p21基因在野生型斑馬魚胚胎發(fā)育過程中的表達*
2016-08-01 09:47:44姬牧遠吳西軍周艷華舒莉萍1何志旭1
貴州醫(yī)科大學(xué)學(xué)報 2016年7期
宋 錦, 姬牧遠, 吳西軍, 周艷華, 舒莉萍1,,4*** , 何志旭1,,3***
(1.貴州醫(yī)科大學(xué) 免疫學(xué)教研室, 貴州 貴陽 550004; 2.貴州醫(yī)科大學(xué) 干細胞與組織工程實驗中心, 貴州 貴陽 550004; 3.貴州醫(yī)科大學(xué)附院 兒科學(xué)教研室, 貴州 貴陽 550004; 4.貴州醫(yī)科大學(xué) 實驗動物中心, 貴州 貴陽 550004)
?
p21基因在野生型斑馬魚胚胎發(fā)育過程中的表達*
宋錦1,2**, 姬牧遠2,3, 吳西軍2, 周艷華2, 舒莉萍1,2,4***, 何志旭1,2,3***
(1.貴州醫(yī)科大學(xué) 免疫學(xué)教研室, 貴州 貴陽550004; 2.貴州醫(yī)科大學(xué) 干細胞與組織工程實驗中心, 貴州 貴陽550004; 3.貴州醫(yī)科大學(xué)附院 兒科學(xué)教研室, 貴州 貴陽550004; 4.貴州醫(yī)科大學(xué) 實驗動物中心, 貴州 貴陽550004)
[摘要]目的: 探究p21基因在斑馬魚早期發(fā)育過程中的表達。方法: Trizol法提取野生型斑馬魚胚胎總RNA,RT-PCR法擴增p21基因片段,與pCS2+載體重組構(gòu)建p21-pCS2+重組質(zhì)粒,以T3 RNA 聚合酶制備地高辛標記的反義mRNA 探針,通過斑馬魚全胚胎原位雜交檢測p21 基因在野生型斑馬魚胚胎早期發(fā)育過程中的表達。結(jié)果: 成功構(gòu)建了p21-pCS2+重組質(zhì)粒,并制備反義mRNA 探針;原位雜交結(jié)果顯示野生型斑馬魚0.5~18 hpf的胚胎均未發(fā)現(xiàn)p21 基因陽性雜交信號,24~36 hpf 胚胎可在脊索部位觀察到陽性信號,48~72 hpf 胚胎可在頭部胸腺附近觀察到陽性信號,表達量均較低。結(jié)論: 成功制備了p21-pCS2+重組質(zhì)粒及斑馬魚p21 基因反義mRNA 探針,p21 基因在野生型斑馬魚胚胎不同發(fā)育時相中表達水平不同。
[關(guān)鍵詞]p21; 斑馬魚; 胚胎; 發(fā)育; 探針; 質(zhì)粒
p21 基因表達產(chǎn)物是細胞周期的一種抑制因子,是細胞周期蛋白依賴性激酶抑制因子CKI 中細胞因子誘導(dǎo)的蛋白/激酶相互作用蛋白Cip/Kip (cytokine-inducible protein/kinase inhibition protein,Cip/Kip) 家族的一員。當在細胞接受到接觸抑制、DNA損傷、低氧及某些細胞因子等信號后產(chǎn)生P21基因表達產(chǎn)物,參與細胞的生長、分化、衰老及死亡過程,同時P21基因表達產(chǎn)物又與腫瘤發(fā)生密切相關(guān),在細胞的生理、病理過程中發(fā)揮重要作用[1-3]。斑馬魚作為一種新興的模式生物,具有成魚體積小,適宜在較小空間里大量養(yǎng)殖,生命力強,成熟周期短,繁殖能力強,在體外受精發(fā)育,胚胎透明,便于在四維領(lǐng)域即其前后、上下、左右及時間上進行觀察,可進行高通量基因篩查,全基因組測序工作已完成等優(yōu)點。同時斑馬魚與哺乳動物在分子途徑調(diào)控造血上高度保守,提供了幾乎與哺乳動物和其他高級脊椎動物一致的造血系統(tǒng)[4]。因此,利用斑馬魚作為模式生物來研究造血系統(tǒng)功能和疾病有著廣闊的前景。本實驗以斑馬魚為對象,構(gòu)建p21-pCS2+重組質(zhì)粒,制備p21反義mRNA探針,以全胚胎原位雜交法觀察斑馬魚胚胎發(fā)育中p21的表達情況,為下一步以斑馬魚作為模型研究p21奠定基礎(chǔ)。
