中國黑戈壁植物多樣性分布格局及其影響因素
2016-08-09 05:52:13王健銘董芳宇巴海那斯拉李景文李俊清馮益明
生態學報 2016年12期
王健銘,董芳宇, 巴海那斯拉,3,李景文,*, 李俊清,馮益明, 盧 琦
1 北京林業大學林學院,北京 100083 2 中國林業科學研究院荒漠研究所,北京 100091 3 新疆重點林業工程質量管理總站, 烏魯木齊 830000
?
中國黑戈壁植物多樣性分布格局及其影響因素
王健銘1,董芳宇1, 巴海那斯拉1,3,李景文1,*, 李俊清1,馮益明2, 盧琦2
1 北京林業大學林學院,北京100083 2 中國林業科學研究院荒漠研究所,北京100091 3 新疆重點林業工程質量管理總站, 烏魯木齊830000
摘要:我國西北地區內陸分布著近20萬km2的黑戈壁,由于其環境的特殊性,使其具有獨特的生態系統,境內分布著多樣的植被,蘊藏著大量特有的自然資源。但由于自然環境苛刻與交通條件不便,目前我國關于黑戈壁區系統的植被與物種多樣性的研究還很缺乏。針對黑戈壁區植物多樣性組成與分布特點,基于遙感及實地調查,采用DCCA排序和半變異函數模型等分析方法,對黑戈壁區植物群落組成,植物多樣性特點及影響因素進行分析。研究結果表明:研究區植物以藜科和蒺藜科灌木或半灌木為主,群落物種生活型具有逐漸趨于簡單甚至單一的特性,重要值>0.1的植物主要有梭梭、紅砂、白刺等13種;群落物種多樣性呈現區域性的斑塊化分布,結構性因子引起的物種多樣性空間異質性占主導地位;作為極端干旱區,該區植物群落類型具有貧乏化及單一化的趨勢,群落結構簡單,植被覆蓋度低,植物生長隨環境的變化具有明顯的可塑性,群落空間分異明顯,群落空間演變具有明顯的水分及土壤結構梯度;DCCA結果顯示氣候、土壤、地形是群落物種及類型變化的主要原因,海拔、坡位、土壤機械組成、降水、溫度等環境因子對群落有著顯著的影響,水土條件的空間異質性是戈壁植物多樣性維持的關鍵因素。
關鍵詞:黑戈壁;植物群落;物種多樣性;分布;因素
植物多樣性作為群落的重要特征,是生態系統結構、功能及其穩定性的基礎[1- 2]。隨著生態研究的不斷深入,植物多樣性格局形成機制研究成為了生態學熱點問題[3-4],該方面的研究是理解區域生物多樣性維持機制重要的內容;同時對生物多樣性的保護管理等有著重要意義[5-8]。我國在生物多樣性格局與維持機制方面開展了很多卓有成效的研究,但對于西北干旱區,特別是自然環境嚴苛,交通不便的極端干旱的戈壁區,這方面工作還很缺乏。目前關于戈壁植物多樣性的研究仍然較少,國內相關研究主要是植被地理與植物分布[9-10]、植物區系特點[11-12],以及周邊區域的相關研究[13- 14],近年來,德國、日本學者在我國西北和蒙古國,主要是在蒙古國一些戈壁地區開展了一些研究工作,包括植被及物種多樣性分布與環境因子關系[15-18],氣候變化對群落生產力影響以及植被生長與沙塵發生關系[18-21]等。而黑戈壁區至今還沒有系統的植物多樣性分布格局的研究。
黑戈壁區位于甘肅與新疆之間,為東起額濟納河,北抵中蒙界山,南臨河西走廊西段,西依天山東段的一個大約20萬km2的區域。該區年降水量在100mm以下,多數區域低于50mm,蒸發量極高,為我國極端干旱區。受嚴苛環境及交通不便的影響,目前黑戈壁這一戈壁荒漠中最干旱區域的植被與物種多樣性格局的系統研究尚未開展。而隨著西部開發及絲綢之路經濟帶發展,正在建設的京新高速公路及鐵路直穿黑戈壁腹地,同時礦產資源開發范圍擴大,人為干擾對該區脆弱的生態系統及其植被的影響日益嚴重。未來該區自然環境維護和植被及其物種多樣性的保護也急需全面系統的本底調查。
針對我國黑戈壁植物多樣性分布格局的研究缺乏問題,從2011年開始由中國林科院荒漠研究所牽頭,組織了多個單位對黑戈壁地區進行了包括地貌特征、水文、土壤、氣候與動植物資源等生態本底的綜合考察[22]。本文在黑戈壁區近2年的植物多樣性全面調查基礎上,重點分析黑戈壁區植物多樣性分布格局及其主要影響因素,闡明黑戈壁植物多樣性分布特征及其特殊性;為黑戈壁區植物多樣性保護、區域生態環境維持提供理論依據。
1研究方法
