17β-雌二醇調控大鼠成肌細胞L6GNR4細胞遷移能力的機制研究
2016-08-12 07:28:11彭召紅辛俊池王莉莉
實用藥物與臨床 2016年7期
吳 迪,彭召紅,辛俊池,王莉莉
?
·論著·
17β-雌二醇調控大鼠成肌細胞L6GNR4細胞遷移能力的機制研究
吳迪,彭召紅,辛俊池,王莉莉*
中國醫科大學附屬盛京醫院實驗研究中心,沈陽 110004
[摘要]目的研究17β-雌二醇對大鼠成肌細胞L6GNR4細胞遷移能力的調控作用及機制。方法L6GNR4細胞培養在含有10%胎牛血清的DMEM高糖增殖培養基中,培養基中添加10 nM雌激素17β雌二醇和/或50 nM雌激素競爭性拮抗劑Tamoxifen。應用細胞劃痕試驗在藥物處理后0、12、24 h觀察17β雌二醇對成肌細胞遷移能力的影響。在藥物處理24 h后,應用熒光定量PCR技術檢測各組細胞中雌激素受體α和β的表達。同時還檢測了成肌細胞遷移相關關鍵基因(Pax3、Fhl1以及Fhl1相互作用蛋白Actg1、Myh10)在各組細胞中的表達情況。結果與對照組比較,17β-雌二醇在劃痕24 h時可以明顯抑制L6GNR4的細胞遷移。17β-雌二醇能夠明顯抑制ERβ的表達,而雌激素競爭性拮抗劑Tamoxifen能夠部分逆轉17β-雌二醇對ERβ的抑制作用。17β-雌二醇能夠明顯抑制成肌細胞遷移相關的關鍵因子Pax3、Fhl1以及Fhl1相互作用蛋白Myh10的表達,而雌激素競爭性拮抗劑Tamoxifen能夠逆轉或部分逆轉17β-雌二醇對Pax3、Fhl1和Myh10的轉錄抑制作用。相同濃度的雌激素對L6GNR4細胞中Actg1的表達改變不明顯。結論雌激素17β-雌二醇通過ERβ轉錄抑制Pax3、Fhl1以及Fhl1的相互作用蛋白Myh10的表達,發揮其抑制大鼠成肌細胞遷移的作用。
[關鍵詞]17β-雌二醇;L6GNR4成肌細胞;細胞遷移
0 引言
骨骼肌組織是人體含量最多的肌肉組織,其構成和大小存在著明顯的性別差異。雌激素(17β-estradiol,E2)與雌激素受體(Estrogen receptor,ER)的配體結合結構域-雌激素受體反應元件結合后,發揮其調控靶基因轉錄的作用[1]。骨骼肌表達雌激素受體ERα和ERβ[2]。因此,雌激素可能在胚胎期肌肉發生、出生后骨骼肌發育以及骨骼肌損傷修復過程中發揮重要作用。肌肉發生指在胚胎發育過程中,體節細胞經過一系列增殖、遷移和分化,最終形成肌肉組織的過程[3]。①由胚胎的體節細胞出現細胞決定(分化方向),并分化為成肌細胞。②成肌細胞向胚胎肢芽遷移并保持增殖能力。③成肌細胞停止分裂,彼此融合成多核體細胞,并表達肌纖維特異性的肌球蛋白,使其分化為成熟的骨骼肌細胞。肌肉發生還包括了成體肌肉干細胞-骨骼肌衛星細胞的激活、分化、融合和成熟等過程。機體遇到肌肉損傷等刺激時,骨骼肌衛星細胞被激活,增殖并分化為成肌細胞。這些細胞再分化并融合進已存在的骨骼肌纖維中,或者彼此融合,形成新的肌纖維。L6GNR4細胞是來自大鼠骨骼肌的成肌細胞,其作為研究胚胎期肌肉發生和骨骼肌損傷修復的工具細胞,具有廣闊的應用前景和實用價值。目前,有關雌激素對成肌細胞遷移功能的影響的相關研究很少。因此,本研究應用17β-雌二醇處理大鼠L6GNR4成肌細胞,觀察其對成肌細胞遷移能力的影響及可能的機制。