1材料和方法
1.1材料
Tuebingen品系斑馬魚參照Westerfield[5]方法養(yǎng)殖在28.5℃的循環(huán)水系統(tǒng)中。根據(jù)Kimmel等[6]文獻中提供的斑馬魚生長發(fā)育圖譜,區(qū)分其發(fā)育階段,收集時相為受精后0.75 h(0.75 hours post-fertilization,hpf)、3.7 hpf、6 hpf、12 hpf、18 hpf、24 hpf、30 hpf、36 hpf、48 hpf以及72 hpf。
1.2方法
1.2.1獲取p21 cDNA收集野生型斑馬魚胚胎,每個時相10 枚,用Trizol方法提取總RNA,使用Transgene公司TransScript First-Strand cDNA Synthesis SuperMix試劑盒反轉(zhuǎn)錄獲取p21 cDNA,操作按說明書。
1.2.2體外克隆p21根據(jù)NCBI數(shù)據(jù)提供的斑馬魚p21基因全長cDNA,設(shè)計引物序列p21-F,5′-CC ATCGAT GG CAGCTCTTGCAGAAGCTC-3′;p21-R,5′-GCTCTAGACTGGCCGG ATTTGCGCGG-3′;引物序列包含ClaⅠ、XbaⅠ酶切位點。利用KOD-plus擴增p21基因,為保證PCR產(chǎn)物的特異性,將上述體系以降落PCR法完成體外擴增。PCR程序如下:94 ℃ 8 min,94 ℃ 40 s,68 ℃ 30 s,72 ℃ 60 s,以后每2個循環(huán)降低退火溫度1 ℃至退火溫度降至60 ℃,94 ℃ 40 s,60 ℃ 30 s,72 ℃ 60 s(20個循環(huán)),72 ℃ 10 min。
1.2.3構(gòu)建p21-pCS2+重組質(zhì)粒將pCS2+質(zhì)粒及p21 基因的PCR 產(chǎn)物分別用ClaⅠ、XbaⅠ雙酶切,酶切后進行電泳分析,回收目的條帶,將回收的經(jīng)酶切的pCS2+質(zhì)粒及p21 基因PCR 產(chǎn)物進行連接。將連接產(chǎn)物轉(zhuǎn)入EcoliDH5α 感受態(tài)菌,通過AMP 抗性篩選,挑選陽性克隆,擴增后用Axygen 質(zhì)粒小抽試劑盒抽提p21-pCS2+重組質(zhì)粒。
1.2.4p21-pCS2+重組質(zhì)粒鑒定 以ClaⅠ及XbaⅠ雙酶切p21-pCS2+重組質(zhì)粒,通過瓊脂糖電泳判斷酶切產(chǎn)物大小,鑒定是否有插入序列DNA 及pCS2+質(zhì)粒DNA。并將p21-pCS2+重組質(zhì)粒委托北京諾賽基因公司進行測序。
1.2.5制備p21 基因反義mRNA 探針將p21-pCS2+重組質(zhì)粒用ClaⅠ單酶切進行質(zhì)粒DNA 線性化,將線性化p21-pCS2+重組質(zhì)粒加入地高辛標記的寡核苷酸,利用T3RNA 聚合酶完成體外轉(zhuǎn)錄,得到地高辛標記的p21 基因反義mRNA 探針,置于-70 ℃保存?zhèn)溆谩?/p>