1.1調查區域與線路的確定
本研究依據中國黑戈壁綜合科學考察線路,設計植被調查的地區和線路,調查區域盡量覆蓋黑戈壁不同地理區域、地形地貌特征、植被類型等,于2011—2012年生長季沿著調查線路對甘肅敦煌、肅北,新疆哈密、伊吾、巴里坤,內蒙額濟納旗等典型黑戈壁區進行植物本底調查,調查線路總長度達到4900km。
1.2調查方法
依據規劃的調查線路,在植被分布區內,根據植被組成、水土條件的變化梯度布設樣點,在每個樣點內根據植被分布面積大小和組成復雜程度設置3—5個10m×10m的臨時樣方,共設置樣點56個,臨時樣方174個;在每個臨時樣方內調查植被類型、物種的組成。植物物種調查指標包括地理坐標、海拔、地形特征、戈壁類型[22]、礫石覆蓋度與粒級,群落特征包括群落類型、群落總蓋度、植物物種蓋度、主要物種組成、個體數量。記錄樣方內所有植物的種類、株數、蓋度、平均高度、生長狀況、物候、分布情況等。
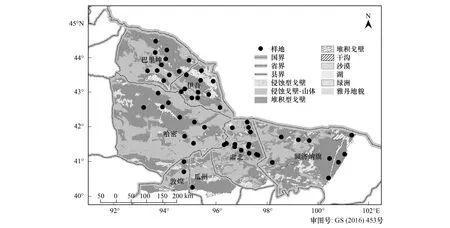
圖1 調查樣點分布圖Fig.1 Distribution of survey samples
2數據處理
2.1群落特征數據及環境相關性分析
依據《中國植被》[23]的分類原則,依據生境特點、群落結構與外貌、物種優勢度及物種組成劃分植物群落[24]。根據臨時樣方內植物個體高度、蓋度、密度以及頻度,計算物種的重要值:重要值=(相對密度+相對頻度+相對蓋度+相對高度)/4[5- 7,25];相對密度=某一種個體數/所有種的個體數總和,相對蓋度=某物種的蓋度/全部種的蓋度總和,相對高度=某一種植株平均高度/所有種植株平均高度總和,相對頻度=某物種的頻度/所有種的頻度之和。依據物種多樣性測度指數應用的廣泛程度以及對群落物種多樣性狀況的反映能力,本文選取以下4種多樣性指數來測度和分析群落物種多樣性特征[5- 7,25]:
(1)Simpson指數(D)
(2)Shannon-Wiener 指數(H)
(3)Pielou 均勻度指數(J)
J=H/lnS
(4)Margalef豐富度指數(M)
M=(S-1)/lnN
式中,S為樣方內物種數;N為樣方內所有物種個體數總和;Pi為種i的相對重要值。
利用SPSS 22軟件對數據進行均值分析,并對群落環境因子及植物群落特征與多樣性指數等數據進行相關性系數及顯著性分析。
2.2植物群落空間分布格局及環境相關性分析
因樣方中分布頻率較低物種的存在,會影響排序結果,使最終解釋與實際有所偏差[25- 27],所以本文在排序分析時,利用CANOCO 4.5軟件設置處理的方法剔除樣方分布頻率小于5%的植物[13,25- 27]。最后得到162(樣方)×36(物種)維物種數據矩陣。環境數據包括年平均溫度、最熱月均溫、年降水量、海拔、坡度、坡位、礫石蓋度、礫石徑級等8個環境因子,得到162(樣方)×8(環境變量)維的環境數據矩陣;其中氣候數據來取自于中國黑戈壁綜合科學考察隊氣候組數據庫[22]。實際分析中使用CANOCO 4.5軟件包中的蒙特卡羅擬合(Monte Carlo permutation test)檢驗植物群落組成與環境變量之間的相關顯著性,以及運用偏典范對應分析(pDCCA)分離環境變量對植物群落變異的相對貢獻率[26- 28]。
2.3群落物種多樣性空間格局及多樣性預測分析
利用GS+9.0軟件對群落物種多樣性空間分布進行半變異函數與方差模型分析,并在模型和方差分析的基礎上,在ArcGIS 10.2上,利用普通克里金插值法,繪制黑戈壁物種多樣性預測圖。
3結果與分析
3.1黑戈壁植物種類及群落數量特征分析