1 材料與方法
1.1材料L6GNR4大鼠成肌細胞購自上海中科院細胞庫,17β雌二醇和雌激素競爭性拮抗劑Tamoxifen購自Sigma公司,定量PCR相關試劑購自TAKARA公司,細胞培養用血清和培養基等購自Gibco公司。
1.2細胞培養L6GNR4細胞培養在含有10%胎牛血清的DMEM高糖增殖培養基中,后者添加雌激素17β雌二醇和/或雌激素競爭性拮抗劑Tamoxifen。每組細胞重復實驗3次。根據Ogawa等[4]報道,本研究在成肌細胞培養基中添加10 nM 17β雌二醇和50 nM Tamoxifen。
1.3細胞劃痕實驗接種前用marker筆在6孔板背面畫橫線標記,拍照時定位同一個視野。每組細胞消化后接入6孔板,細胞長滿板底后,用20 μL槍頭垂直于孔板制造細胞劃痕。吸去細胞培養液,用PBS沖洗孔板3次,洗去劃痕產生的細胞碎片。加入無血清培養基,不同時間點拍照記錄。
1.4定量PCR用TRIZOL試劑提取各組細胞的總RNA。用反轉錄酶和Olingo (dT) 20引物合成cDNA。定量PCR的引物序列見表1。在定量PCR過程中,每個基因重復擴增3次,基因的相對表達水平應用2-△△ct方法計算。PCR產物經2%瓊脂糖膠分離并拍照。
1.5統計學分析應用SPSS 10.0軟件統計分析各組細胞間基因表達水平是否存在差異。
2 結果
2.117β-雌二醇抑制大鼠成肌細胞L6GNR4細胞遷移在細胞劃痕實驗中,連續觀察24 h 17β-雌二醇對大鼠成肌細胞L6GNR4遷移能力的影響,結果顯示,與對照組比較,17β-雌二醇在劃痕24 h時,可以明顯抑制L6GNR4的細胞遷移(見圖1)。細胞劃痕實驗重復3次,每次的實驗結果一致。2.217β-雌二醇抑制大鼠成肌細胞L6GNR4中ERβ的表達大鼠成肌細胞L6GNR4中,有雌激素受體ERα和ERβ的表達,并且以ERβ為主(ERα相對表達水平為1時,ERβ的相對表達水平為3.05±0.55)。各組細胞ERα的相對表達水平比較差異無統計學意義(數據未顯示)。對照組、E2組、Tam組、E2+Tam組細胞ERβ的相對表達水平分別為0.99±0.005、0.17±0.09、0.78±0.02、0.38±0.01。結果顯示,17β-雌二醇能夠明顯抑制ERβ的表達(對照組 vs.E2組,P=0.0020),而Tamoxifen能夠部分逆轉17β-雌二醇對ERβ的抑制作用(E2組 vs.E2+Tam組,P=0.0199)。
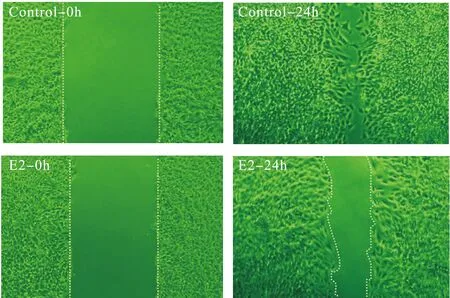
圖1 17β-雌二醇抑制大鼠成肌細胞L6GNR4細胞遷移
2.317β-雌二醇抑制大鼠成肌細胞L6GNR4中調節細胞遷移的關鍵基因的表達為了探索17β-雌二醇抑制大鼠成肌細胞L6GNR4細胞遷移的機制,對成肌細胞遷移相關的關鍵因子Pax3和Fhl1以及Fhl1相互作用蛋白Actg1、Myh10在各組細胞中mRNA的表達水平[4]。Pax3相對表達水平:對照組1.00±0.09,E2組0.51±0.05,Tam組1.20±0.06,E2+Tam組1.21±0.03(對照組 vs.E2組,P=0.000 1;E2組 vs.E2+Tam組,P=0.002 7)。