時空變化;土壤種子庫
一年生植物是全球荒漠植物區系中重要而獨特的組成部分[1-5],具有生活史短暫、高光合能力、繁殖力強、抗干擾能力強、種群持續力強、蓋度大、生物量高等特點[6]。在荒漠地區的植被恢復過程中發揮著重要作用,對荒漠生態系統的穩定和生物多樣性的維持也有重要影響。
古爾班通古特沙漠位于新疆北部準噶爾盆地腹地,是我國最大的固定與半固定沙漠,面積為4.88×104km2,其中97%呈固定與半固定狀態。本沙漠一年生植物占優勢,已統計到有97種,約占該沙漠植物總種數的47%[7]。而短命植物是一種特殊類型的一年生植物,在生長季節的4、5、6三個月是北疆沙漠風沙活動強盛期,此時該類植物的蓋度分別達到13.9%、40.2%和14.1%,能夠有效阻止地表風沙流動,對保持沙漠穩定、防風固沙起到重要作用,是植被穩定沙面的主要貢獻者[8]。
蟲實屬(Corispermum)是藜科(Chenopodiaceae)的一個大屬,約有60余種,均為一年生沙生植物。全都分布在北半球溫帶的干旱地區,除少數種類出現于北美外,絕大多數分布于歐亞大陸而以亞洲最多[9]。我國有20余種,沙漠地區常見有11種。新疆分布有3種,即:中亞蟲實(C.heptapotamicum)、蒙古蟲實(C.mongolicum)和倒披針葉蟲實(C.lehmannianum),前兩者分布于塔里木盆地。而倒披針葉蟲實集中分布于準噶爾盆地南緣的古爾班通古特沙漠,為一年生短命植物,株高常10—30 cm,花果期4—6月[10-11]。張榮[5]等人通過對古爾班通古特沙漠植物多樣性近5年的調查研究結果顯示,倒披針葉蟲實在古爾班通古特沙漠廣泛分布,且其重要值達到31.45,僅次于該沙漠的建群種白梭梭(Haloxylonpersicum),防風固沙的生態功能顯著。因此對該植物進行深入研究具有重要的生態學意義。
目前關于蟲實屬植物的研究國外主要集中在分類學方面[12-14],而國內學者則對蟲實屬植物的研究主要有,He Y H等[15]對不同類型沙地一年生植物長穗蟲實的繁殖分配及其與個體大小的關系進行了研究;Huang Y X等[16]研究了土壤養分、水分及種群密度對大果蟲實異速生長的影響;Wang Y H等[17]研究了機械擾動對蒙古蟲實植株機械壓力的影響;劉有軍等[18]對碟果蟲實種子萌發對策及生態適應性進行了研究;Liu Y J等[19]研究了不同儲藏溫度對碟果蟲實種子萌發和休眠特性的影響。但是對于古爾班通古特沙漠廣泛分布的倒披針葉蟲實時空分布規律的研究目前尚未見報道。
通過野外調查發現,古爾班通古特沙漠不同坡位沙面穩定性存在差異,一般丘間和坡底呈固定狀態,中部和上部較活化,坡頂呈流動狀態。因此,不同坡位植被分布也就存在差異,固定的丘間和坡底物種豐富度高,活化的中部和上部次之,流動的頂部幾乎沒有植被分布。而倒披針葉蟲實在古爾班通古特沙漠廣泛分布,那么其在沙丘不同坡位的分布特征是怎樣的?在整個生長季節,不同坡位倒披針葉蟲實的種群數量動態又是怎樣的?這是本研究要解決的關鍵問題。
為了解決上述問題,本文在對古爾班通古特沙漠51個樣地進行廣泛調查的基礎上,選取倒披針葉蟲實分布的典型樣區——莫索灣地區為研究區,定點對比研究了沙丘不同坡位該植物一個生長季種群統計學(幼苗萌發出苗,死亡和存活,種群密度,單株種子數量,單位面積植株種子總產量)、土壤種子庫特征及不同坡位物種組成,以期了解倒披針葉蟲實在沙丘上的分布特征及種群數量動態,有助于理解倒披針葉蟲實能在古爾班通古特沙漠中成為短命植物片層中優勢種的原因。
1 材料與方法
1.1研究區概況