調查中記錄黑戈壁區植物154種,分屬28科,85屬,重要值與分布頻度較高的植物主要有梭梭(Haloxylonammodendron)、紅砂(Reaumuriasongarica)、芨芨草(Achnatherumsplendens)、白刺(Nitrariatangutorum)、沙拐棗(Calligonummongolicum)、膜果麻黃(Ephedraprzewalskii)、霸王(Sarcozygiumxanthoxylon)、合頭草(Sympegmaregelii)、泡泡刺(Nitrariasphaerocarpa)、戈壁藜(Iljiniaregelii)、鹽生草(Halogetonglomeratus)、蘆葦(Phragmitesaustralis)、檉柳(Tamarixhohenackeri)等13種植物,為黑戈壁主要優勢植物(表1)。總體上,黑戈壁優勢植物以藜科和蒺藜科灌木或半灌木為主,草本所占的比例很小,反映了黑戈壁環境嚴苛和異質性程度低的條件下,植被組成物種生活型逐漸趨于簡單甚至單一的特性。
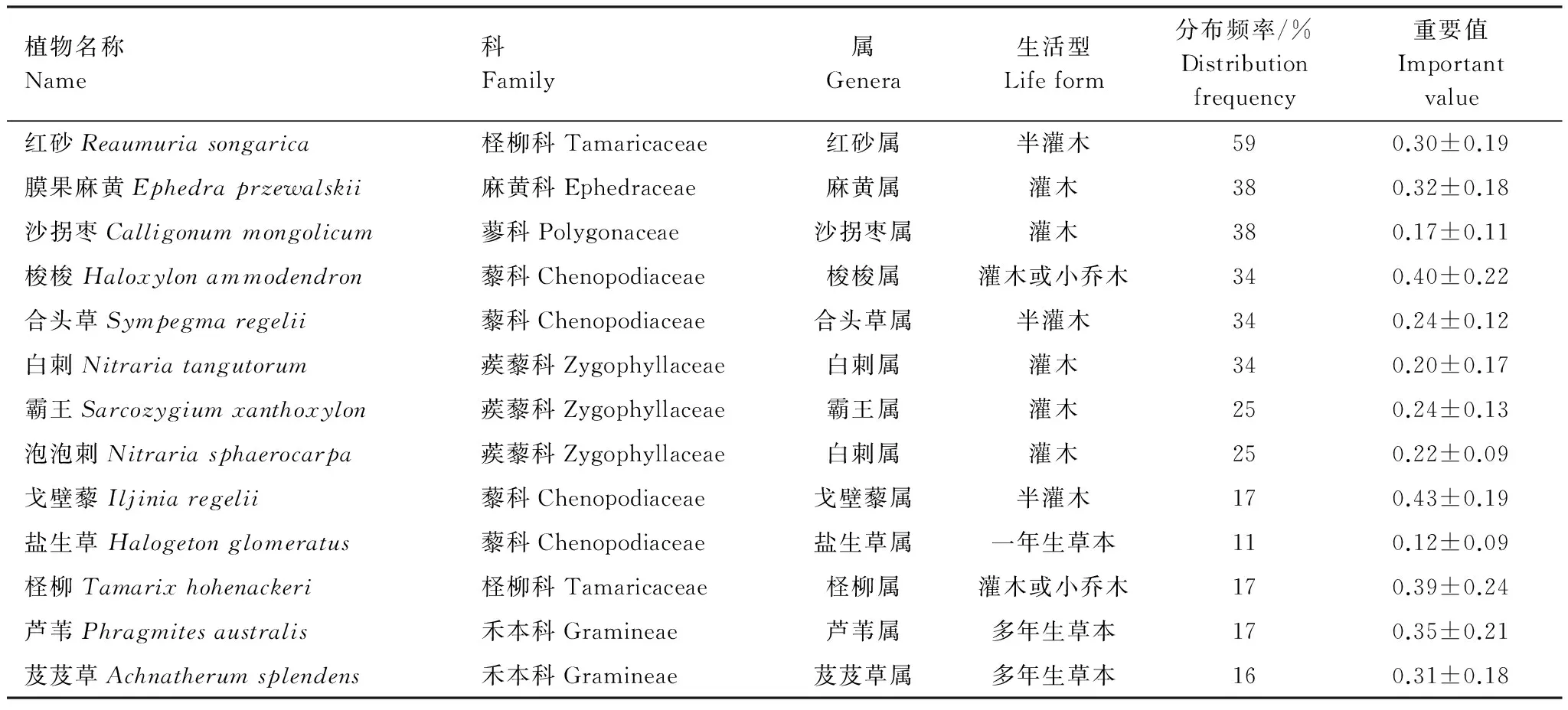
表1 優勢植物種類及其重要值
紅砂在所有樣方中出現的頻率最高,是分布范圍最廣的物種,梭梭、膜果麻黃、沙拐棗、白刺等植物也是黑戈壁主要的優勢物種,在樣方內出現頻率也較高;但在不同生境中植物的優勢度及生長指標差異很大,反映了植物對不同環境梯度具有生長適應性。檉柳、蘆葦等群落,主要分布在季節性積水河道、湖盆或有地質斷裂帶的泉水生境中,屬于該區隱域性植被,分布范圍小,生境較為特殊。而戈壁針茅(StipatianschanicaRoshev vargobica)等主要分布在黑戈壁海拔相對較高的邊緣地區及荒漠草原的過渡區內。
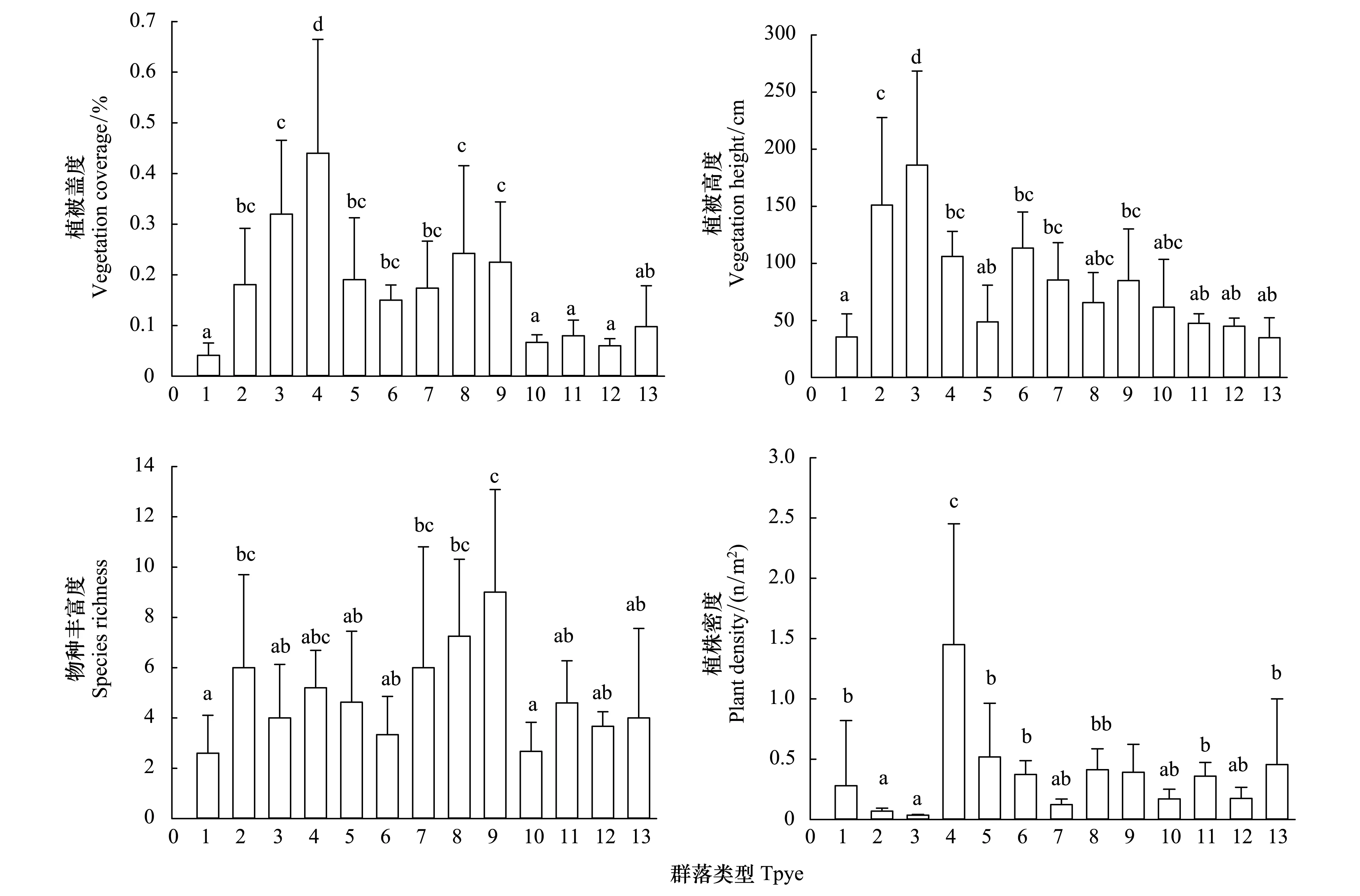
圖2 主要群落數量特征Fig.2 Quantitative characteristics of the major plant communities1—13分別代表:戈壁藜、梭梭、檉柳、蘆葦、紅砂、芨芨草、膜果麻黃、合頭草、霸王、白刺、泡泡刺、沙拐棗、鹽生草群落
如圖2所示,不同優勢群落間植被蓋度及高度、群落植株密度存在極顯著的差異(P<0.01),物種豐富度也存在顯著差異(P<0.05),戈壁藜、白刺、泡泡刺、鹽生草等群落主要分布在地表粗糙度高的礫石堆積戈壁,植被稀疏,群落覆蓋度低,群落植物個體密度變異明顯;霸王、合頭草等群落多分布在海拔較高、降水相對豐富的地區,其植被生長較為良好,物種豐富度也較高;而檉柳、蘆葦等隱域性植被,多分布在礫石覆蓋少的地勢較低區域,其植被高度和蓋度情況最好。
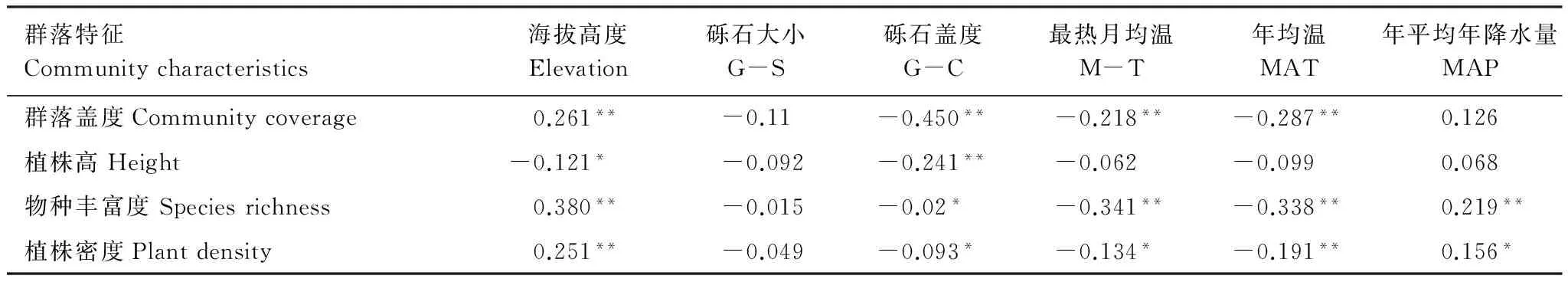
表2 群落數量特征與環境因子Pearson相關性分析
**P<0.01, *P<0.05
由表2可知,群落物種豐富度與溫度、礫石蓋度呈顯著負相關,與年平均降水量及海拔呈正相關;群落植被蓋度與海拔呈顯著正相關,與溫度及礫石蓋度呈負相關;其植被高度僅與海拔高度及礫石蓋度呈負相關,與其它環境因子則沒有顯著關系;環境因子與群落植株密度的關系和其與植被蓋度的關系相似,不同之處在于群落植株密度還與降水呈正相關。這表明群落數量特征與海拔、地表覆蓋物、水熱條件有著密切的關系,地形及大氣路徑導致的水熱條件在空間上的異質性[22]對植物群結構和組成存在顯著影響,植物地上部分隨著環境梯度的變化具有生長(蓋度及高度)的可塑性。
3.2黑戈壁植物群落DCCA排序分析
通過對數據進行DCCA排序分析,并在CANOCO 4.5軟件包中對排序軸進行蒙特卡羅擬合檢驗,結果顯示物種變量與環境變量之間呈現極顯著的相關性(P=0.0001),表明環境因子對群落的物種組成具有顯著的影響。與此同時,對8個環境變量進行Monte Carlo檢驗,檢驗變量是否存在冗余現象,結果顯示8個環境因子均與植物群落存在顯著的相關性,不存在冗余現象(P<0.001)。

圖3 DCCA植物物種排序圖Fig.3 Plant species ordination diagram of DCCA圖中列出了物種加權值大于10%的優勢物種:Ⅰ,泡泡刺Nitraria sphaerocarpa;Ⅱ,白刺Nitraria tangutorum;III,灌木亞菊Ajania fruticulosa;IV,短葉假木賊Anabasis brevifolia;Ⅴ,五柱紅紗Reaumuria kaschgarica ;Ⅵ,紅砂Reaumuria songarica;Ⅶ,松葉豬毛菜Salsola laricifolia ;Ⅷ,檸條錦雞兒Caragana Korshinskii;IX,白沙蒿Artemisia blepharolepis ;Ⅹ,膜果麻黃Ephedra przewal;Ⅺ,合頭草Sympegma regelii;Ⅻ,芨芨草Achnatherum splendens;,木本豬毛菜Salsola arbuscula;,蘆葦Phragmites australis;XV,脹果甘草Glycyrrhiza inflata ;XVI,檉柳Tamarix hohenackeri;XVII,黑果枸杞Lycium ruthenicum;XVIII,戈壁針茅Stipa tianschanica Roshev var gobica;XIX,密頭蒿Tanacetum crassipes;XX,沙拐棗Calligonum mongolicum;XXI,駱駝刺Alhagi sparsifolia ;XXII,花花柴Karelinia caspia;XXIII,鹽生草Halogeton glomeratus;XXIV,戈壁藜Iljinia regelii;XXV,梭梭Haloxylon ammodendron;;XXVI,霸王Sarcozygium xanthoxylon;XXVII,甘青鐵線蓮Clematis tangutica;XXVIII,駝絨藜Ceratoides laten;XXIX,白皮錦雞兒Caragana leucophloea;XXX,木旋花Convolvulus fruticosus;A-T,年均溫;M-T:最熱月均溫;A-P:年平均降雨量;Elevation:海拔;G-S:礫石大小;Slope:坡度:G-T:坡位;G-C:礫石蓋度
采用8個環境變量,對研究區植物群落格局與環境的關系進行排序分析,結果顯示第一和第二軸的特征值分別為0.655、0.396,貢獻率分別為0.492、0.297,前兩軸特征值之和占全部特征值總和的78.9%,包含了絕大多數的排序信息,因此采用前兩軸的數據來分析環境因子與植物群落的關系。
從圖3可以看出,DCCA排序圖第一軸主要反映海拔、氣候、坡位等環境因子對植物分布的影響;其中海拔對植物群落分布影響最顯著, 地形是植物所處環境氣候、地表覆蓋度產生差異的重要原因,蘆葦、檉柳、花花柴(Kareliniacaspia)等植被主要分布在地勢較低、溫度較高的季節性河流、湖盆生境,紅砂、梭梭等典型的戈壁植物則分布在廣闊的山前戈壁生境上,而戈壁針茅、密頭蒿(Tanacetumcrassipes)、灌木亞菊(Ajaniafruticulosa)、五柱紅砂(Reaumuriakaschgarica)等植物主要分布在海拔較高、降水較好的生境,形成戈壁向草原過渡的植被。第二軸主要反映了地形起伏、地表覆蓋、土壤質地對植物群落空間分布的影響;戈壁針茅、芨芨草、檸條錦雞兒(Caraganakorshinskii) 等植物主要分布在地形起伏小、礫石較少的地區,白皮錦雞兒(Caraganaleucophloea)、戈壁藜、短葉假木賊(Anabasisbrevifolia)、木旋花(Convolvulusfruticosus)等植物等分布在地形起伏較大、地表覆蓋粗糙的生境中。
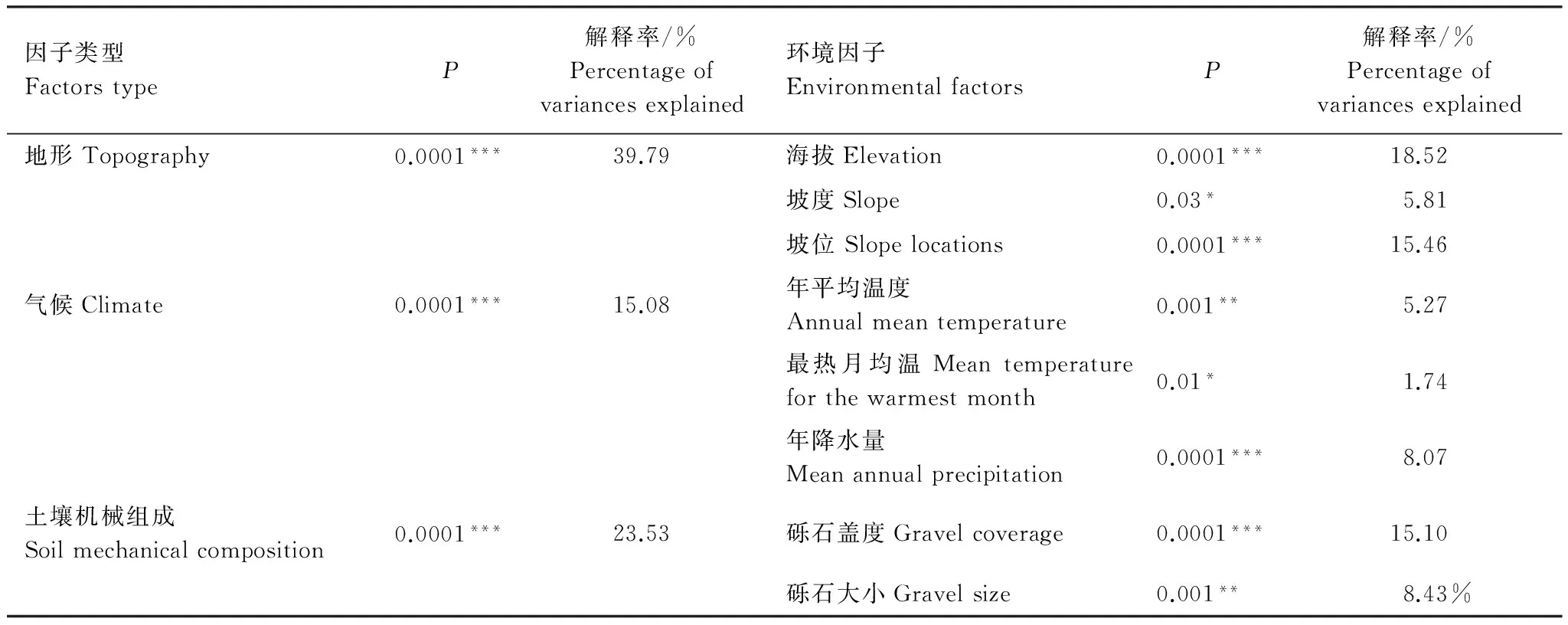
表3 環境因子對植物群落影響的Monte Carlo檢驗及相對貢獻率分析
***P<0.001,**P<0.01,*P<0.05
運用偏典范對應分析(pDCCA)分離環境變量對植物群落變異的解釋能力,并將8個環境因子分為地形、氣候、土壤3部分,來分析各環境因子對植物群落格局的影響[29]。由研究結果可知,地形(39.79%)是影響黑戈壁群落格局的主控因子,此外土壤(23.53%)、氣候(15.08%)對群落格局也有著顯著的影響;單個環境因子中,海拔(18.52%)、坡位(15.46%)、礫石蓋度(15.1%)對群落的影響最明顯,年平均年降水量(8.07%)、礫石大小(8.43%)、溫度(7.01%)也對群落格局存在重要作用,此外未解釋部分為48.1%,可能包括土壤化學性質、生物相互作用、干擾因素和隨機因素等。
