極端高溫對西花薊馬存活、繁殖特性及體內海藻糖、山梨醇含量的影響
2016-07-18 09:32:20李洪剛萬方浩鄭長英
中國農業科學 2016年12期
姜 姍,李 帥,張 彬,李洪剛,2,萬方浩,3,鄭長英
(1青島農業大學農學與植物保護學院山東省重點昆蟲生態學實驗室,山東青島 266109;2山東省植物保護總站,濟南 250100;3中國農業科學院植物保護研究所植物病蟲害生物學國家重點實驗室,北京 100193)
?
極端高溫對西花薊馬存活、繁殖特性及體內海藻糖、山梨醇含量的影響
姜姍1,李帥1,張彬1,李洪剛1,2,萬方浩1,3,鄭長英1
(1青島農業大學農學與植物保護學院山東省重點昆蟲生態學實驗室,山東青島 266109;2山東省植物保護總站,濟南 250100;3中國農業科學院植物保護研究所植物病蟲害生物學國家重點實驗室,北京 100193)
摘要:【目的】西花薊馬(Frankliniella occidentalis)是中國的重要外來入侵和檢疫性害蟲, 對各地的蔬菜和花卉造成了巨大的經濟損失。明確極端高溫(45℃)對西花薊馬存活率、繁殖力以及體內海藻糖、山梨醇含量的影響,為西花薊馬的防治提供理論依據。【方法】設置45℃條件下高溫熱激西花薊馬當代成蟲、2齡若蟲2 h,經過24 h變溫模式22℃(4 h)-25℃(8 h)-28℃(4 h)-25℃(8 h)培養后再進行一次45℃ 2 h的熱激處理。研究熱激處理后其親代、F1代和F2代的種群參數(親代各蟲態存活率、雌成蟲壽命,F1和F2代種群數量、存活率、性比)以及2齡若蟲和成蟲體內海藻糖和山梨醇含量的變化。【結果】45℃ 2 h高溫熱激兩次后,西花薊馬各個蟲態的存活率均小于50%,且預蛹和蛹的存活率為0,成蟲的存活率最高為41.38%,顯著高于其他蟲態(1齡若蟲存活率為5%,2齡若蟲存活率為21.36%)(P<0.05);西花薊馬2齡若蟲和成蟲在45℃高溫條件下熱激2 h兩次后,其親代雌成蟲壽命和繁殖力均明顯降低,且F1代也受到較大影響, F1代種群數量、存活率、性比(♀∶♂)顯著降低,但F2代種群有所恢復,另外熱激處理西花薊馬成蟲的雌成蟲壽命及繁殖力在親代、F1代中都明顯高于熱激處理2齡若蟲(P<0.05);西花薊馬親代成蟲和2齡若蟲在45℃熱激2 h兩次后其體內的海藻糖含量明顯降低,并且影響到F1和F2代種群體內的海藻糖含量,另外熱激處理親代成蟲其體內海藻糖含量低于熱激處理親代2齡若蟲的海藻糖含量, 且F1和F2代與親代趨勢相同(P<0.05);西花薊馬親代成蟲和2齡若蟲在45℃熱激2 h兩次后其體內的山梨醇含量明顯升高,熱激親代成蟲的山梨醇含量高于熱激2齡若蟲體內的山梨醇含量,且這種熱激處理能夠影響到后代種群體內山梨醇的含量,F1和F2代體內山梨醇含量的變化趨勢與親代相同(P <0.05)。【結論】45℃極端高溫處理不僅對西花薊馬親代的生長發育、繁殖和生理生化產生了明顯的影響,而且這種影響延續到了子代(F1、F2代)的不同發育階段。同時,西花薊馬對極端高溫的應對與其體內的海藻糖、山梨醇的含量具有一定的相關性。
關鍵詞:極端高溫;西花薊馬;繁殖力;海藻糖;山梨醇
聯系方式:姜姍,E-mail:906342335@qq.com。通信作者鄭長英,E-mail:zhengcy67@qau.edu.cn
0 引言
【研究意義】溫度是影響昆蟲分布、存活、種群動態、行為特征以及適合度的一個重要的非生物因子[1],昆蟲的正常生長發育需要在特定的溫度范圍才能進行,溫度過高或過低都會對昆蟲的生長發育產生不利影響,在田間或溫室中,昆蟲會經常遭受高溫脅迫,有時甚至超過其生理上限造成死亡[2],因此高溫悶棚方法已用于控制一些溫室害蟲[3-4];然而在低于致死高溫的一定溫度范圍內,高溫會造成昆蟲器官的熱損傷,從而影響到存活率及健康狀況,尤其是導致其生殖適應性降低,從而影響到下一代的種群規模[5]。研究亞致死高溫對昆蟲存活、繁殖以及生理上的影響,對研究昆蟲的種群適合度和種群動態具有一定的指導意義。【前人研究進展】昆蟲對高溫敏感主要是由于其弱小的體型和變溫生理特性造成的[6]。在生物學方面,高溫對昆蟲的影響主要包括降低存活率、繁殖力、致使昆蟲發育延遲或體型畸形等;而在生理上可致蛋白質變性、改變細胞膜通透性、酶的結構以及水分喪失等[7]。海藻糖是生物體內一種重要的雙糖。作為昆蟲體內重要血糖,海藻糖存在于昆蟲幾乎所有的組織和器官中。研究發現海藻糖可在高溫、高寒、高滲透壓及干燥失水等惡劣環境條件下在細胞表面形成獨特的保護膜,有效地保護蛋白質分子使之不變性失活,從而維持生命體的生命過程和生物特征[8-9]。在高溫條件下,海藻糖的表現尤為明顯,并且可能與耐熱性存在某種聯系[10]。另外,生物體內多羥基化合物的積累對其抵抗極端溫度的脅迫也十分重要。例如,研究發現山梨醇在蚜蟲和粉虱耐高溫方面起了非常重要的作用[11-12];高溫刺激了蚜蟲和粉虱取食量的明顯增加,可以使食物中獲取的糖分轉變成山梨醇來提高耐熱性[13]。