Fhl1的相對表達水平:對照組1.00±0.12,E2組0.37±0.12,Tam組0.97±0.00,E2+Tam組1.00±0.05(對照組 vs.E2組,P=0.0305;E2組 vs.E2+Tam組,P=0.029 4)。Actg1的相對表達水平:對照組1.00±0.03,E2組0.81±0.12,Tam組0.99±0.03,E2+Tam組1.02±0.08(對照組 vs.E2組,P=0.050 8;E2組 vs.E2+Tam組,P=0.059 7)。Myh10的相對表達水平:對照組1.00±0.03,E2組0.03±0.01,Tam組0.98±0.11,E2+Tam組0.56±0.01(對照組 vs.E2組,P=0.0003;E2組 vs.E2+Tam組,P=0.0046)。結果顯示,17β-雌二醇能夠明顯抑制成肌細胞遷移相關的關鍵因子Pax3、Fhl1及Myh10的表達(P<0.05),而Tamoxifen能夠逆轉17β-雌二醇對Pax3、Fhl1的轉錄抑制作用。同時Tamoxifen能夠部分逆轉17β-雌二醇對Myh10的轉錄抑制作用。相同濃度的雌激素對L6GNR4細胞中Actg1的表達改變不明顯,見圖2、圖3。
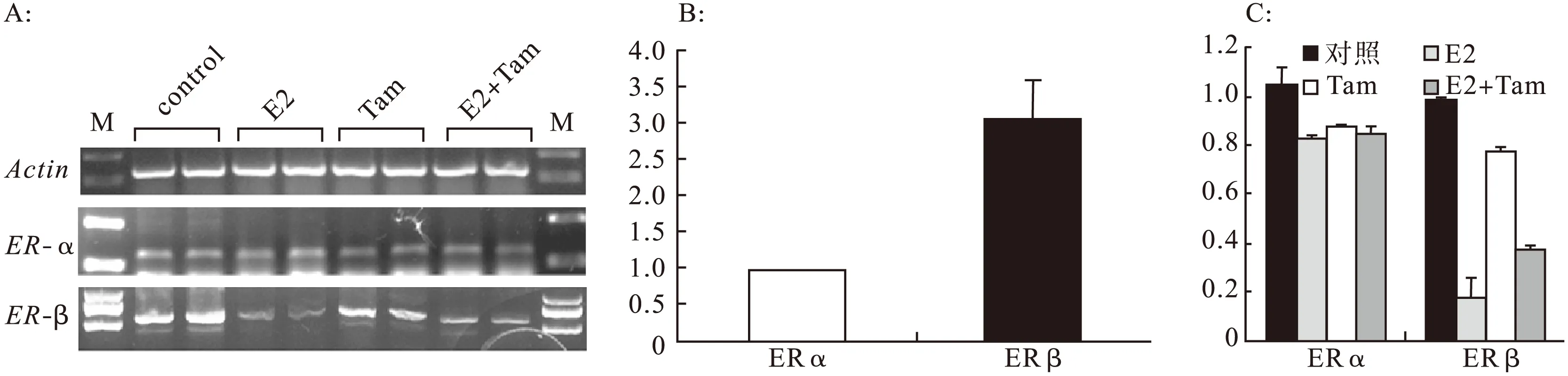
圖2 17β-雌二醇抑制大鼠成肌細胞L6GNR4ERβ的表達

圖3 17β-雌二醇抑制大鼠成肌細胞L6GNR4中關鍵細胞遷移相關因子的表達

基因引物序列長度(bp)復性溫度(℃)ActinGGAGATTACTGCCCTGGCTCCTAGACTCATCGTACTCCTGCTTGCTG13956ERαCGATGGTGCATTGGTTTGTGAGACTCGCTACTGTGCTGTG14160ERβGCCCTTGTTACTGATGTGCCATGCCCTGGTCTGGGTGATT11260Pax3CTGCTCTGCGGTGAAGGTGGAGGAGGAGGCGGATTTAG19160Fhl1CGTGCCAGTAGCGATTCTTATGCTGCCTGAAGTGCTTTGAC10756Actg1AATGCCGTGCTCAATAGGGTCGCAATGGAAGAAGAAATCG22960Myh10ACTTGCCAAAGCGAGATGAGACAAAGGAAGAAAGGACCAC12960
3 討論
雌激素和ERα的配體結合結構域-雌激素受體反應元件結合后發揮其調控靶基因轉錄的作用。而ERβ不僅能和雌激素受體反應元件結合發揮其轉錄因子的作用,還能夠以不依賴配體結合的方式發揮其作用[2]。ERα和ERβ競爭性地結合同一個雌激素受體反應元件,或者特異地結合在不同雌激素受體反應元件上,因此其在調控基因表達的功能上也不相同[5-6]。大鼠成肌細胞L6GNR4中有雌激素受體ERα和ERβ的表達,并且以ERβ為主。研究表明,17β-雌二醇可以抑制成肌細胞中骨骼肌分化標記物(Myosin heavy chain,MHC)、Tropomyosin和Myogenin的表達水平,并且可以抑制成肌細胞融合形成肌管的過程[7]。但Galluzzo等[8]研究結果表明,17β-雌二醇可以提高大鼠成肌細胞L6GNR4中Myogenin和MHC的表達。17β-雌二醇還可以通過PI3K/Akt信號通路抑制成肌細胞的凋亡[9]。目前,關于雌激素對成肌細胞遷移功能影響的相關研究很少。因此,本研究應用17β-雌二醇處理大鼠成肌細胞,觀察其對成肌細胞遷移能力的影響及可能的機制。