3.3植物群落物種多樣性分布格局分析3.3.1黑戈壁植物物種多樣性水平變化分析
從圖4可以看出,多數樣方內物種多樣性指數都比較低,不同生境下樣方多樣性水平存在明顯的差異。經向變化規律表現為多樣性指數整體連續波動,變化劇烈,在92.6°E、93.3°E、93.9°E、96.1°E、97.5°E、100.5°E形成6個物種多樣性高值區,在93.9E附近達到了最高值,而最低值則出現在100.9°E附近。從緯向變化上分析,多樣性指數同樣持續波動變化,在40.9°N、41.8°N、42°N、43.4°N、43.9°N、44.2°N這6個區域達到峰值,并在43.9°N達到最高值,在41.92°N、42.6°N附近出現了最低值區。
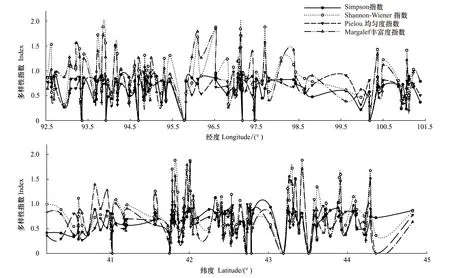
圖4 物種多樣性指數經度和緯度梯度變化Fig.4 Variation of species diversity index along latitude and longitude
整體而言,物種多樣性隨著經度與緯度的變化呈現為持續劇烈波動的復雜變化特征,在經向及緯向均沒有明顯的變化趨勢。結合樣方環境因子的數據分析,發現幾個峰值出現的區域主要為西北部降水較多的天山南北的山前戈壁、中部海拔較高的肅北地區以及額濟納綠洲黑河流經的地區,水熱條件的分配對物種多樣性有著顯著的影響。
3.3.2黑戈壁植物物種多樣性垂直變化分析
黑戈壁地區海拔變化較大,調查的樣方海拔在300—2400m之間。由于海拔導致的溫度和降水的差異,使植被隨著海拔的變化,形成不同的群落類型。由于地表粗糙度過大時,容易導致單優群落的出現,因此隨著海拔的升高,物種多樣性極高值和極低值同時存在,并在幾個海拔區間上出現峰值;物種多樣性在海拔300—1200m之間比較穩定,變化趨勢趨于平坦,但當海拔增加到1400m以上時,物種多樣性隨著海拔的升高有著明顯的增加趨勢,并且與水平方向相比,垂直方向上物種多樣性的變化趨勢要更加明顯。
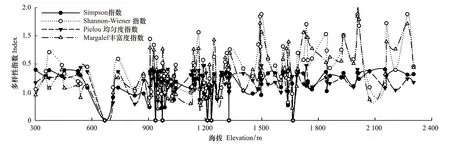
圖5 物種多樣性垂直梯度變化Fig.5 Variation of species diversity index along vertical gradient
3.3.3植物群落物種多樣性空間異質性分析
從表4物種多樣性半變異函數模型及方差分析結果可知,黑戈壁4種多樣性指數除了均勻度指數為球面模型外,其他都更加符合指數模型,半變異函數模型能夠很好地反映物種多樣性指數的空間結構特性。在分析模型中塊金值表征由于取樣誤差和小于取樣尺度上隨機部分引起的空間異質性,基臺值越大,總的空間異質性越高;塊金與基臺的比值表示由系統變量引起的空間相關性,比值小于0.25表示空間相關性強烈,0.25—0.75表示中等的空間相關性,大于0.75表示空間相關性較弱[30];4種多樣性指數比值都在0.25—0.75之間,因此研究區的物種多樣性為中等的空間相關性,物種多樣性呈斑塊化分布。
同時也說明在大于研究尺度,小于變程的范圍內結構性因子所占比例為0.673—0.742,隨機性因子所占比例為0.258—0.327之間,說明由氣候、海拔、地形、土壤理化條件等結構性因子引起的空間異質性占主導地位,而隨機因子,如人為活動引起的空間異質性較弱。
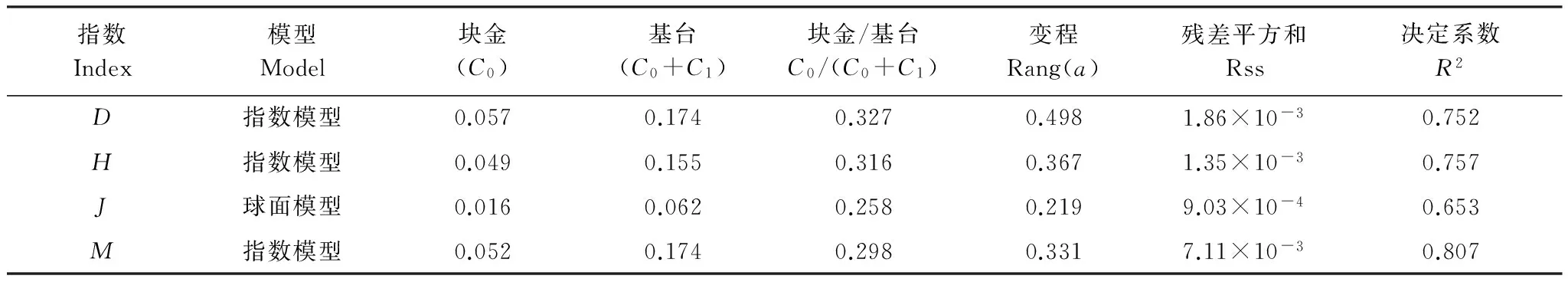
表4 物種多樣性半變異函數模型及其相關參數
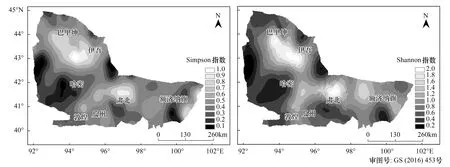
圖6 黑戈壁物種多樣性預測圖Fig.6 Predicting maps of species diversity in Black Gobi
在半方差結構分析和球面模型的基礎上,利用ArcGIS 10.2普通克里金插值法得到黑戈壁地區物種多樣性預測圖6,從中可以明顯的看出黑戈壁地區的植物物種多樣性的空間分布狀況。物種多樣性沒有明顯的經向和緯向變化規律,受大氣活動路徑以及地形對氣候的影響,更多表現為自西北天山北麓向東和向南延伸的過程中不斷減少的趨勢,東天山北麓至北山及馬鬃山迎風坡區域的物種多樣最豐富,敦煌、哈密南部等降水極少地區,生物多樣性水平普遍較低,表現出明顯的斑塊化分布特點。