研究區位于古爾班通古特沙漠腹地莫索灣地區,經緯度為45°06′N,86°19′E,海拔為350 m。該地區屬典型的溫帶大陸性干旱荒漠氣候。全年降雨量小于120 mm,平均積雪深度17 cm,年蒸發量為1942.1 mm。全年日照2777 h,年積溫3594 ℃,春夏季為風季,最大風速20 m/s,地下水位13 m左右。沙丘丘間和坡底沙面固定,上部和中部呈活化狀態,頂部呈流動狀態。
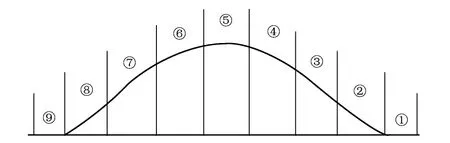
圖1 研究區調查樣地沙丘地貌分布圖Fig.1 Landforms map of study area dunes ①陰坡丘間(P1);②陰坡底部(P2);③陰坡中部(P3);④陰坡上部(P4);⑤坡頂部(P5);⑥陽坡上部(P6);⑦陽坡中部(P7);⑧陽坡底部(P8);⑨陽坡丘間(P9)
調查沙丘的主要物種組成有白梭梭(H.persicum)、梭梭(H.ammodendron)、淡枝沙拐棗(Calligonumleucocladum)、準噶爾沙蒿(Artemisiasongarica)及其他一些草本植物。倒披針葉蟲實在其間廣泛分布,調查沙丘劃分為9個坡位(圖1)。
1.2研究方法
1.2.1樣地設置
在古爾班通古特沙漠南部的莫索灣沙地中選擇一個代表性樣地,在2013年3下旬幼苗出土時期,采用樣方法,在沙丘陰坡和陽坡的丘間、底部、中部、上部和頂部分別設置3 m×3 m的固定樣方3—5個。
1.2.2種群統計學的觀測方法
從3月底到6月初,共進行了4次調查。每個觀測時間點上,記錄樣方內倒披針葉蟲實存活數、新萌數和死亡數及種群密度,同時記錄其他物種名、數量、株高及冠幅。植株成熟時,在各個坡位的每個樣方采集大(≥15 cm)、中(10—15 cm)和小(≤10 cm)3種類型的植株,每種類型至少10株帶回實驗室,進行種子產量的測定。
1.2.3土壤種子庫及種子活力分析
2013年3月13日(植物種子萌發之前)采用隨機取樣的方法取樣,在沙丘的不同坡向(陰坡和陽坡)的各個坡位(丘間、底部、中部、上部和頂部)的3個重復樣方內,采集深度0—5 cm和6—10 cm土樣,每個土層均取3個重復,采樣面積50 cm×50 cm。土樣帶回實驗室,采用不同目的土壤篩篩選倒披針葉蟲實的種子。對篩出的種子采用萌發法進行活力測定,并統計土壤中具活力種子數量。土壤種子庫密度用單位面積土壤內所含具活力種子數量來表示,將取樣面積內的種子數量換算成1 m2的數量,計算出不同坡位0—5 cm和6—10 cm土層中土壤種子庫密度(粒/m2)。
1.3數據處理方法
實驗同時收集當地降雨資料。實驗所得數據采用Excel和SPSS17.0軟件進行處理。對沙丘不同坡位倒披針葉蟲實種子庫密度、成熟植株單株種子數量和單位面積種子數量進行單因素方差分析(One-way ANOVA),并采用鄧肯法(Ducan)進行多重比較。
計算出樣方內各植物種的多度、相對多度、蓋度、相對蓋度、頻度、相對頻度、高度、相對高度以及每個種在樣方中的重要值。重要值的計算公式:重要值(IV)=(相對蓋度+相對頻度+相對多度+相對高度)/4。制圖采用origin8.5軟件(origin Inc.)。
2 結果與分析
2.1沙丘不同坡位物種組成特征
在研究區沙丘9個坡位共調查到18個物種,隸屬8科18屬(表1)。在陰坡,丘間和底部各有14個物種,占調查樣地總物種數的77.78%,其中丘間重要值較高物種有:倒披針葉蟲實(C.lehmannianum)、彎花黃耆(Astragalusflexus)、梭梭(Haloxylonammodendron)、硬萼軟紫草(Arnebiadecumbens)、狹果鶴虱(Lappulasemiglabra)、彎曲四齒芥(Tetracmerecurvata)和卷果澀薺(Malcolmiascorpioides);坡底重要值較高物種有:倒披針葉蟲實、梭梭、彎花黃耆、狹果鶴虱和彎曲四齒芥。