1.2.6斑馬魚p21全胚胎原位雜交分別收集0.75~72 hpf時相的野生型斑馬魚胚胎,去除卵膜,4%多聚甲醛(PFA)固定24 h,用1×PBST 洗3次,然后依次用25%甲醇(PBS)、50%甲醇(PBS)、75%甲醇(PBS)和100%甲醇洗滌,-20 ℃保存過夜。體視顯微鏡下挑選已脫水的完整胚胎,每個時相挑選20~30 枚,依次用75 %甲醇(PBST)、50%甲醇(PBST)、25%甲醇(PBST) 溶液洗滌1次。24 hpf之后的斑馬魚胚胎需用蛋白酶K 處理,以利于探針滲透。將胚胎移入68 ℃雜交爐,加入預(yù)熱的預(yù)雜交液HYB(-), 15 min。吸出HYB(-),加入HYB(+),68℃ 1 h;再加入地高辛標記的p21反義mRNA 探針,68 ℃過夜。在雜交爐中,用50%甲酰胺(2×SSCT)1mL洗滌兩次,每次68 ℃ 30 min。再用2×SSCT 1 mL洗滌1次,68 ℃ 15 min。最后用0.2×SSCT 1 mL洗滌兩次,每次68 ℃ 30 min。將胚胎移至十二孔板中,用MABT 2.5 mL洗3次,每次室溫輕搖5 min。加入阻滯液2.5 mL于室溫下輕搖1 h。加入抗地高辛抗體至終濃度為1∶5 000,4 ℃輕搖過夜。吸去抗體,用阻滯液 lml洗滌1次,室溫輕搖30 min,再用MABT 2 mL洗滌兩次,室溫輕搖1 h及30 min,用染色緩沖液洗滌3次。加入NBT/BCIP染液染色,并在體視顯微鏡下觀察,直至染色成功。將胚胎用1×PBST洗滌后,用4%的多聚甲醛固定,4 ℃保存,在體視顯微鏡(Nikon-SMZ150)下拍照。
2結(jié)果
2.1重組質(zhì)粒p21-pCS2+及反義p21 基因反義mRNA 探針
PCR 產(chǎn)物經(jīng)凝膠電泳分析顯示500 bp 附近可見一特異擴增帶,大小與預(yù)期525 bp 結(jié)果相符(圖1);重組質(zhì)粒p21-pCS2+電泳結(jié)果顯示,在4 000 bp 附近可見兩條條帶,525 bp 附近可見1條條帶(圖2),提示重組質(zhì)粒構(gòu)建成功。p21 基因反義mRNA 探針電泳結(jié)果顯示,在500 bp位置有明顯條帶,重組質(zhì)粒測序結(jié)果通過Genbank 比對顯示與所需p21 基因片段完全一致,提示p21 基因反義mRNA 探針制備成功(圖3、圖4)。2.2p21 基因在野生型斑馬魚全胚胎原位雜交結(jié)果
0.75~18 hpf 胚胎未見p21 基因陽性雜交信號(圖5 A-F),24~36 hpf 胚胎部分脊索可觀察到p21 基因陽性雜交信號(圖5 G-I紅色箭頭所示),但表達量并不高;48~72 hpf 胚胎在頭部可見陽性信號(圖5 J-K綠色箭頭所示),兩側(cè)對稱出現(xiàn)。0.75~72 hpf在斑馬魚胚胎造血區(qū)如ICM 區(qū)、AGM 區(qū)未觀察到陽性雜交信號,提示野生型斑馬魚造血系統(tǒng)無p21 基因表達或p21 基因表達極低(圖5)。

注:1為DNA marker Ⅲ,2為PCR產(chǎn)物,3為GAPDH,4為空白對照圖1 p21 基因PCR 擴增產(chǎn)物電泳Fig.1 Electrophoresis of PCR products of p21

注:1為DNA marker Ⅲ,2為Enzyme digested p21-pCS2+,3為Linearity p21-pCS2+,4為Linearity plasmid pCS2+,5為空白對照圖2 p21-pCS2+限制性內(nèi)切酶酶切鑒定Fig.2 The restriction enzymes digest p21-pCS2+with ClaⅠand XbaⅠ