4結論與討論
4.1黑戈壁植物群落特征及其與環境因子的關系
黑戈壁地區地處我國的溫帶荒漠區,環境惡劣,土壤貧瘠,群落類型貧乏,物種生活型單一,主要以溫帶荒漠灌木和半灌木群落為主,并在部分區域形成溫帶灌叢和草甸。與其他荒漠區比較[11-12],在群落物種組成上,灌木、半灌木、小灌木所占比例更高,境內水土條件更加嚴苛,群落組成更為簡單,優勢群落優勢度更高,群落組成物種通常為2—5個;建群種多為灌木、半灌木植物;與沙漠植被組成相比,則缺乏短命植物。
植被分布格局是不同尺度上包括氣候、土壤、地形在在內多種環境因子共同影響的結果。區域尺度上,地帶性氣候(溫度與降水)是決定植被類型分布及物種生活型的決定性因子,是地帶性植被形成的基礎[31- 32];而局部尺度上,包括微生境及土壤等非地帶性環境因子決定著植被格局[31- 33],地形通過調節局部氣候條件,進而控制地表的水熱等因子,最終對群落的分布格局產生重要影響[34]。本研究結果證實了氣候、土壤、地形是導致群落類型與組成產生差異的主要因素,研究區極端干旱的氣候條件是戈壁荒漠植被形成的基礎,而受到海拔等地形因子的影響,水土資源在局部尺度上的空間異質性是群落空間分異的主因,這與前人[34- 35]的研究結果一致;同時有48.1%的環境變量沒有得到解釋,表明群落分布還受到包括土壤化學性質、生物相互作用因素、干擾因素和隨機因素等的影響[36]。
4.2黑戈壁植物群落物種多樣性空間變化
黑戈壁地區物種多樣性分布主要受到水熱條件的控制;受天山、馬鬃山等山脈的影響使得水熱條件發生再分配,整體上出現了西北—東南走向的水分和熱量的變化梯度[22],物種多樣性沒有明顯的經向和緯向變化規律,更多表現為自西北天山北麓向東和向南延伸的過程中不斷減少的趨勢。我國內陸干旱區物種多樣性多表現為中部隆起型的垂直梯度格局[30],黑戈壁地區物種多樣性格局與其非常相似,在西部地區受到天山影響,海拔落差在2400m以上,而中部地區馬鬃山等海拔也在2000m以上,東部的額濟納地區綠洲為發源于祁連山的黑河下游地區;研究區水熱條件垂直分異明顯,海拔對物種多樣性有著重要的影響,海拔落差越大,這種影響越顯著。
整體上,黑戈壁區不同群落間空間異質性強烈,為中等空間關聯性,地理因素和海拔導致的水熱條件和土壤條件等結構性因子異質性是群落產生空間異質性的主導因素;利用克里金插值得到的物種多樣性預測圖也說明了地理因素對植物物種多樣有著重要影響。黑戈壁物種多樣性的空間變異模式表明植物群落空間異質性強,呈現為斑塊性分布,但總體上受到水熱條件的控制。
4.3黑戈壁植物群落及其物種多樣性維持的一般機理
關于干旱地區植被維持的問題,不同學者觀點也不一致。目前,主要的觀點包括空間異質性影響、空間自組織等,或者兼而有之[31-35,37-40]。Wesche通過對戈壁地區植被分布研究提出[16],水資源的可利用性是決定植被存在與分布的最關鍵因素。地形可能對于水熱條件再分配起到重要的作用,在植被格局形成過程中,植物對有限資源的競爭和利用被認為是最重要的驅動力[41]。而物種在競爭過程中所形成的生長、繁殖以及擴散能力的分化對于物種的共存是非常重要的[40-42]。在黑戈壁地區,植物群落組成主要以灌木和半灌木植物為主,并且密集型的克隆生長比較普遍,植物有性繁殖方式也很特殊甚至特化[12,22]。目前的研究也表明:植物克隆生長和不同繁殖特性對于個體生存以及群落動態有著重要的作用[43]。戈壁植物群落格局及其物種多樣性與水文過程以及戈壁植物特殊的生長與繁殖適應性等因素有關,但戈壁植物群落格局及物種多樣性維持機理與過程具體是怎樣的,這些問題還需要深入研究來解決。
參考文獻(References):
[1]Levine J M, HilleRisLambers J. The importance of niches for the maintenance of species diversity. Nature, 2009, 461(7261): 254- 257.