中部和上部物種數分別為9種和7種,占總物種數的50%和38.89%,重要值較高物種均為:倒披針葉蟲實、白梭梭(Haloxylonpersicum)。在陽坡,丘間和底部物種數分別為12種和11種,分別占總物種數的66.67%和61.11%,其中丘間重要值較高的物種有:倒披針葉蟲實、梭梭、彎花黃耆、狹果鶴虱和卷果澀薺;坡底重要值較高的物種有:倒披針葉蟲實、梭梭、彎花黃耆、狹果鶴虱、沙蓬(Agriophyllumsquarrosum)、早熟豬毛菜(Salsolapraecox)和彎曲四齒芥。中部和上部均有8個物種,占總物種數的44.44%,中部和上部重要值較高的物種均為:倒披針葉蟲實和白梭梭。而頂部僅分布有3個物種,占總物種數的16.67%,重要值高的物種為倒披針葉蟲實。
進一步分析發現,倒披針葉蟲實在沙丘各個坡位均占有較高的重要值,但不同坡位重要值大小存在差異:在陰坡,丘間為45.58,底部為56.01,中部為63.79,上部為63.60;頂部為69.82;在陽坡,丘間為42.13,底部為44.14,中部68.21,上部為69.82。由此可以看出,在沙丘中上部倒披針葉蟲實的重要值明顯大于丘間和坡底,說明在沙丘中上部倒披針葉蟲實分布的重要性要高于在丘間和坡底的重要性。

表1 沙丘不同坡位植被組成及其重要值Table 1 Species composition and their Important Value (IV) on different slope positions
2.2不同坡位倒披針葉蟲實種子萌發出苗及死亡情況
圖2是莫索灣2013年3月到6月的逐日降雨量圖(數據來自石河子氣象局網站),這一時間是當地一年生植物的主要生長季節,從圖2中看出4月和5月有大量的降雨,總體的降雨特征具有很強的隨機性和不確定性。而4月和5月份的溫度也是該地區大多數一年生植物種子萌發的適宜溫度。

圖2 2013年3—6月莫索灣逐日降雨量Fig.2 Daily precipitation in mosuowan from March to June, 2013
由表2可以看出,流動狀態的坡頂除外,從丘間(沙面固定樣地)到坡上部(沙面活化樣地),倒披針葉蟲實種子萌發數量均逐漸增多,種子萌發數量表現出隨沙面的活化程度增加逐漸增加的變化趨勢。從3月份到6月份,除坡頂部外,其他各個坡位倒披針葉蟲實出苗數量呈先增加后減少的趨勢。其中,4月份出苗數量在除坡頂部外的其他8個坡位均達到最大值。當月各坡位的幼苗數量,在陰坡為:丘間(251±43)株,底部(293±78)株,中部(502±67)株,上部(805±53)株;坡頂部為:(0)株;在陽坡為:丘間(167±48)株,底部(278±23)株,中部(472±69)株,上部(509±62)株。5月份各坡位有少量種子萌發,6月份各坡位無種子萌發。由表2也可以看出,倒披針葉蟲實主要分3批萌發出苗,大量出苗集中在4月份,之后出苗量逐漸減少。

表2 9個坡位調查樣方內倒披針葉蟲實種子萌發出苗及死亡情況(平均值±標準誤)Table 2 Germination and death ofCorispermumLehmannianumon 9 slope positionsin in different periods (Mean Value±SE)
從4月份到6月份,沙丘不同坡位倒披針葉蟲實植株死亡數量呈先增加后減少的趨勢。其中,5月份各坡位植株死亡數量均達到最大值,該月9個坡位植株死亡數量,陰坡為:丘間(213±48)株,底部(249±49)株,中部(95±20)株,上部(82±17)株;坡頂部為:(53±25)株;陽坡為:丘間(87±13)株,底部(143±46)株,中部(103±27)株,上部(91±19)株(表2)。