注:1為DNA marker Ⅳ,2和3為反義p21 mRNA探針,4為空白對照圖3 p21 基因反義mRNA 探針電泳Fig.3 Electrophoresis of the anti-sense p21 mRNA probe
3討論
p21基因最初是通過酵母雙雜交及消減雜交等方法得到,主要參與DNA損傷的細胞應(yīng)答反應(yīng)[7-8]。Harpeer等在研究CDK2的調(diào)節(jié)蛋白時發(fā)現(xiàn),p21作為CDK2及其他CDK的抑制物,在細胞分裂過程中起著關(guān)卡作用[9]。Vogelstein等在尋找受p53轉(zhuǎn)錄調(diào)控的基因中發(fā)現(xiàn)了p21基因,將其稱為WAFl(wild-type p53-activated fragment1),將其導(dǎo)入腫瘤細胞時,可發(fā)現(xiàn)其與p53有同樣的抑瘤效應(yīng)[10]。目前,有關(guān)p21基因的結(jié)構(gòu)功能的認識逐漸增多,許多研究表明其與細胞終末分化、損傷、衰老有著重要的功能意義,并參與癌癥的發(fā)生[11-13]。因此對p21基因相關(guān)功能的研究可能成為治療癌癥的新靶點。
p21基因最核心的功能是其參與調(diào)節(jié)細胞周期的功能,可影響細胞的自我更新、分裂、分化及凋亡等。當DNA損傷時,一系列蛋白激酶被激活、核苷酸的耗竭、氧供不足和過剩等可激活p53,p21基因編碼區(qū)上游2.4 kb和約8 kb處有2個p53共有的結(jié)合區(qū),因而p53基因激活會導(dǎo)致其靶基因p21基因的迅速轉(zhuǎn)錄。轉(zhuǎn)錄出的p21蛋白可與CDK2 等因子結(jié)合,抑制其活性,使受到損傷的細胞停滯于G1期,不能進一步進行DNA復(fù)制和有絲分裂,從而使受損的細胞有充分的時間修復(fù), 不能修復(fù)的則發(fā)生凋亡[10,14]。研究發(fā)現(xiàn),在血清供給量不足、接觸性抑制和衰老等條件下,通過腫瘤壞死因子、組織型纖溶酶原激活劑等因素的作用,p21基因可以快速發(fā)生反應(yīng),使受損的細胞發(fā)生細胞周期阻滯,在正常組織細胞的發(fā)育、分化中非依賴p53途徑的p21基因的調(diào)節(jié)同樣存在[15]。p21基因可以與增殖細胞核抗原(proliferating cell nuclear antigen ,PCNA)相互作用,在S期發(fā)生DNA損傷,通過p21 C端競爭性抑制DNA聚合酶和PCNA的結(jié)合,阻斷PCNA活化DNA聚合酶的活性從而抑制DNA的合成,使細胞周期停滯[16]。
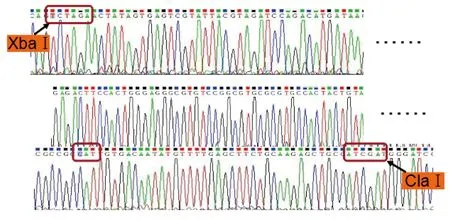
圖4 p21-pCS2+重組質(zhì)粒測序Fig.4 DNA Sequence of the recombinant plasmid of p21-pCS2+
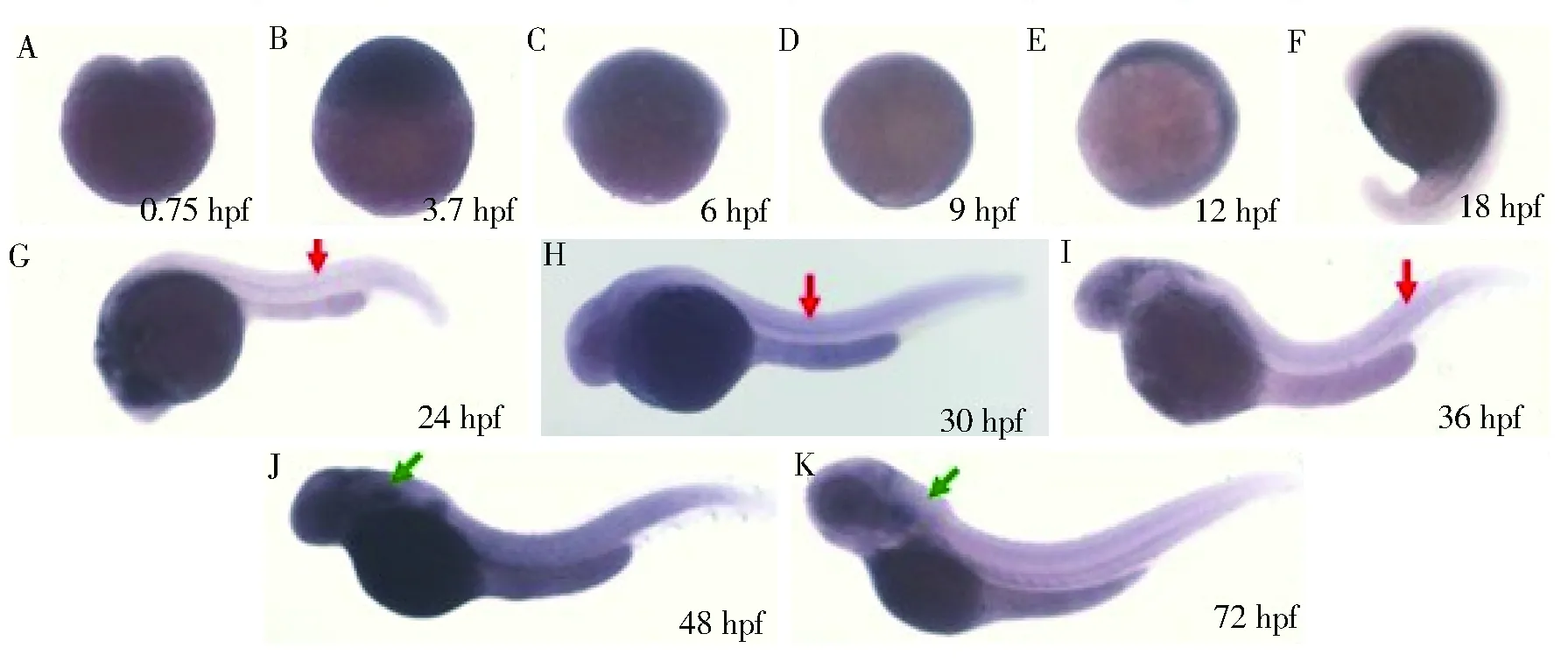
注:紅色箭頭為脊索,綠色箭頭為胸腺圖5 p21基因在野生型斑馬魚胚胎發(fā)育過程中的表達Fig.5 Expression of p21 gene in wild-type zebrafish embryos