【本研究切入點】西花薊馬(Frankliniella occidentalis)是世界范圍分布的危險性入侵害蟲[14]。具有個體小、善于隱藏、繁殖能力強、適生區廣泛以及抗藥性不斷增強等特點,防治困難[15]。高溫悶棚是防治西花薊馬的一種安全、有效,且對環境無污染的物理防治措施,已用于溫室中防治害蟲[16],但其機制還不明確。探討西花薊馬對高溫響應的生理機制有利于對其進行科學防治,前期研究發現在45℃熱激處理一次西花薊馬親代2 h,其親代和F1代的繁殖參數變化不明顯。因此,本試驗探究兩次熱激對西花薊馬種群親代和 F1代的影響以及這種熱激是否會對F2代產生影響。【擬解決的關鍵問題】45℃高溫處理西花薊馬不同蟲態,探究高溫連續熱激兩次對其存活率、繁殖力以及海藻糖、山梨醇含量的影響,了解高溫對昆蟲產生的生理損傷,以及這些損傷與昆蟲的生長發育、存活、繁殖等的關系,從生理學、生態學角度為闡明西花薊馬對環境脅迫的適應機制提供依據。
1 材料與方法
試驗于2014—2015年在青島農業大學完成。
1.1 供試材料與儀器
供試西花薊馬采自青島農業大學校園內三葉草(Trifolium repens)上,采回后在室內變溫模式下用紫甘藍(Brassica oleracea)飼養多代后備用。
MGC-250BP-2型光照培養箱(上海恒科學儀器有限公司,溫差:±0.5℃)、酶標儀、超低溫冰箱、Eppendorf冷凍離心機、恒溫磁力攪拌器、Thermo超低溫冰箱。
1.2 試驗方法
依據溫室通常情況下的平均溫度,溫度設為變溫模式22℃(4 h)-25℃(8 h)-28℃(4 h)-25℃(8 h),相對濕度55%—60%,光周期L∶D=16 h∶8 h。極端溫度設定為45℃,熱激時間為2 h,次數為48 h內兩次。
1.2.1 西花薊馬各蟲態存活率 用毛筆分別挑取西花薊馬的1齡若蟲、2齡若蟲、預蛹、蛹、成蟲各60頭分別放于內置新鮮甘藍葉片養蟲瓶中,然后于45℃的培養箱內熱激2 h(培養箱緩慢升溫,到45℃開始計時2 h),之后移到預先設定好的變溫條件下培養,24 h后,再次放于45℃的培養箱內熱處理2 h。記錄各蟲態的存活情況。每處理設5個重復。若蟲和蛹可發育到下一蟲態,記為存活,否則記為死亡。成蟲則用軟毛刷輕觸蟲體,無活動或四肢無抖動判定為死亡。
對照設置:將西花薊馬1齡若蟲、2齡若蟲、預蛹、蛹、成蟲各60頭放置預先設定好的變溫模式22℃(4 h)-25℃(8 h)-28℃(4 h)-25℃(8 h)下培養。

蟲態存活率=1-校正死亡率(圖中此百分率需轉化為小數)
1.2.2 西花薊馬親代 2齡若蟲、成蟲孤雌生殖下親代、F1代的繁殖參數 經過預試驗45℃兩次熱處理1齡若蟲死亡率達到95%,預蛹和蛹死亡率達到100%,故選取成蟲和2齡若蟲法進行下一步試驗。
熱激處理及對照設置:收集西花薊馬2齡若蟲、成蟲于內置新鮮甘藍葉片養蟲瓶中,然后于45℃的培養箱內分別熱激2 h兩次,方法同1.2.1。西花薊馬2齡若蟲或成蟲在預先設定好的變溫模式22℃(4 h)-25℃(8 h)-28℃(4 h)-25℃(8 h)培養后,不經熱激處理,作為對照。
親代成蟲處理:試驗開始時將若干剛羽化西花薊馬成蟲(雌雄均有)置于有甘藍葉片的養蟲瓶中,放于45℃的培養箱內熱處理2 h,之后移到預先設定好的變溫條件下培養,24 h后,再次放于45℃的培養箱內熱處理2 h,待其恢復2 h后,用吸蟲器吸取20頭幸存雌成蟲置于放有紫甘藍葉片的100 mL離心管中,離心管放置于預先設定好的變溫培養箱中培養。每隔24 h更換新鮮的甘藍葉片,并將前一天的甘藍葉片取出,放入相應的9 cm培養皿中,培養皿放置在變溫箱中培養,每天調查1次培養皿中剛孵化出的F1代1齡若蟲數量作為親代產卵量(卵產于葉肉中用肉眼難以觀察),直至離心管中親代雌性薊馬全部死亡。將F1代1齡若蟲全部移走,放入養蟲瓶中,大約7 d后成蟲羽化開始記錄F1代成蟲數,每天記錄1次,直至若蟲全部羽化為成蟲。試驗每個處理重復 3次,共 60頭。統計親代的雌蟲壽命、計數F1代若蟲和成蟲總數量、計算F1代存活率(F1代均為雄蟲,無性比)。
由于西花薊馬孤雌產雄,因此在F1代的雄成蟲剛羽化之時,取20頭雄蟲,另取20頭未經高溫處理的雌成蟲與之配對。每個處理重復3次,共120頭。統計 F1代的雌蟲壽命、計數 F2代若蟲和成蟲總數量、計算F2代存活率以及F2代性比。
親代2齡若蟲處理:45℃熱激處理西花薊馬親代2齡若蟲兩次,待長成成蟲后孤雌生殖下產卵,處理方法同1.2.1。
后代存活率=(成蟲數量/ 1齡若蟲數量)×100%(此百分率在下文表中無需轉換)
F2代性比:F1代所產生的20頭雄蟲與未處理的20頭雌蟲交配,計數后代雌雄數量,雌蟲與雄蟲的比值即為F2性比(3次重復的平均值)。
為方便表達,下文中所有“F1代孤雌生殖”是指親代孤雌產雄生殖得到的F1代,“F2代孤雌生殖”是指未處理的雌蟲與 F1代雄蟲進行有性生殖得到的 F2代。