從3月份到6月份,沙丘各坡位樣方內倒披針葉蟲實存活植株數量變化情況為,陰坡丘間、陰坡底部、陽坡丘間和陽坡底部樣方內存活植株數量4月份達到最大值,分別為:(455±24)株、(575±125)株、(236±25)株、(469±32)株,5、6月份植株數量逐漸減少;而陰坡中部、陰坡上部、陽坡中部和陽坡上部樣方內存活植株數量5月份達到最大值,分別為:(985±106)株、(1335±124)株、(625±30)株、(736±81)株,6月份幾乎保持不變;坡頂部在3月份樣方內存活植株數量最多為(222±65)株,4月份到6月份樣方內植株數量呈明顯的下降趨勢(表2)。
綜上所述,整個生長季,從空間尺度上來看,除沙面呈流動狀態的坡頂外,丘間到沙丘上部,隨著沙面的活化,倒披針葉蟲實植株數量增加,沙面呈活化狀態的中部和上部植株數量明顯多于丘間和坡底;從時間尺度上來看,沙丘各坡位倒披針葉蟲實種子萌發出苗規律為:4—5月份各坡位均具有很高的幼苗死亡率,同時在這個時間段又有大量的新植株補充進來,高的出生率來補償死亡數量,當到達6月份后,死亡數量逐漸減少,種群數量趨于穩定。
2.3倒披針葉蟲實幼苗種群密度變化
同一時期不同坡位,倒披針葉蟲實幼苗種群密度存在差異(圖3)。除3月份外,各時期種群密度大小總體表現為:中部和上部>底部和丘間>頂部。5月份沙丘各坡位種群密度均達到最大值,其中中上部密度最大值達到(148±14)株/m2,而下部(底部和丘間)的最大密度僅為(64±14)株/m2,沙丘中上部最大密度是下部最大密度的2.32倍。在整個生長季結束時,各個坡位種群密度分別為:陰坡丘間(33±7)株/m2,陰坡底部(43±12)株/m2,陰坡中部(108±12)株/m2,陰坡上部(147±14)株/m2,坡頂部(12±7)株/m2,陽坡丘間(17±4)株/m2,陽坡底部(38±4)株/m2,陽坡中部(67±3)株/m2,陽坡上部(80±9)株/m2。

圖3 不同坡位各時期倒披針葉蟲實種群密度曲線(平均值±標準誤)Fig.3 Population density of Corispermum Lehmannianum of different slope positions in different periods (Mean Value±SE)
同一坡位不同時期,除坡頂倒披針葉蟲實種群密度呈直線下降趨勢外,其他各坡位整體呈現先增加后下降的趨勢:下部(丘間和坡底),4月份種群密度達到最大值,5月份下降,6月份趨于穩定;中部和上部,5月份種群密度達到最大值,6月份稍微下降并趨于穩定(圖3)。
2.4不同坡位倒披針葉蟲實單株及單位面積植株種子數量
沙丘不同坡位,倒披針葉蟲實在成熟時期單株種子數量存在差異(圖4)。結果發現:在陽坡,中部和上部>底部和丘間>頂部;而在陰坡,丘間和底部>中部和上部>頂部。
進一步分析發現,在陽坡,中部單株種子數量最多,與其它坡位存在極顯著性差異(P<0.01),上部次之,與底部差異不顯著(P>0.05),而與丘間和頂部差異極顯著(P<0.01)。在陰坡,丘間單株種子數量最大,與其它坡位存在極顯著性差異(P<0.01),上部最少,與中部差異性不顯著(P>0.05)。
對不同坡位單位面積植株種子數量進行對比分析發現,陽坡和陰坡均為:中部和上部>丘間和底部>頂部(圖5)。沙丘中部和上部單位面積種子數量與丘間、坡底和坡頂存在極顯著性差異(P<0.01),而中部與上部差異性不顯著(P>0.05)。

圖4 不同坡位倒披針葉蟲實單株種子數量 Fig.4 Seed numbers of individual plants of Corispermum Lehmannianum on different slope positions