[2]Thibaut L M, Connolly S R. Understanding diversity-stability relationships: towards a unified model of portfolio effects. Ecology Letters, 2013, 16(2): 140- 150.
[3]Burke A. Classification and ordination of plant communities of the Naukluft Mountains, Namibia. Journal of Vegetation Science, 2001, 12(1): 53- 60.
[4]張雪妮, 呂光輝, 王庭權, 馬玉, 阿布里孜·阿不都熱合, 趙曉英, 郭振潔, 朱修逸. 荒漠區垂直河岸帶植物多樣性格局及其成因. 生態學報, 2015, 35(18): 5966- 5974.
[5]張錦春, 王繼和, 趙明, 劉虎俊, 廖空太, 徐先英. 庫姆塔格沙漠南緣荒漠植物群落多樣性分析. 植物生態學報, 2006, 30(3): 375- 382.
[6]袁蕾, 周華榮, 宗召磊, 葉琴. 烏魯木齊地區典型灌木群落結構特征及其多樣性研究. 西北植物學報, 2014, 34(3): 595- 603.
[7]孟瑩瑩, 周莉, 周旺明, 于大炮, 包也, 王曉雨, 郭焱, 代力民. 長白山風倒區植被恢復26年后物種多樣性變化特征. 生態學報, 2015, 35(1): 142- 149.
[8]Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220- 227.
[9]陳昌篤, 張立運. 中國的極旱荒漠. 干旱區資源與環境, 1987, 1(3/4): 1- 12.
[10]吳正. 中國沙漠及其治理. 北京: 科學出版社, 2009.
[11]陳鵬, 潘曉玲. 河西走廊地區植物的區系特征. 植物研究, 2001, 21(1): 24- 30.
[12]夏延國, 寧宇, 李景文, 李俊清, 馮益民, 吳波, 盧琦. 中國黑戈壁地區植物區系及其物種多樣性研究. 西北植物學報, 2013, 33(9): 1906- 1915.
[13]朱軍濤, 于靜潔, 王平, 王志勇. 額濟納荒漠綠洲植物群落的數量分類及其與地下水環境的關系分析. 植物生態學報, 2011, 35(5): 480- 489.
[14]魚騰飛, 馮起, 司建華, 席海洋, 陳麗娟. 黑河下游額濟納綠洲植物群落物種多樣性的空間異質性. 應用生態學報, 2011, 22(8): 1961- 1966.
[15]Pfeiffer M, Chimedregzen L, Ulykpan K. Community organization and species richness of ants (Hymenoptera/Formicidae) in Mongolia along an ecological gradient from steppe to Gobi desert. Journal of Biogeography, 2003, 30(12): 1921- 1935.
[16]Wesche K, Miehe S, Miehe G. Plant communities of the Gobi Gurvan Sayhan National park (South Gobi Aymak, Mongolia). Candollea, 2005, 60(1): 149- 205.
[17]Von Wehrden H, Wesche K. Relationships between climate, productivity and vegetation in southern Mongolian drylands. Basic and Applied Dryland Research, 2007, 1(2): 100- 120.
[18]Wesche K, von Wehrden H. Surveying Southern Mongolia: application of multivariate classification methods in drylands with low diversity and long floristic gradients. Applied Vegetation Science, 2011, 14(4): 561- 570.
[19]Sasaki T, Okayasu T, Takeuchi K, Jamsran U, Jadambaa S. Patterns of floristic composition under different grazing intensities in Bulgan, South Gobi, Mongolia. Grassland Science, 2005, 51(3): 235- 242.
[20]Okayasu T, Muto M, Jamsran U, Takeuchi K. Spatially heterogeneous impacts on rangeland after social system change in Mongolia. Land Degradation & Development, 2007, 18(5): 555- 566.
[21]Sugimoto N, Hara Y, Yumimoto K, Uno I, Nishikawa M, Dulam J. Dust emission estimated with an assimilated dust transport model using lidar network data and vegetation growth in the Gobi desert in Mongolia. SOLA, 2010, 6: 125- 128.
[22]中國黑戈壁地區生態本底科學考察隊. 中國黑戈壁研究. 北京: 科學出版社, 2014.
[23]吳征鎰. 中國植被. 北京: 科學出版社, 1980.
[24]Jennings M D, Faber-Langendoen D, Loucks O L, Peet R K, Roberts D. Standards for associations and alliances of the U.S. National Vegetation Classification. Ecological Monographs, 2009, 79(2): 173- 199.
[25]張金屯. 數量生態學. 北京: 科學出版社, 2004.
[27]曹靜, 苗艷明, 馮飛, 許強, 張欽弟, 畢潤成. 稀有種不同處理對典范對應分析排序結果影響的比較. 植物生態學報, 2015, 39(2): 167- 175.
[28]ter Braak C J F,milauer P. CANOCO Reference Manual and CanoDraw for Windows User′s Guide: Software for Canonical Community Ordination (Ver. 4.5). New York: Microcomputer Power, Ithaca, 2002.