1.2.3 西花薊馬親代 2齡若蟲、成蟲兩性生殖下親代、F1代的繁殖參數 熱激處理方法同1.2.2,區別為高溫熱激后存活2齡若蟲待其培養到成蟲后,從中選取20頭雌性薊馬和20頭雄性薊馬放入每個離心管中進行兩性生殖,共3組,120頭;高溫熱激后存活的成蟲,從中選取20頭雌性薊馬和20頭雄性薊馬放入每個離心管中進行兩性生殖,共3組,120頭。
取F1代的20頭雄成蟲與20頭雌成蟲配對。每個處理重復3次,共120頭。統計親代F1代的雌蟲壽命、計數F1代和F2代若蟲和成蟲總數量、計算F1代和F2代存活率以及性比。
后代性比:親代處理過的20頭雌蟲與20頭雄蟲交配,F1代雌蟲與雄蟲的比值即為F1代性比(3次重復的平均值);F1代種群內20頭雌蟲與20頭雄蟲交配,F2代雌蟲與雄蟲的比值即為F2代性比(3次重復的平均值)。
1.2.4 西花薊馬體內海藻糖和山梨醇的含量變化海藻糖的測定:將若干西花薊馬成蟲和2齡若蟲置于有甘藍葉片的養蟲瓶中,分別進行上述1.2.1的高溫處理。處理后用吸蟲器吸取活蟲300頭,每個處理5個重復。另外,重新高溫處理西花薊馬成蟲和2齡若蟲,取存活的成蟲和2齡若蟲西花薊馬置于預先設定好的變溫培養箱中培養,親代產卵后分別取 F1代成蟲和F1代2齡若蟲個體,測定其體內海藻糖含量變化。同樣地,培養到F2代個體,測定其體內海藻糖含量變化,從而與親代、F1代進行比較。將親代、F1代、F2代的成蟲和2齡若蟲薊馬(每次重復約300頭)分別放入1.5 mL離心管內,用液氮澆在離心管外部,使薊馬迅速死亡,固定體內成分,置于-80℃超低溫冰箱內,保持其體內物質的穩定性,待測。
參考杜堯[17]的測定方法,制作葡萄糖標準曲線,得出標準曲線的回歸方程,相關系數R2,求出0—l 000 μg·mL-1之間葡萄糖與DNS吸光值的線性關系。
用分析天平稱量出西花薊馬的質量。測定待測液中原有葡萄糖含量Cl值,再測定出酶分解后所含有葡萄糖含量 C2值。依據公式:海藻糖含量=(C2-Cl)×1.05×4/薊馬質量,來計算出樣品海藻糖含量。
山梨醇的測定:將若干西花薊馬成蟲和2齡若蟲置于有甘藍葉片的養蟲瓶中,然后分別進行上述1.2.1的高溫處理。處理方法同上。
首先制作山梨醇含量標準曲線,得出標準曲線的回歸方程,相關系數R2,進而求出0—2 mg·mL-1之間山梨醇含量與吸光度的線性關系。然后制備酶液,加500 μL蒸餾水,5 000 r/min離心10 min,取上清液;用500 μL蒸餾水再離心提取,合并上清液。記下測試液體積V。在655 nm下進行比色,用空白試管溶液調零,記錄吸光度。對比標準曲線,求得標準曲線查得的山梨醇含量C0(ng·mg-1)。
最后求得山梨醇含量:C =(C0×V)/(薊馬質量×l 000)
1.3 數據分析
采用Excel將所得數據繪制表格,通過SPSS19.0進行統計分析,采用LSD檢測法比較不同處理之間的差異顯著性。顯著性水平取P<0.05。
2 結果
2.1 對西花薊馬不同齡期存活率的影響
45℃連續兩次熱激處理西花薊馬不同蟲態2 h后,各個蟲態的存活率都小于50%,且預蛹和蛹的存活率為 0,顯著低于成蟲和 2齡若蟲(F4,20=985.28,P<0.05)。1齡、2齡以及成蟲的存活率分別為5%、21.36% 和 41.38%,存在顯著差異(F4,20=985.28,P<0.05)(圖1)。
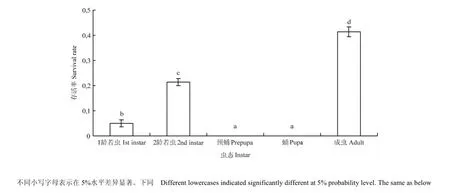
圖1 西花薊馬不同齡期的存活率Fig. 1 The survival rate of different instars of F. occidentalis
2.2 對西花薊馬繁殖力的影響
2.2.1 不同生殖方式下親代、F1代雌蟲的壽命 45℃連續兩次熱激2 h后,對于親代孤雌生殖,45℃熱激當代成蟲其當代雌蟲壽命(34.33 d)顯著高于熱激2齡若蟲(25.33 d),顯著低于對照的(40.67 d)(F2,6=267.17,P<0.05)。親代兩性生殖與親代孤雌生殖有相似的趨勢(F2,6=46.33,P<0.05)。F1代種群有恢復的趨勢。但對于兩性生殖F1代的雌蟲壽命,處理和對照之間仍存在顯著差異(F2,6=22.79,P<0.05)(表1)。
2.2.2 不同生殖方式下F1、F2代的若蟲數 無論是孤雌生殖還是兩性生殖,西花薊馬親代經短時高溫熱激對其后代若蟲數量都有很大的影響,且對照和兩個處理之間在0.05水平差異顯著。對于F1代孤雌生殖,熱激當代成蟲(雌蟲20頭)其后代若蟲數(1 428.33)顯著高于熱激2齡若蟲(1 261.00),顯著低于對照(2 582.33)(F2,6=9 317.13,P<0.05)。對于F1代兩性生殖(雌蟲20頭,雄蟲20頭)與孤雌生殖有相似的趨勢。孤雌生殖F2代若蟲數量顯著高于F1代,但仍低于對照;有性生殖F2代若蟲數量有恢復的趨勢,但與對照間仍存在顯著差異(孤雌:F2,6=166.64,P<0.05;兩性:F2,6=122.73,P<0.05)(表2)。