圖5 不同坡位倒披針葉蟲實單位面積種子數量 Fig.5 Seed numbers of unit area of Corispermum Lehmannianum on different slope positions
2.5不同坡位倒披針葉蟲實種子庫數量差異
沙丘不同坡位和不同土層,倒披針葉蟲實土壤種子庫密度均存在差異(表3)。在9個不同坡位上,0—5 cm和6—10 cm土層中,兩個坡向土壤種子庫密度均為,中上部>丘間和坡底>坡頂,且中部和上部極顯著高于其他坡位。

表3 沙丘不同部位0—5 cm和6—10 cm土層中倒披針葉蟲實種子庫密度差異(平均±標準誤)Table 3 Seed bank density differences ofCorispermumLehmannianumon different positions of the sand dunes including two soil layers 0—5 cm and 6—10 cm
在兩個不同土層中,中上部6—10 cm土層土壤種子庫密度高于0—5 cm土層,而丘間和坡底0—5 cm土層土壤種子庫密度高于6—10 cm土層。
3 討論
通過對研究區沙丘9個坡位物種組成特征進行分析發現,倒披針葉蟲實在沙丘各坡位均占有較高的重要值,為該區域的優勢物種,尤其沙丘中上部優勢度更明顯。因此,對保持該區域的沙漠穩定以及防風固沙發揮著重要作用。
在整個生長發育季節,對沙丘不同坡位倒披針葉蟲實種子萌發出苗及死亡情況進行對比研究發現,除沙面呈流動狀態的坡頂外,其他各坡位種子當年萌發出苗總量從丘間樣地到坡上樣地逐漸增加,這是由于沙丘中部和上部具有豐富的土壤種子庫,為倒披針葉蟲實在該坡位的分布提供了充足的種子資源。
荒漠一年生植物為適應環境波動,種子萌發采用兩頭下注的萌發對策[20-21],即一批生產的種子不會在一個生長季節全部萌發,而是分年和月份分批萌發。避免個體在一次降雨后隨之而來的干旱中全部死亡,這是極端生境下植物長期適應自然選擇的結果[22-23]。本研究發現,除坡頂外,其他坡位倒披針葉蟲實種子主要在3—6月間3批萌發出苗,不同時期種子萌發數量存在差異,3月份有大量種子萌發,4月份種子萌發數量最多,5月份種子萌發量逐漸減少。坡頂3月份種子萌發數量最多,4月至5月均無種子萌發。這可能受以下兩個因素影響:1)坡頂流動性高,種子被埋得過深,土壤中的低溫、低O2濃度、高濕和高CO2濃度會完全抑制種子萌發;2)種子萌發但由于沙埋過深幼苗沒有出土。倒披針葉蟲實種子的這種分批萌發策略對其種群的延續和更替具有非常重要的意義。
對于荒漠區來說降水是限制植物種子萌發的主要因素。在自然條件下,降水主要以兩種方式輸入到陸地生態系統——降雨和降雪,其中降雪是以固態的形式輸入到生態系統,其累積與消融對植物的生長發育有著重要的作用[24]。有研究表明古爾班通古特沙漠冬季積雪蒸發率極低。降雪及雪融水可以得到高效儲存,有78.8%—92%的雪融水可以轉化為土壤水,沙丘各部位在春季普遍有厚度不等的懸濕沙層。3月沙丘各坡位倒披針葉蟲實種子萌發主要利用的是融雪水。4—5月份由于日照強度增加、地溫上升沙層中的融雪水大量散失,干沙層開始形成。同時4—6月份是古爾班通古特沙漠風沙活動的頻繁期,對沙面較活化的中部和上部而言水分散失更快。而對研究區3—6月份的逐日降雨量進行分析發現,4、5月份均有一定份額的降雨,4、5月份種子萌發主要依賴于這部分降雨。因此,早春雪融水和春雨對倒披針葉蟲實種子萌發、幼苗定居和在沙丘不同坡向、不同坡位分布格局的形成過程中具有重要的作用和貢獻。