在細胞劃痕實驗中,筆者連續觀察24 h 17β-雌二醇對大鼠成肌細胞L6GNR4遷移能力的影響,結果顯示,17β-雌二醇在劃痕24 h時,可以明顯抑制L6GNR4的細胞遷移。同時,檢測17β-雌二醇對成肌細胞中不同雌激素受體亞型和調節細胞遷移的關鍵基因(Pax3、Fhl1等)表達的影響。結果表明,17β-雌二醇能夠明顯抑制ERβ的表達,而Tamoxifen能夠部分逆轉17β-雌二醇對ERβ的抑制作用。17β-雌二醇能夠明顯抑制成肌細胞遷移相關的關鍵因子Pax3、Fhl1及Myh10的表達,而Tamoxifen能夠逆轉或部分逆轉17β-雌二醇對Pax3、Fhl1及Myh10的轉錄抑制作用。
Pax3、Fhl1及Myh10在骨骼肌發育中發揮重要作用。在胚胎期,肌肉前體細胞從軸旁體節遷移至其他形成肌肉組織的部位,如:肢體。肌肉前體細胞的遷移在肌肉發生中發揮重要作用,而Pax3通過調節c-Met參與調節肌肉前體細胞的遷移過程[10-11]。在小鼠骨骼肌So18細胞中,重組表達的Fhl1可以調節integrin介導的細胞骨架的重排。細胞表面integrin激活直接導致細胞骨架actin重組,同時伴有粘著斑復合物和彈力纖維的形成。在成肌細胞中GFP-Fhl1的過表達可以通過-5-1-integrin特異地抑制成肌細胞粘附力,并提高細胞的擴散和遷移能力[12]。Non-muscle myosin 2B(Myh10)和actin細胞骨架的結合在細胞的運動、黏附和細胞形態的維持中發揮重要作用[13-14]。有關Non-muscle myosin在骨骼肌發育中的作用的相關研究很少。有研究表明,在成肌細胞中,non-muscle myosins 2A/2B和actin的相互作用能夠改變成肌細胞的形態、黏附和運動能力。此外,non-muscle myosin 2B敲除后,可以明顯抑制成肌細胞的尾部收縮,增加細胞長度,干擾肌管形成過程中細胞核的重分布[15]。
綜上所述,雌激素17β-雌二醇通過ERβ轉錄抑制Pax3、Fhl1及Myh10的表達,以發揮抑制大鼠成肌細胞遷移的作用。
參考文獻:
[1]Vivar OI,Zhao X,Saunier EF,et al.Estrogen receptor beta binds to and regulates three distinct classes of target genes[J].J Biol Chem,2010,285(29):22059-22066.
[2]Wiik A,Ekman M,Johansson O,et al.Expression of both oestrogen receptor alpha and beta in human skeletal muscle tissue[J].Histochem Cell Biol,2009,131(2):181-189.