表1 對孤雌生殖和兩性生殖下親代、F1代雌蟲壽命的影響Table 1 Effect of EHT on adult females longevity of parental generation and F1under parthenogenesis and sexual reproduction conditions
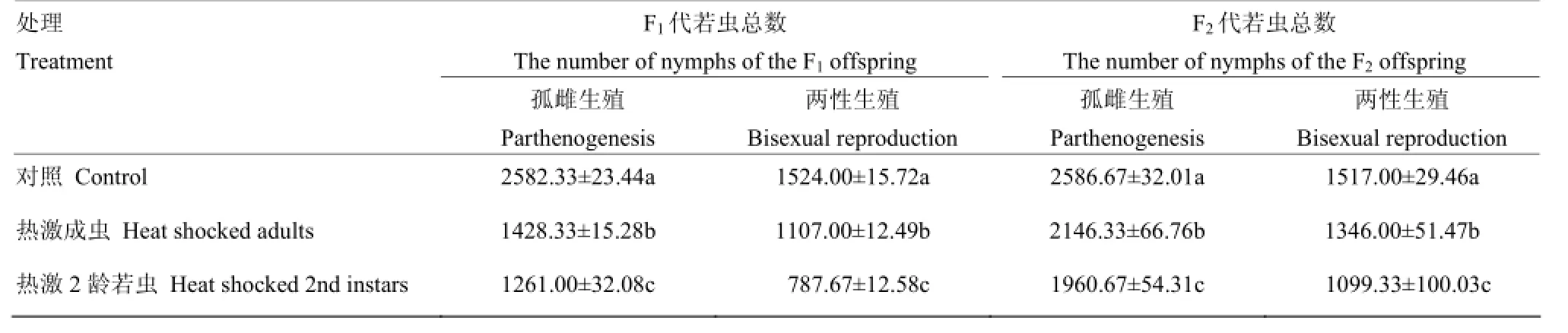
表2 對孤雌生殖和兩性生殖下F1、F2代若蟲數的影響Table 2 Effect of EHT on the numbers of F1, F2nymphs under parthenogenesis and sexual reproduction conditions
2.2.3 不同生殖方式下F1、F2代的成蟲數 西花薊馬親代經歷短時高溫熱激后,對其成蟲的數量也有很大的影響。對于孤雌生殖來說,熱激當代成蟲F1代成蟲數(853.33)顯著高于熱激2齡若蟲(699.67),顯著低于對照(1 905.33)(F2,6=2 571.37,P<0.05)。 兩性生殖和孤雌生殖有相似的趨勢(F2,6=2 186.00,P<0.05)。孤雌生殖F2代成蟲數量顯著高于F1代,但仍低于對照;有性生殖F2代成蟲數量有恢復的趨勢,但與對照間仍存在顯著差異(孤雌:F2,6=110.35,P<0.05;兩性:F2,6=29.34,P<0.05)(表3)。
2.2.4 不同生殖方式下F1、F2代的總存活率 西花薊馬親代經歷短時高溫熱激后,對其后代的總存活率也有很大的影響。對于孤雌生殖來說,熱激當代成蟲和 2齡若蟲其 F1代存活率分別為 59.75%、55.50%,與對照(73.78%)存在顯著差異(F2,6=582.14,P<0.05)。而兩性生殖,熱激當代成蟲和2齡若蟲后其 F1代存活率分別為 60.74%、59.40%,與對照存在顯著差異(F2,6=114.01,P<0.05)。對于 F2代總存活率,無論是孤雌還是兩性生殖,種群整體有恢復趨勢,熱激當代成蟲其F2代和熱激2齡若蟲其F2代存活率分別與對照種群存在顯著差異,而熱激成蟲和熱激 2齡若蟲之間差異不顯著(孤雌:F2,6=19.63,P>0.05;兩性:F2,6=24.44,P>0.05)(表4)。

表3 對孤雌生殖和兩性生殖下F1、F2代成蟲數的影響Table 3 Effect of EHT on the numbers of F1, F2adult under parthenogenesis and sexual reproduction conditions

表4 對孤雌生殖和兩性生殖下F1、F2代總存活率的影響Table 4 Effect of EHT on the survival rates from nymph to adult of F1and F2under parthenogenesis and sexual reproduction conditions
2.2.5 不同生殖方式下 F1、F2代的性比 短時高溫熱激對西花薊馬性比(♀∶♂)的影響較大。對于兩性生殖下F1代,熱激當代成蟲其F1代性比(2.53∶1)高于熱激2齡若蟲(2.42∶1),低于對照(2.66∶1)。對于兩性生殖下F2代,熱激成蟲其F2代的性比(2.43∶1)低于對照種群(2.77∶1),但是熱激成蟲和熱激2齡若蟲其F2代的性比差異不明顯。對于孤雌生殖來說,F1代全部為雄性,無性比。F2代中對照的性比(2.65∶1)高于其他兩個處理(熱激成蟲:2.43∶1;熱激2齡若蟲2.41∶1)(表5)。

表5 對孤雌生殖和兩性生殖下F1、F2代性比的影響Table 5 Effect of EHT on the sex ratio (female∶male) of F1and F2under parthenogenesis and sexual reproduction conditions
2.3 對西花薊馬體內海藻糖含量的影響
2.3.1 成蟲體內海藻糖含量的變化 西花薊馬經過45℃高溫2 h連續兩次處理后,其成蟲體內的海藻糖含量變化較大。對照西花薊馬成蟲體內的海藻糖含量為17.23 mg·g-1,而 45℃處理西花薊馬當代成蟲后,其體內的海藻糖含量降低為8.43 mg·g-1,顯著低于對照種群(F5,24=130.11,P<0.05)。熱激當代成蟲和若蟲后,其F1代成蟲體內的海藻糖含量(熱激成蟲:12.86 mg·g-1;熱激若蟲11.73 mg·g-1)顯著低于對照成蟲體內的海藻糖含量(F5,24=130.11,P<0.05)。F2代成蟲體內的海藻糖含量與 F1代相比有所上升,但與對照仍然差異顯著(F5,24=130.11,P<0.05)。熱激當代成蟲其F1代成蟲體內的海藻糖含量(12.86 mg·g-1)顯著高于熱激當代若蟲其F1代的成蟲體內含量(11.73 mg·g-1),且F2代與F1代趨勢相同(F5,24=130.11,P<0.05)(圖2)。