在一個生長季,植物種群數量呈現動態變化,是出苗和死亡綜合作用的結果。研究結果發現,3月下旬出土的幼苗,由于有早春融雪的支持,沒有死亡植株。4月份降雨,促使第2批種子大量萌發,植株死亡數量也極少。5月份雖然有降雨,但沙丘不同坡位倒披針葉蟲實植株死亡情況明顯不同,沙丘中部和上部死亡數量極少,而丘間和坡底幼苗大量死亡。造成該現象的原因可能是種間競爭和土壤種子庫對沙丘不同坡位種群動態起到重要的調節作用,沙丘中部和上部物種豐富度低,種間競爭弱,而同時具有大量的土壤種子庫,即使密度依賴的自疏導致植株死亡,土壤種子庫中的種子也能利用降雨補充大量的幼苗;丘間和坡底由于物種豐富度高,種間競爭強烈植株大量死亡,土壤種子庫密度低不能補充足夠的幼苗,致使種群數量呈下降的趨勢。幼苗經過兩個多月的生長發育,到6月初大部分植株已經開始成熟,種群密度逐漸趨于穩定。上述分析說明倒披針葉蟲實種群的續存與其種子庫中種子的分批萌發對策密切相關,并且同一個生長季內制約種群數量變動的關鍵因素會隨水分條件的不同而不同。另外,野外調查還發現倒披針葉蟲實種子萌發后由于缺水干燥死亡的種子,當重新吸水后幼根能夠繼續生長;黃振英等[25]的研究結果發現植株在喪失50%水分時仍能存活。這些特征能夠解釋倒披針葉蟲實在沙丘中部和上部呈優勢分布。研究發現植物群落幼苗存活與降雨量有關,尤其是荒漠一年生植物的種群動態可能是嚴酷的自然環境、競爭、生活史對策以及土壤特性等綜合作用的結果。此外,除3月份外,在生長發育的不同階段倒披針葉蟲實種群密度大小在沙丘各坡位總體上均表現為:沙面活化的中部和上部>沙面固定的丘間和坡底>流動的坡頂,不同坡位該植物植物種群密度不同還可能與土壤養分有關。
種子萌發后幼苗的生長和存活對于植物的豐富度和分布具有非常重要的意義,幼苗以及繁殖體的定居主要依賴于它們生存于合適的生境[26,27]。而植物的種群數量是出苗和死亡綜合作用的結果[28]。本研究中,在集中出苗的4月份,沙丘中上部密度最大達到(148±14)株/m2,下部(底部和丘間)的最大密度(64±14)株/m2。而生長季結束時,沙丘中上部最大密度為(147±14)株/m2,下部(底部和丘間)的最大密度為(43±12)株/m2,其種群密度往往小于集中出苗時的種群密度,并且種群密度從6月開始逐漸趨于穩定。沙丘不同坡位倒披針葉蟲實種群動態曲線表明,倒披針葉蟲實存活曲線屬于C型曲線,而具有C型存活曲線的種群其特點是早期具有很高的死亡率,需要有高出生率來補償,當到某一時間后,死亡率逐漸降低,種群密度趨于穩定[22,29]。同時,研究認為,在生存競爭中,k對策者以“質”取勝,而r對策者,則是以“量”取勝[30],本研究中,沙丘中部和上部單位面積植株產生種子數量極顯著高于丘間、坡底和坡頂。因此,倒披針葉蟲實是較為典型的r對策者。
倒披針葉蟲實在古爾班通古特沙漠廣泛分布,且在莫索灣地區其在沙丘各坡位的重要值是所有植物中最高的,該植物是一類抗逆性極強的沙生植物,對該地區沙面的穩定起到極其重要的作用。
[1]Goldblati P. An analysis of the flora of southern Africa: its characteristics, relationships, and orgins. Annals of the Missouri Botanical Garden, 1978, 65(2): 369-436.
[2]Ludwig J A, Cunningham G, Whitson P D. Distribution of annual plants in North American deserts. Journal of Arid Environments, 1988, 15: 221-227.