目前斑馬魚作為一種模式生物已廣泛應(yīng)用于各種疾病的研究。對致癌化合物引起的斑馬魚腫瘤的研究顯示,其在基因組學(xué)及組織病理等方面與人類腫瘤高度相似[17-18]。2003年,Langenau等[19]創(chuàng)建了首個斑馬魚白血病模型,并且將該白血病模型斑馬魚的血細胞轉(zhuǎn)入亞致死量射線照射過的野生型斑馬魚,同樣可以引起白血病癥狀,這進一步引起了通過斑馬魚來研究造血系統(tǒng)疾病的熱潮。p21基因是一個高度保守的基因,在氨基酸水平上人的p21基因與斑馬魚有87%的同源性。因此本研究選擇斑馬魚為模式生物,有利于對p21基因的功能的研究,也是國內(nèi)外首次報道p21基因在野生斑馬魚中的表達情況。
在本研究中,通過對野生型斑馬魚胚胎進行全胚胎原位雜交,在胚胎發(fā)育早期(0.75~18 hpf)未觀察到p21基因表達,p21基因為細胞周期中與G1/S檢查點相關(guān)的關(guān)鍵調(diào)節(jié)因子,與細胞增殖的機制相關(guān);當有錯誤的DNA復(fù)制時,p21基因表達增加,限制細胞周期的進程,使其進行DNA修復(fù)后再繼續(xù)分裂。故對此推測在細胞增殖受阻的部位p21基因呈高表達,而在細胞分裂、分化旺盛的部位其表達降低,早期斑馬魚胚胎生長發(fā)育旺盛,細胞大量分化,所以p21基因表達量低,通過原位雜交方法不能觀測到其表達,但具體機制有待于進一步的研究。24~36 hpf胚胎可在脊椎部位觀察到陽性信號,表明p21基因與斑馬魚早期胚胎發(fā)育過程中可能與神經(jīng)系統(tǒng)發(fā)育相關(guān)。48~72 hpf胚胎可在頭部觀察到陽性信號,兩側(cè)對稱出現(xiàn),考慮該部位為胸腺。下一步可通過胸腺發(fā)育相關(guān)的基因進行進一步認證p21是否在斑馬魚胸腺內(nèi)表達,利用斑馬魚優(yōu)勢模式生物,可能揭示T細胞的發(fā)生和分化與p21的關(guān)系。
4參考文獻
[1] Arthur LM, Heber-Katz E. The role ofp21 in regulating mammalian regeneration[J]. Stem Cell Res Ther, 2011(3):30.
[2] Ronald Van OS,Leonie M, Kamminga, et al. A Limited Role forp21Cip1/Waf1 in Maintain- ing Normal Hematopoietic Stem Cell Functioning[J]. Stem Cells, 2007(4):836-843.
[3] Andrea V, Francesca DF, Annette O,et.al.Cell-cycle restriction limits DNA damage and maintains self-renewal of leukaemia stem cells[J].Nature, 2009(7225):51-56.
[4] Mushtaq MY, Verpoorte R,Kim HK. Zebrafish as a model for systems biology[J]. Biotechnol Genet Eng Rev, 2013(29):187-205.
[5] Runft DL, Mitchell KC, Abuaita BH, et al. Zebrafish as a natural host model for Vibrio cholerae colonization and transmission[J]. Appl Environ Microbiol, 2014(5):1710-1717.