2.3.2 若蟲體內海藻糖含量的變化 西花薊馬經過45℃高溫2 h連續兩次處理后,其若蟲體內的海藻糖含量變化明顯。對照西花薊馬的若蟲體內海藻糖含量為27.12 mg·g-1,而45℃處理西花薊馬當代若蟲后,其體內的海藻糖含量降低為12.04 mg·g-1,顯著低于對照種群(F5,24=384.27,P<0.05)。熱激當代成蟲和若蟲后,其F1代若蟲體內的海藻糖含量(熱激成蟲:16.48 mg·g-1;熱激若蟲14.20 mg·g-1)顯著低于對照若蟲體內的海藻糖含量(F5,24=384.27,P<0.05)。F2代若蟲體內的海藻糖含量與F1代相比有所上升,但與對照差異顯著(F5,24=384.27,P<0.05)。熱激當代成蟲其F1代若蟲體內的海藻糖含量(16.48 mg·g-1)顯著高于熱激當代若蟲其 F1代若蟲體內的含量(14.20 mg·g-1),且F2代與F1代趨勢相同(F5,24=384.27,P <0.05)(圖3)。
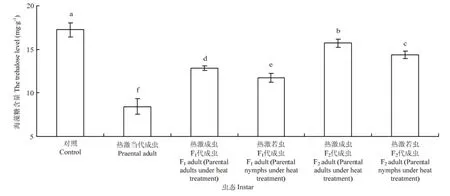
圖2 對親代、F1代、F2代成蟲體內海藻糖含量的影響Fig. 2 Effect of EHT on the trehalose level in adult of parent generation, F1and F2
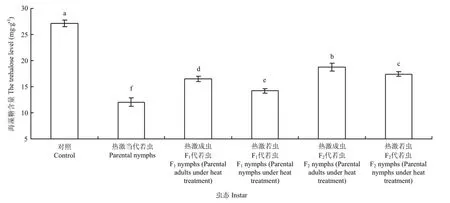
圖3 對親代、F1代、F2代若蟲體內海藻糖含量的影響Fig. 3 Effect of EHT on the trehalose level in nymphs of parent generation, F1and F2
2.4 對西花薊馬體內山梨醇含量的影響
2.4.1 成蟲體內山梨醇含量的變化 西花薊馬經過45℃高溫2 h連續兩次處理后,其成蟲體內的山梨醇含量變化較大。對照西花薊馬的成蟲體內山梨醇含量為0.95 ng·mg-1,而 45℃處理西花薊馬當代成蟲后,其體內的山梨醇含量上升為 2.73 ng·mg-1,顯著高于對照種群(F5,24=554.57,P<0.05)。熱激當代成蟲和若蟲后,其F1代成蟲體內的山梨醇含量(熱激成蟲:1.45 ng·mg-1;熱激若蟲:1.38 ng·mg-1)顯著高于對照成蟲體內的山梨醇含量(0.95 ng·mg-1)(F5,24=318.30,P<0.05)。而F2代成蟲體內的山梨醇含量與F1代相比有所降低,分別為1.22、1.15 ng·mg-1,但與對照仍然存在差異顯著性(F5,24=318.30,P<0.05)。熱激當代成蟲其F1代成蟲體內的山梨醇含量(1.45 ng·mg-1)與熱激當代若蟲其F1代成蟲體內的含量(1.38 ng·mg-1)無明顯差異,F2代也無明顯差異(F5,24=384.27,P>0.05)(圖4)。
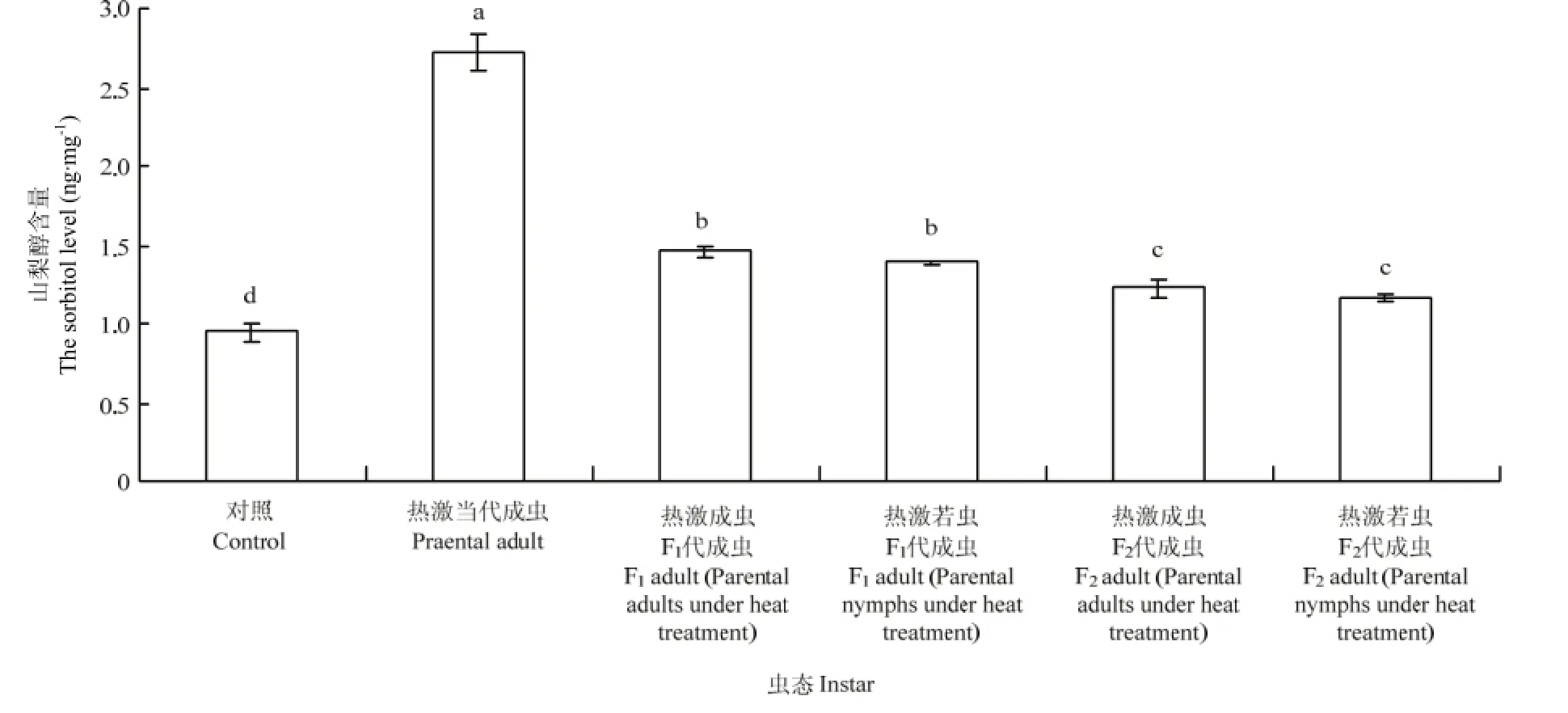
圖4 對親代、F1代、F2代成蟲體內山梨醇含量的影響Fig. 4 Effect of EHT on the sorbitol level in adult of parent generation, F1and F2
2.4.2 若蟲體內山梨醇含量的變換 西花薊馬經過45℃高溫2 h連續兩次處理后,其若蟲體內的山梨醇含量的變化明顯。對照條件下,西花薊馬的若蟲體內山梨醇含量為0.70 ng·mg-1,而45℃處理西花薊馬當代若蟲后,其體內的山梨醇含量上升為2.13 ng·mg-1,顯著高于對照種群(F5,24=318.30,P<0.05)。熱激當代成蟲和若蟲后,其F1代若蟲體內的山梨醇含量(熱激成蟲:1.63 ng·mg-1;熱激若蟲1.61 ng·mg-1)顯著高于對照若蟲體內的山梨醇含量(0.70 ng·mg-1)(F5,24=318.30,P<0.05)。F2代若蟲體內的山梨醇含量與F1代相比有所降低,熱激成蟲后F2代若蟲(0.84 ng·mg-1)仍與對照有差異顯著性(F5,24=318.30,P<0.05),而熱激若蟲后F2代若蟲(0.70 ng·mg-1)與對照無差異顯著性。熱激當代成蟲其F1代若蟲體內的山梨醇含量(1.63 ng·mg-1)與熱激當代若蟲其F1代若蟲體內的含量(1.61 ng·mg-1),但熱激當代成蟲其 F2代若蟲體內的山梨醇含量(0.84 ng·mg-1)顯著高于熱激當代若蟲其 F2代若蟲體內的含量(0.70 ng·mg-1)(F5,24=384.27,P<0.05)(圖5)。
3 討論
3.1 高溫對西花薊馬存活率的影響