[3]Brown G. Species richness, diversity and biomass production of desert annuals in an ungrazedRhanteriumepapposumcommunity over three growth seasons in Kuwait. Plant Ecology, 2003, 165(1): 53-68.
[4]梁存柱, 劉鐘齡, 朱宗元, 王煒. 阿拉善荒漠區一年生植物層片物種多樣性及其分布特征. 應用生態學報, 2003, 14(6): 897-903.
[5]張榮, 劉彤. 古爾班通古特沙漠南部植物多樣性及群落分類. 生態學報, 2012, 32(19): 6056-6066.
[6]Grime J P. Plant Strategies, Vegetation Processes, and Ecosystem Properties. New York: John Wiley & Sons, 2006.
[7]張立運, 陳昌篤. 論古爾班通古特沙漠植物多樣性的一般特點. 生態學報, 2002, 22(11): 1923-1932.
[8]王雪芹, 蔣進, 雷加強, 張偉民, 錢亦兵. 古爾班通古特沙漠短命植物分布及其沙面穩定意義. 地理學報, 2003, 58(4): 598-605.
[9]Mosyakin S L. New subsections inCorispermumL. (Chenopodiaceae). Thaiszia, 1997, 7(1): 9-15.
[10]劉媖心. 中國沙漠植物志 (第一卷). 北京: 科學出版社, 1985.
[11]毛祖美, 張佃民. 新疆北部早春短命植物區系綱要. 干旱區研究, 1994, 11(3): 1-26.
[12]Mosyakin S L. New taxa ofCorispermumL. (Chenopodiaceae), with preliminary comments on the taxonomy of the genus in North America. Novon, 1995, 5(4): 340-353.
[13]Sukhorukov A P. Fruit anatomy and its taxonomic significance inCorispermum(Corispermoideae,Chenopodiaceae). Willdenowia, 2007, 37(1): 63-87.
[14]Robson D B. An analysis of the distribution, ecology, and status of bugseeds (Corispermum) in Canada. Canadian Field-Naturalist, 2010, 124(3): 246-255.
[15]He Y H, Zhao H L, Liu X P, Zhao X Y, Zhang T H, Draker S. Reproductive allocation ofCorispermumelongatumin two typical sandy habitats. Pakistan Journal of Botany, 2009, 41(4): 1685-1694.
[16]Huang Y X, Zhao X Y, Zhou D W, Luo Y Y, Mao W. Allometry ofCorispermummacrocarpumin response to soil nutrient, water, and population density. Botany, 2010, 88(1): 13-19.
[17]Wang Y H, He W M, Yu F H, Zhang L L, Cui Q G, Chu Y, Dong M. Brushing effects on the growth and mechanical properties ofCorispermummongolicumvary with water regime. Plant Biology, 2009, 11(5): 694-700.
[18]劉有軍, 劉世增, 紀永福, 馬全林, 張德魁, 張錦春, 魏林源. 碟果蟲實種子萌發對策及生態適應性. 生態學報, 2010, 30(24): 6910-6918.
[19]Liu S Z, Liu Y J, Ji Y F, Li Z Y, Zhang J H, Chen F. Seed germination ofCorispermumpatelliformein different storage length at room temperature: a dominant annual species in the deserts of northern China. Pakistan Journal of Botany, 2013, 45(3): 737-742.
[20]Venable D L. Bet hedging in a guild of desert annuals. Ecology, 2007, 88(5): 1086-1090.
[21]Donohue K, Rubio de Casas R, Burghardt L, Kovach K, Willis C G. Germination, postgermination adaptation, and species ecological ranges. Annual Review of Ecology, Evolution, and Systematics, 2010, 41: 293-319.
[23]Gremer J R, Venable D L. Bet hedging in desert winter annual plants: optimal germination strategies in a variable environment. Ecology Letters, 2014, 17(3): 380-387.
[24]范連連, 馬健, 吳林峰, 徐貴青, 李彥, 唐立松. 古爾班通古特沙漠南緣草本層對積雪變化的響應. 植物生態學報, 2012, 36(2): 126-135.
[25]黃振英, 吳鴻, 胡正海. 30種新疆沙生植物的結構及其對沙漠環境的適應. 植物生態學報, 1997, 21(6): 521-530.