[6] Shinya M,Sakai N. Generation of highly homogeneous strains of zebrafish through full sib-pair mating[J]. G3 (Bethesda), 2011(5):377-386.
[7] Hoeferlin LA, Oleinik NV, Krupenko NI, et al.Activation ofp21-Dependent G1/G2 arrest in the absence of DNA damage as an antiapoptotic response to metabolic stress[J]. Genes Cancer, 2011(9):889-899.
[8] Armstrong MJ, Stang MT, Liu Y, et al. Interferon regulatory factor 1 (IRF-1) inducesp21(WAF1/CIP1) dependent cell cycle arrest andp21(WAF1/CIP1) independent modulation of survivin in cancer cells[J]. CancerLett, 2012(1):56-65.
[9] Sun X, Liu B, Wang J, et al. Inhibition ofp21-activated kinase 4 expression suppresses the proliferation of Hep-2 laryngeal carcinoma cells via activation of the ATM/Chk1/2/p53 pathway[J]. Int J Oncol, 2013(2):683-689.
[10]Russo A, Esposito D, Catillo M, et al. Human rpL3 induces G(1)/S arrest or apoptosis by modulatingp21 (waf1/cip1) levels in a p53-independent manner[J]. Cell Cycl, 2013 (1):76-87.
[11]Graham K, Moran-Jones K, Sansom, et al. FAK deletion promotes p53-mediated induction ofp21, DNA-damage responses and radio-resistance in advanced squamous cancer cells[J]. PLoS One, 2011(12):e27806.
[12]Chiang YT,Jin T.p21-Activated protein kinases and their emerging roles in glucose homeostasis[J].Am J Physiol Endocrinol Metab, 2014(7):E707-722.
[13]Hu Z, ZhangD, Hao J,et al. Induction of DNA damage andp21-dependent senescence by Riccardin D is a novel mechanism contributing to its growth suppression in prostate cancer cells in vitro and in vivo[J]. Cancer Chemother Pharmacol, 2014(2):397-407.