高溫可導致昆蟲體內水分的散失[18]、膜結構的破壞和蛋白的降解,從而限制了相關酶催化的反應[19]。同時,高溫對昆蟲神經系統的傷害常導致許多功能的喪失[20-21],而這些傷害主要反映在昆蟲存活率上。前期結果顯示,西花薊馬成蟲在45℃熱激一次2 h后,其存活率在80%左右,本研究在此結果的基礎上進行連續兩次熱激,結果表明西花薊馬各個蟲態的存活率極低,并且不同蟲態表現出的耐熱性不同,預蛹和蛹的存活率為0,1齡若蟲的存活率為5%。相比較而言,成蟲的耐熱性是最高的,其次是2齡若蟲。這表明,45℃的極端高溫對西花薊馬的存活極其不利,這可能是由于如此的高溫超出了西花薊馬所能承受的生理極限。西花薊馬成蟲的耐熱性是最高的,推測原因可能是由于若蟲的體表較柔軟,角質層較薄;而成蟲的體表相對堅硬,角質層較厚,降低了體內水分損失,保持體內的正常生命活動的進行。西花薊馬蛹期的耐熱性最低,可能是由于蛹期時西花薊馬幾乎處于不食不動的狀態,無法迅速躲避外界高溫。
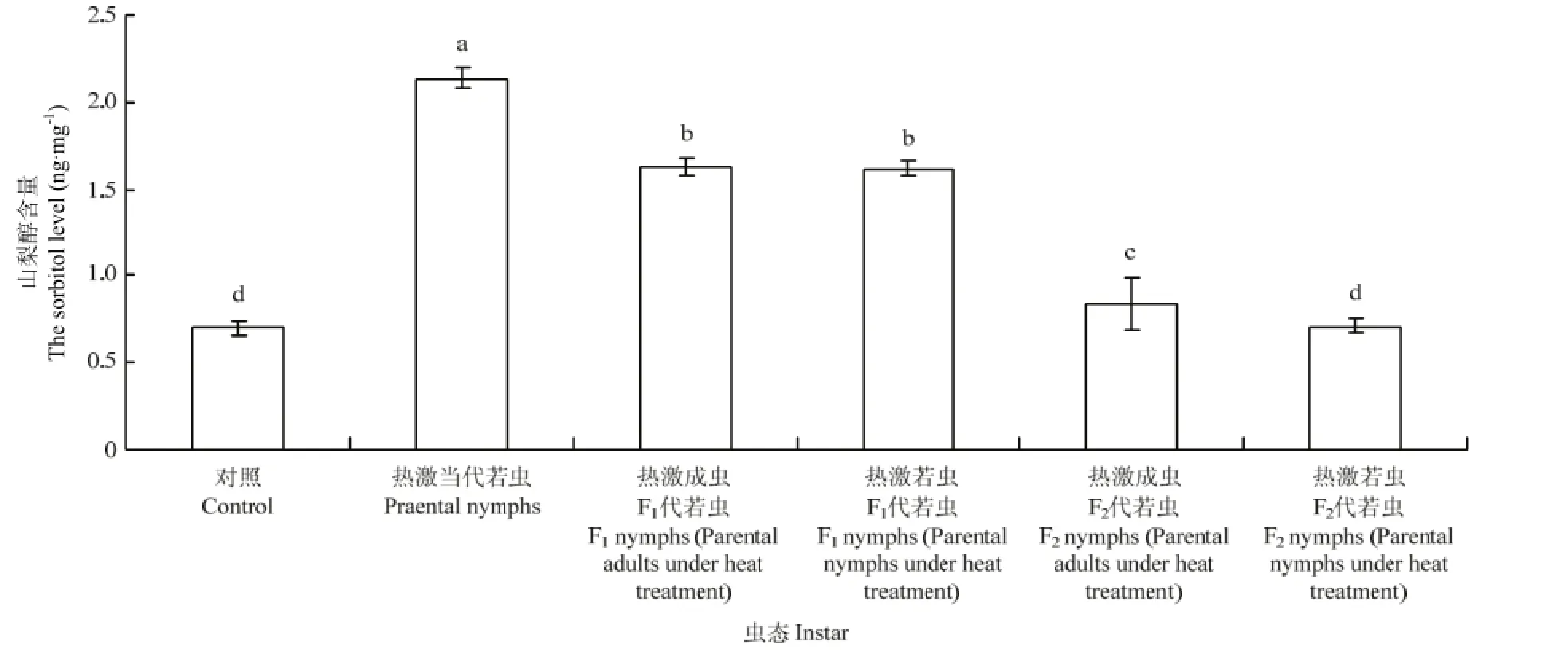
圖5 對親代、F1代、F2代若蟲體內山梨醇含量的影響Fig. 5 Effect of EHT on the sorbitol level in nymphs of parent generation, F1and F2
3.2 高溫對西花薊馬生殖的影響
一般而言,大多數昆蟲存活及發育的上限溫度范圍為 40—50℃。超出此范圍后,昆蟲將面臨兩個結局:存活或死亡[7]。此外,即使一部分昆蟲在遭遇高溫后能存活下來,但其適合度必定會受到影響[22]。研究發現,煙粉虱雌蟲的壽命從20℃時的44.36 d下降到35℃時的10.43 d[23]。小菜蛾雌蟲的壽命也隨著溫度升高而明顯縮短,15℃下46.4 d,35℃下平均壽命僅為3.6 d[24]。前期試驗結果顯示,西花薊馬種群在45℃的高溫2 h熱激一次后,雌成蟲壽命從34.65 d縮短到27.91 d[25]。本研究顯示,45℃的高溫2 h熱激兩次后,無論是孤雌生殖還是兩性生殖,雌性成蟲的壽命都明顯縮短,甚至影響到F2代雌蟲壽命。這可能是西花薊馬在抵抗高溫脅迫時消耗了部分能量或者體內生理物質發生改變,影響了以后的生存。
溫度對昆蟲繁殖起著至關重要的作用。許多研究表明,在高溫作用下,昆蟲的生殖受到明顯地抑制。研究人員發現,將棉鈴蟲暴露在 42—46℃的高溫范圍一段時間后,其產卵量受到嚴重影響。當在 40、42.5、45℃下暴露360、120、15 min后,棉鈴蟲在整個生活史時期內均不產卵[26]。在相關的研究中發現,將紅尾肉蠅(Sarcophaga crassipalpis)暴露于45℃ 1 h后,其產卵量僅為對照的10%。更為重要的是,這些卵全部不可育[22]。Q型煙粉虱(Bemisia tabaci)成蟲在44℃下暴露1 h后單頭雌蟲的產卵量為77. 2粒,低于常溫 26℃下的產卵量 100.1粒[27];溫室白粉虱(Trialeurodes vaporariorum)成蟲在37—43℃暴露2 h后單雌產卵量由62.4粒下降到1.5粒,45℃暴露2 h后甚至停止了產卵[28]。另外麥無網長管蚜(Metopolophium dirhodum)經過高于29℃的高溫處理后,產卵量和壽命明顯降低,且高溫對成蟲期的傷害要大于若蟲期[29]。與上述研究不同的是,本試驗的結果是熱激對若蟲的傷害大于成蟲,F1代有性生殖結果顯示,熱激親代成蟲和若蟲后產卵量從對照的1 524.00粒分別下降至1 107.00粒和787.67粒。無論是兩性生殖還是孤雌生殖,F1代種群數量都明顯減少,兩個處理間和對照都有顯著性差異。對于孤雌生殖F2代,由于與F1代雄成蟲交配的雌成蟲是未經處理,所以產卵量高于孤雌生殖F1代,但低于對照,這說明高溫對F1代雄性有一定影響。對于有性生殖 F2代,種群有明顯的恢復,但是與對照仍然存在顯著性差異。分析其原因,在多數昆蟲中,高溫能破壞雌雄兩性的生殖系統功能[30-31]。已有的研究表明,高溫能夠影響雌蟲卵巢和卵子的發育,從而導致產卵量的降低。此外,高溫也能對雄蟲的輸精管及精子造成傷害,導致其不育[22,32],從而導致F1代種群降低。
熱激對西花薊馬后代性比和存活率也有很大的影響。關于高溫對昆蟲后代存活率和性比的影響,在不同昆蟲上有不同表現:趙鑫等[33]研究表明,蓮草直胸跳甲(Agasicles hygrophila)經短時高溫暴露后,后代存活率下降;另有研究表明,溫度對煙粉虱的性比變化有影響[34]。B型煙粉虱成蟲在比較溫和的高溫下(39 和41℃)熱激l h,其后代中雌蟲的比例上升;經過43℃及以上高溫暴露后其后代性比產生逆轉,雄蟲所占的比例上升。在本試驗中,孤雌生殖情況下,熱激西花薊馬成蟲、若蟲2 h后,F1代總存活率和對照都有顯著差異,且熱激當代成蟲的存活率高于熱激2齡若蟲;F2代兩個處理與對照有顯著差異,但兩個處理間差異不大。F2代的總存活率總體來說高于F1代,仍與對照存在顯著差異。熱激處理后F1代兩性生殖的性比和對照相比明顯降低,兩個處理間也有所差異。F2代處理與對照仍有差異,但是處理間差異不大。造成這一現象的原因可能是西花薊馬孤雌生殖產生雄蟲,兩性生殖產雌蟲和雄蟲,西花薊馬親代在45℃高溫處理后可能出現雄性不育的現象,導致兩性生殖產雌率降低,后代雌雄比例降低。而F2代種群整體有所恢復,因此會有更多的雌性薊馬選擇兩性生殖,F2代就會產生更多的雌性個體,雌雄性比在 F2代相對增加,F2代以后的種群數量就會趨于穩定。