[26]Jennifer A, Dunne V, Thomas P. Species-mediated soil moisture availability and patchy establishment ofPseudotsugamenziesiiin chaparral. Oecologia, 1999, 119(1): 36-45.
[27]曾曉玲, 劉彤, 沈雪瑩, 牛攀新. 古爾班通古特沙漠種子秋萌的環境依賴性. 生態學雜志, 2011, 30(8):1604-1611.
[28]楊慧, 婁安如, 高益軍, 宋宏濤. 北京東靈山地區白樺種群生活史特征與空間分布格局. 植物生態學報, 2007, 31(2): 272-282.
[29]尚玉昌, 蔡曉明. 普通生態學 (上冊). 北京: 北京大學出版社, 1992.
[30]張大勇. 植物生活史進化與繁殖生態學. 北京: 科學出版社, 2004.
Spatial and temporal distribution characteristics of ephemeralCorispermumlehmannianumin the Gurbantunggut Desert
CHEN Zhengxia, LIU Tong*, LIU Zunchi, HE Wenqin, TANG Jingguang, HAO Xiaoran, SUN Yuanyuan, ZENG Yong
CollegeofLifeScience,ShiheziUniversity,Shihezi832003,China
Corispermumlehmannianum, which ranks only second toHaloxylonpersicumin terms of Importance Value, is a valuable component of the desert flora. It is widely distributed in the Gurbantunggut Desert and plays a very important role in the stabilization of sand dunes. Consequently, this study investigated the distribution characteristics and dynamics of the population ofC.lehmannianumunder natural conditions, adopting the method of fixed quadrant. The results showed the following: 1)C.lehmannianumhas a higher Importance Value on all slope positions of the dunes, especially in the middle and upper parts of sand dunes where sand is more unstable compared with other slope positions. 2) Throughout the entire growth season, germination rate and emergence of seedlings on each slope position, aside from the top of the dunes, differentiates three groups. Different levels of seed germination are observed in different periods; a large number of seeds germinate and a large number of seedlings emerge in March. The highest levels of seed germination and emergence of seedlings occur in April, gradually decreasing in May. However, on the top of sand dunes, germination and emergence are the highest in March, whereas no germination and emergence were observed from April to May. Therefore, seed germination and population density on other slope positions, except at the top of sand dunes, increase, and then decrease. At the end of the growth season, population density demonstrates the following tendency in variation: the middle and upper parts of sand dunes > the lower part of sand dunes and interdunes > the top of sand dunes. 3) The seed yield per unit area of mature plants shows the same tendency as the population density. Seed number per unit area in the middle and upper parts of sand dunes are significantly higher than those in the interdunes, the bottom of the sand dunes, and the top of the sand dunes (P<0.01). 4) Given the seed bank densities at depths of 0—5 cm and 6—10 cm, the experimental results suggest that the number of viable seeds in the central and upper parts of sand dunes is significantly higher than those in other slope positions. In summary,C.lehmannianumare dominant in the middle and upper parts of sand dunes where sand is less stable compared with other slope positions of sand dunes and the species is well adapted to the desert environment. Thus,C.lehmannianumis one of the plants with the most potential to contribute to the stabilization of sand dunes.
ephemeral plant;Corispermumlehmannianum; temporal population dynamics; spatial population dynamics; seed bank
10.5846/stxb201411132248
國家自然科學基金-新疆聯合基金重點項目(U1130304);新疆生產建設兵團工業科技計劃項目(2012BA012)
2014-11-13; 網絡出版日期:2015-10-30
Corresponding author.E-mail: betula@126.com
陳正霞,劉彤,劉尊馳,何文琴,湯景光,郝曉冉,孫園園,曾勇.沙生植物倒披針葉蟲實在古爾班通古特沙漠的分布.生態學報,2016,36(13):4064-4073.
Chen Z X, Liu T, Liu Z C, He W Q, Tang J G, Hao X R, Sun Y Y, Zeng Y.Spatial and temporal distribution characteristics of ephemeralCorispermumlehmannianumin the Gurbantunggut Desert.Acta Ecologica Sinica,2016,36(13):4064-4073.
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