[14]Tarakji B. Baroudi K, Hanouneh S, et al. Expression ofp21 is dependent on or independent of p53 in carcinoma ex pleomorphic adenoma (undifferentiated and adenocarcinoma types)[J]. Pol J Pathol, 2012(4):286-291.
[15]Zhang Y,Yan B. Cell cycle regulation by carboxylated multiwalled carbon nanotubes through p53-independent induction ofp21 under the control of the BMP signaling pathway[J]. Chem Res Toxicol, 2012(6):1212-1221.
[16]Zhang L, Mei Y, Fu NY, et al. TRIM39 regulates cell cycle progression and DNA damage responses via stabilizingp21[J]. Proc Natl Acad Sci USA, 2012(51):20937-20942.
[17]Delvecchio C, Tiefenbach J, Krause HM.The zebrafish: a powerful platform for in vivo, HTS drug discovery[J].Assay Drug Dev Technol, 2011(4):354-361.
[18]Stern HM,Zon LI.Cancer genetics and drug discovery in the zebrafish [J].Nat Rev Cancer, 2013(7):533-539.
[19]Langenau DM,Traver D,Ferrondo AA,et al.Myc-induced T cell leukemia in transgenic zebrafish [J].Science, 2003(5608):887-890.
(2016-05-10收稿,2016-06-21修回)
中文編輯: 文箐潁; 英文編輯: 趙毅
*[基金項目]國家自然科學(xué)基金項目(30960412); 貴州省科技基礎(chǔ)條件平臺項目[(2009)4005]; 貴州省優(yōu)秀科技教育人才省長專項資金(S2008-4)
[中圖分類號]R394.3
[文獻標識碼]A
[文章編號]1000-2707(2016)07-0765-05
DOI:10.19367/j.cnki.1000-2707.2016.07.005
The Expression ofp21 in Development of Wild Type Zebrafish Embryo
SONG Jin1,2, JI Muyuan2,3, WU Xijun2, ZHOU Yanhua2, SHU Liping1,2,4, HE Zhixu1,2,3
(1.DepartmentofImmunology,GuizhouMedicalUniversity,Guiyang550004,Guizhou,China; 2.TheCenterofStemCellandTissueEngineeringResearch,GuizhouMedicalUniversity,Guiyang550004,Guizhou,China; 3.DepartmentofPaediatrics,AffiliatedHospitalofGuizhouMedicalUniversity,Guiyang550004,Guizhou,China; 4.LaboratoryAnimalCenter,GuizhouMedicalUniversity,Guiyang550004,Guizhou,China)
[Abstract]Objective: To explore the expression of p21 in zebrafish. Methods: Extract 0.75 hpf~120 hpf wild-type Zebrafish embryos in multiple phase and total RNA by Trizol, amplificating p21 gene by RT-PCR, and constructing p21-pCS2+plasmid. Preparing antisense mRNA probe with T3 RNA polymerase, using zebrafish whole-mount in situ hybridization to examine the p21 expression pattern in early embryo development. Results: Successfully constructed p21-pCS2+plasmid, preparing antisense mRNA probe; within situ hybridization, there is no expression of p21 in wild-type zebrafish at 0.5 hpf~18 hpf; positive signal in notochord was found in 24 hpf~36 hpf of the wild-type Zebrafish embryos, and positive signal could be found near head and thymus, but indicating low expression. Conclusion: In this research, it has been successfully constructed p21-pCS2+plasmid, antisense mRNA probe preparation, and have a preliminary observation in the expression of p21 gene in Zebrafish embryos in different periods.
[Key words]p21; zebrafish;embryo;development; probe; plasmid
**貴州醫(yī)科大學(xué)2013級碩士研究生
***通信作者 E-mail:gyslp456@gmc.edu.cn; hzx@gmc.edu.cn
網(wǎng)絡(luò)出版時間:2016-07-17網(wǎng)絡(luò)出版地址:http://www.cnki.net/kcms/detail/52.5012.R.20160717.1318.018.html