綜上所述,親代進行短時高溫暴露后,存活率降低,F1代繁殖速度相應變慢,這些都是西花薊馬在短時高溫下的不利反應。而F1代可以通過自身的種群調節,保證相應齡期的存活率和產生較多的雌性后代來保證后代種群的延續,降低了高溫暴露對后代不利影響的累積效應。
3.3 高溫對西花薊馬小分子物質變化的影響
3.3.1 高溫對西花薊馬海藻糖變化的影響 昆蟲屬于變溫動物,保持和調節體內溫度的能力不強。因此,昆蟲的代謝速率十分依賴于環境溫度的變化。一旦溫度出現急劇變化,將引發其體內一系列代謝反應。海藻糖是昆蟲體內的主要糖分,主要用于滿足昆蟲的各種需能活動,細胞內高含量的海藻糖能保護生物分子抵御傷害,增強生存能力。海藻糖能在海藻糖酶的水解作用下轉化成葡萄糖,而葡萄糖在昆蟲體內糖類代謝過程中可與糖原互相轉化,參與能量代謝[10]。西花薊馬成蟲和2齡若蟲受到45℃高溫處理后其體內的海藻糖含量明顯降低,這說明在熱激處理后,其體內的海藻糖已被海藻糖酶分解為葡萄糖,大部分用于抵抗高溫脅迫,因此不足以為其正常的生命活動提供足夠的能量,從而影響其健康性甚至存活,這方面可以用于解釋高溫能夠降低西花薊馬的存活率。另外,除了能量代謝,海藻糖在昆蟲生殖腺和生殖細胞發育等方面也起著重要的作用。其含量的顯著改變,將破壞昆蟲正常生理功能,使之生理狀態紊亂,生長發育受阻。這一點也可以解釋西花薊馬不同蟲態經過高溫處理后,其F1代若蟲總數、成蟲總數和后代總存活率都下降。結果結果還顯示,西花薊馬親代經過45℃熱激處理后,親代成蟲體內的海藻糖含量明顯低于2齡若蟲的,這說明親代成蟲大部分的海藻糖被分解用于抵抗高溫脅迫,再次證明了西花薊馬成蟲的耐熱性高于 2齡若蟲的,對其后代影響較小,這與高溫熱激成蟲后其F1、F2代若蟲數和成蟲數較高相吻合。相似的結果也表現在恒溫條件下培養的西花薊馬種群(數據未發表)。
3.3.2 高溫對西花薊馬山梨醇變化的影響 在高溫脅迫下,昆蟲可能通過合成小分子物質來抵御高溫的傷害。例如,在高溫脅迫下蚜蟲和粉虱體內的山梨醇含量顯著增加,由此推測,蚜蟲和粉虱將從食物中獲得果糖作為底物,在NADPH酮糖還原酶的作用下將果糖轉變成甘露醇或山梨醇[15]。同時也有研究發現,甘露醇和山梨醇在昆蟲體內的含量隨氣溫變化而有所差異。在溫度較高的中午,2種物質的含量要顯著高于早晨[15]。因此,甘露醇和山梨醇在昆蟲抵御高溫過程中可能具有重要作用[14,35]。本研究表明,西花薊馬親代成蟲和2齡若蟲經過45℃熱激處理后,其體內的山梨醇含量明顯升高,且在F1、F2代種群中仍然存在較高的含量,這說明高溫熱激后,西花薊馬取食量增加,從而將食物中的果糖轉變成山梨醇。另外西花薊馬親代不同蟲態經熱激處理后,成蟲體內的山梨醇含量明顯高于若蟲,說明在高溫脅迫下成蟲的取食量大于2齡若蟲,與高溫影響西花薊馬繁殖力的生態數據相吻合(熱激處理西花薊馬親代2齡若蟲,其成蟲壽命低于熱激處理成蟲,F1、F2代若蟲數和成蟲數低于熱激處理成蟲)。因此,在西花薊馬若蟲遭受高溫脅迫時,其適合度和體內小分子物質的積累受到的影響大于成蟲期,在進行生物化學防治時可以著重考慮若蟲期的悶棚和施藥。
4 結論
極端高溫處理對西花薊馬生殖適應性有很大的影響,從生態角度上來看,45℃高溫2 h連續2 d處理會導致西花薊馬的各個指標明顯下降,各蟲態的存活率降低、雌成蟲壽命縮短、后代種群數量波動以及性比的降低,整個種群的生長發育受到影響,甚至F2代種群的規模也受到影響;另外高溫脅迫也會影響西花薊馬體內海藻糖和山梨醇含量,若蟲階段的高溫處理對西花薊馬生態和生理的影響比成蟲階段明顯。而且,熱激親代蟲態之后對其后代有持續性的影響,并隨代數的增加影響逐漸減小。
西花薊馬具有個體小、隱蔽性危害、繁殖快等特點,農藥防治易產生抗藥性,而且會對環境造成污染;而高溫悶棚技術無污染、低成本、易操作,45—46℃悶棚2 h兩次對寄主植物并無太大影響,且能在短時間內在一定程度上控制種群快速增長,是生產實踐中溫室防治該害蟲的一項值得推廣應用的技術措施。
References
[1] HALLMAN G J, DENLINGER D L. Introduction: temperature sensitivity and integrated pest management//HALLMAN G J,DENLINGER D L. Temperature Sensitivity in Insects and Application in Integrated Pest Management. Boulder, CO: Westview Press, 1998:1-5.