[29]Borcard D, Legendre P, Drapeau P. Partialling out the spatial component of ecological variation. Ecology, 1992, 73(3): 1045- 1055.
[30]馬斌, 周志宇, 張莉麗, 高文星, 陳善科, 張寶林. 阿拉善左旗植物物種多樣性空間分布特征. 生態學報, 2008, 28(12): 6099- 6106.
[31]Van Couwenberghe R, Collet C, Lacombe E, Pierrat J C, Gégout J C. Gap partitioning among temperate tree species across a regional soil gradient in windstorm-disturbed forests. Forest Ecology and Management, 2010, 260(1): 146- 154.
[32]Siefert A, Ravenscroft C, Althoff D, Alvarez-Yépiz J C, Carter B E, Glennon K L, Heberling J M, Jo I S, Pontes A, Sauer A, Willis A, Fridley J D. Scale dependence of vegetation-environment relationships: a meta‐analysis of multivariate data. Journal of Vegetation Science, 2012, 23(5): 942- 951.
[33]李國慶, 王孝安, 郭華, 朱志紅. 陜西子午嶺生態因素對植物群落的影響. 生態學報, 2008, 28(6): 2463- 2471.
[34]余敏, 周志勇, 康峰峰, 歐陽帥, 米湘成, 孫建新. 山西靈空山小蛇溝林下草本層植物群落梯度分析及環境解釋. 植物生態學報, 2013, 37(5): 373- 383.
[35]沈澤昊, 張新時, 金義興. 地形對亞熱帶山地景觀尺度植被格局影響的梯度分析. 植物生態學報, 2000, 24(4): 430- 435.
[36]趙從舉, 康慕誼, 雷加強. 準噶爾盆地典型地段植物群落及其與環境因子的關系. 生態學報, 2011, 31(10): 2669- 2677.
[37]Couteron P, Lejeune O. Periodic spotted patterns in semi-arid vegetation explained by a propagation inhibition model. Journal of Ecology, 2001, 89(4): 618- 628.
[38]Van de Koppel J, Rietkerk M, Van Langevelde F, Kumar L, Klausmeier C A, Fryxell J M, Hearne J W, van Andel J, de Ridder N, Skidmore A, Stroosnijder L, Prins H T P. Spatial heterogeneity and irreversible vegetation change in semi-arid Grazing systems. The American Naturalist, 2002, 159(2): 209- 218.
[39]Rietkerk M, Ouedraogo T, Kumar L, Sanou S, van Langevelde F, Kiema A, van de Koppel J, van Andel J, Hearne J, Skidmore A, de Ridder N, Stroosnijder L, Prins H H T. Fine-scale spatial distribution of plants and resources on a sandy soil in the Sahel. Plant and Soil, 2002, 239(1): 69- 77.
[40]HilleRisLambers R, Rietkerk M, Van Den Bosch F, Prins H H T, de Kroon H. Vegetation pattern formation in semi-arid grazing systems. Ecology, 2001, 82(1): 50- 61.
[41]Ryabov A B, Blasius B. A graphical theory of competition on spatial resource gradients. Ecology Letters, 2011, 14(3): 220- 228.
[42]RietKerk M, Boerlijst M C, van Langevelde F, HilleRisLambers R, Van De Koppel J, Kumar L, Prins H H T, De Roos A M. Self-Organization of vegetation in arid ecosystems. The American Naturalist, 2002, 160(4): 524- 530.
[43]Charpentier A. Consequences of clonal growth for plant mating. Evolutionary Ecology, 2002, 15(4/6): 521- 530.
基金項目:林業行業公益項目(201404304);中國林業科學研究院重大項目 (CAFYBB2011002);國家自然科學基金資助項目(31570610)
收稿日期:2015- 06- 10;
修訂日期:2016- 01- 25
*通訊作者
Corresponding author.E-mail: lijingwen@bjfu.edu.cn
DOI:10.5846/stxb201506101176
Plant distribution patterns and the factors influencing plant diversity in the Black Gobi Desert of China
WANG Jianming1, DONG Fangyu1, BahaiNasina1,3, LI Jingwen1,*,LI Junqing1,FENG Yiming2, LU Qi2
1TheCollegeofBeijingForestryUniversity,Beijing100083,China2InstituteofDesertificationStudies,ChineseAcademyofForestry,Beijing100091,China3XinjiangKeyForestryEngineeringQualityManagementStation,Urumchi830000,China
Abstract:The Black Gobi Desert covers a large area in the northwest of China, with a total area of about 200,000 km2. The general climate of the Black Gobi can be characterized as having a high evaporative demand, with variable but low precipitation, making it the most arid region of the country′s dryland areas. Nevertheless, despite being considered the most dry and bare ecosystem in the world, it is affluent in natural resources. Because of its harsh environment and poor accessibility, systematic studies of plant diversity in the Black Gobi Desert are lacking. Understanding the underlying causes of geographic plant diversity gradients in arid and semiarid regions is important for the conservation and sustainable use of biodiversity there. However, the underlying mechanisms behind the Black Gobi Desert′s plant diversity patterns are still elusive. Therefore, the aim of our study was to analyze the plant community composition, as well as the species diversity of the Black Gobi ecosystem. We sampled 174 plots across the Black Gobi to demonstrate the vegetation pattern and its relationship with the environmental factors. We observed a trend of the plant communities becoming poorer and simpler in composition while vegetation cover decreases. Moreover, the overall structure and composition of the plant communities were simple, the majority of plants having an importance value lower than 0.1. The plant communities consisted mainly of Haloxylon ammodendron, Reaumuria songarica, Nitraria tangutorum, and 10 other species. Species diversity showed non-significant latitudinal and longitudinal trends, but was significantly influenced by climate, elevation, and topography. In addition, structural factors were most important in determining the spatial variation of species diversity. The result of a detrended cross-correlations analysis (DCCA) indicated that climate, elevation, topography, soil texture, and surface cover had a significant effect on community composition and plant distribution. Overall, the vegetation spanned a gradient in elevation,soil texture and water availability. Plant species are limited by water availability, and it should be noted that plant community composition was strongly related to changes in elevation, which influence soil texture and water availability. In sum, the spatial heterogeneity of the soil texture and water availability were the main factors to determine plant distribution and species diversity.
Key Words:Black Gobi; plant communities; species diversity; plant distribution; spatial heterogeneity
Wang J M, Dong F Y, BahaiNasina, Li J W,Li J Q,Feng Y M, Lu Q.Plant distribution patterns and the factors influencing plant diversity in the Black Gobi Desert of China.Acta Ecologica Sinica,2016,36(12):3488- 3498.