[3]Wang LL,Peng ZH,Fan Y,et al.Dynamic expression of molecules that control limb muscle development including Fhl1 in hind limbs of different gestational age[J].APMIS,2014,122(9):766-771.
[4]Wang L,Miao J,Li L,et al.Identification of an FHL1 protein complex containing gamma-actin and non-muscle myosin IIB by analysis of protein-protein interactions[J].PLoS One,2013,8(11):e79551.
[5]Hayashi SI,Eguchi H,Tanimoto K,et al.The expression and function of estrogen receptor alpha and beta in human breast cancer and its clinical application[J].EndocrRelat Cancer,2003,10(2):193-202.
[6]Lazennec G,Bresson D,Lucas A,et al.ER beta inhibits proliferation and invasion of breast cancer cells[J].Endocrinology,2001,142(9):4120-4130.
[7]Ogawa M,Yamaji R,Higashimura Y,et al.17β-estradiol represses myogenic differentiation by increasing ubiquitin-specific peptidase 19 through estrogen receptor α[J].J Biol Chem,2011,286(48):41455-41465.
[8]Galluzzo P,Rastelli C,Bulzomi P,et al.17beta-Estradiol regulates the first steps of skeletal muscle cell differentiation via ER-alpha-mediated signals[J].Am J Physiol Cell Physiol,2009,297(5):1249-1262.
[9]La Colla A.Vasconsuelo AA.Boland R.Estradiol exerts antiapoptotic effects in skeletal myoblasts via mitochondrial PTP and MnSOD[J].J Endocrinol,2013,216(3):331-341.
[10]Birchmeier C,Brohmann H.Genes that control the development of migrating muscle precursor cells[J].Curr Opin Cell Biol,2000,12(6):725-730.
[11]Buckingham M,Relaix F.PAX3 and PAX7 as upstream regulators of myogenesis[J].Semin Cell Dev Biol,2015,44:115-125.
[12]Robinson PA,Brown S,Mc Grath MJ,et al.Skeletal muscle LIM protein 1 regulates integrin-mediated myoblast adhesion,spreading,and migration[J].Am J Physiol Cell Physiol,2003,284(3):681-695.
[13]CramerLP.Organization and polarity of actin filament networks in cells:implications for the mechanism of myosin-based cell motility[J].BiochemSoc Symp,1999,65:173-205.
[14]Conti MA,Adelstein RS.Nonmuscle myosin II moves in new directions[J].J Cell Sci,2008,121(Pt 1):11-18.
[15]Swailes NT,Colegrave M,Knight PJ,et al.Non-muscle myosins 2A and 2B drive changes in cell morphology that occur as myoblasts align and fuse[J].J Cell Sci,2006,119(Pt 17):3561-3570.
收稿日期:2016-03-22
基金項目:國家自然科學基金(81100434,81471466)
*通信作者
DOI:10.14053/j.cnki.ppcr.201607001
Mechanism of 17β-estradiol regulating the migration of L6GNR4 myoblast
WU Di,PENG Zhao-hong,XIN Jun-chi,WANG Li-li*
(Medical Research Center,Shengjing Hospital of China Medical University,Shenyang 110004,China)
[Abstract]ObjectiveTo investigate the effect of 17β-estradiol on the migration of L6GNR4 myoblast and discuss the molecular mechanism.MethodsL6GNR4 cells were cultured in high-glucose DMEM containing 10 nM 17β-estradiol and/or 50 nM tamoxifen.The effect of 17β-estradiol on the migration of L6GNR4 myoblasts was detected by Wound-healing assays at 0 h,12 h and 24 h after medication.The expression levels of ER and key genes involved in myoblast migration (Pax3,Fhl1,Fhl1 interactor-Actg1 and Myh10) were examined by real-time RT-PCR in each group.Results17β-estradiol inhibited the migration of myoblast after 24 h and down-regulated the expression of ERβ,Pax3,Fhl1 and Myh10 which could be totally or partly reversed by tamoxifen,while the same dose of estrogen has no obvious effect on Actg1 expression in L6GNR4.Conclusion17β-estradiol can inhibit the expression of Pax3,Fhl1 and Myh10 by ERβ-mediated transcription,thus restraining the L6GNR4 myoblast migration
Key words:17β-estradiol;L6GNR4 myoblast;Migration