[2] BALE J S, HAYWARD S A L. Insect overwintering in a changing climate. The Journal of Experimental Biology, 2010, 213: 980-994.
[3] FIELDS P G. The control of stored-product insects and mites with extreme temperatures. Journal of Stored Products Research, 1992,28(2): 89-118.
[4] MOURIER H, POULSEN K P. Control of insects and mites in grain using a high temperature/short time (HTST) technique. Journal of Stored Products Research, 2000, 36: 309-318.
[5] DENLINGER D L, YOCUM G D. Physiology of heat sensitivity// HALLMAN G J, DENLINGER D L. Temperature Sensitivity in Insects and Application in Integrated Pest Management. Boulder, CO:Westview Press, 1998: 11-18.
[6] RINEHART J R, YOCUM G D, DENLINGER D L. Thermotolerance and rapid cold hardening ameliorate the negative effects of brief exposures to high or low temperatures on fecundity in the flesh fly,Sarcophaga crassipalpis. Physiological Entomology, 2000, 25:330-336.
[7] GULLAN P J, CRANSTON P S. The Insects: An Outline of Entomology. 3rd ed. Davis, USA: Blackwell Publishing Ltd., 2005.
[8] 戴秀玉, 程蘋, 周堅, 江慧修. 海藻糖的生理功能、分子生物學研究及應用前景. 微生物學通報, 1995, 22(2): 102-103. DAI X Y, CHENG P, ZHOU J, JIANG H X. Physiological function,molecular biology and applications of trehalose. Microbiology Bulletin, 1995, 22(2): 102-103. (in Chinese)
[9] 聶凌鴻, 寧正祥. 海藻糖的生物保護作用. 生命的化學, 2001,21(3): 206-209. NIE L H, NING Z X. Biological protective function of the trehalose. Chemistry of Life, 2001, 21(3): 206-209. (in Chinese)
[10] CROWE J, CROWE L, CHAPMAN D. Preservation of membranes in anhydrobiotic organisms: The role of trehalose. Science, 1984,223(4637): 701-703.
[11] SALVUCCI M E, HENDRIX D L, WOLFE G R. Effect of high temperature on the metabolic processes affecting sorbitol synthesis in the silverleaf whitefly, Bemisia argentifolii. Journal of Insect Physiology, 1999, 45(1): 21-27.
[12] SALVUCCI M E, STECHER D S, HENNEBERRY T J. Heat shock proteins in whiteflies, an insect that accumulates sorbitol in response to heat stress. Journal of Thermal Biology, 2000, 25: 363-371.
[13] HENDRIX D L, SALVUCCI M E. Polyol metabolism in homopterans at high temperatures: accumulation of mannitol in aphids (Aphididae:Homoptera) and sorbitol in whiteflies (Aleyrodidae: Homoptera). Comparative Biochemistry and Physiology, A, 1998, 120(3): 487-494.
[14] KIRK W D J, TERRY L I. The spread of the western flower thrips Frankliniella occidentalis (Pergande). Agricultural and Forest Entomology, 2003, 5: 301-310.
[15] CHILDERS C C, ACHORS D S. Thrips feeding and oviposition injuries to economic plants, subsequent damage and host response to infestation//PARKER B L, SKINNER M, LEWIS T. Thrips Biology and Management. New York and London: Plenum Press, 1995: 31-50.
[16] 周永豐, 唐峻嶺. 高溫對南美斑潛蠅的致死作用. 昆蟲知識, 2003,40(4): 372-373. ZHOU Y F, TANG J L. Lethal effect of high temperature on the leafminer fly, Liriomyza huidobrensis in a greenhouse. Entomological Knowledge, 2003, 40(4): 372-373. (in Chinese)
[17] 杜堯. 高溫模式對麥蚜實驗種群影響的生態機制研究[D]. 北京:中國農業科學院, 2007. DU Y. Ecological mechanism of effects of high temperature patterns on laboratory population of cereal aphid[D]. Beijing: Chinese Academy of Agricultural Sciences, 2007. (in Chinese)
[18] YODER J A, CHAMBERS M J, TANK J L, KEENEY G D. High temperature effects on water loss and survival examining the hardiness of female adults of the spider beetles, Mezium affine and Gibbium aequinoetiale. Journal of Insect Science, 2009, 9: Article 68.
[19] CHOWN S L, NICHOLSON S W. Insect Physiological Ecology:Mechanisms and Patterns. New York: Oxford Press, 2004.
[20] ROBERTSON R M. Modulation of neural circuit operation by prior environmental stress. Integrative and Comparative Biology, 2004,44(1): 21-27.
[21] CHOWN S L, TERBLANCHE J S. Physiological diversity in insects:Ecological and evolutionary contexts. Advances in Insect Physiology,2006, 33: 50-152.
[22] RINEHART J R, YOCUM G D, DENLINGER D L. Thermotolerance and rapid cold hardening ameliorate the negative effects of brief exposures to high or low temperatures on fecundity in the flesh fly,Sarcophaga crassipalpis. Physiological Entomology, 2000, 25:330-336.
[23] DROST Y C, VAN LENTEREN J C, VAN ROERMUND H J W. Life-history parameters of Bemisia tabaci (Hemiptera: Aleyrodidae) in relation to temperature and host plant: a selective review. Bulletin of Entomological Research, 1998, 88(3): 219-229.
[24] 劉蕓, 阮傳清, 劉波, 朱育菁. 溫度對小菜蛾成蟲繁殖和壽命的影響. 中國農學通報, 2013, 29(12): 190-193. LIU Y, RUAN C Q, LIU B, ZHU Y J. Effects of temperature on oviposition and longevity of adult diamondback moth (Plutella xylostella L.). Chinese Agricultural Science Bulletin, 2013, 29(12):190-193. (in Chinese)
[25] WANG J C, ZHANG B, WANG J P, LI H G, WANG S F, SUN L J,ZHENG C Y. Effects of heat stress on survival of Frankliniella occidentalis (Thysanptera: Thripidae) and Thrips tabaci (Thysanoptera:Thripidae). Journal of Economic Entomology, 2014, 107(4): 1426-1433.
[26] MIRONIDIS G K, SAVOPOULOU-SOULTANI M. Effects of heat shock on survival and reproduction of Helicoverpa armigera (Lepidoptera: Noctuidae) adults. Journal of Thermal Biology, 2010,35: 59-69.
[27] 朱紹光, 李照會, 萬方浩. 短時高溫暴露對Q型煙粉虱存活和生殖適應性的影響. 昆蟲知識, 2010, 47(6): 1141-1144. ZHU S G, LI Z H, WAN F H. Effects of brief exposure to high temperature on survival and reproductive adaptation of Bemisia tabaci Q-biotype. Chinese Bulletin of Entomology, 2010, 47(6): 1141-1144. (in Chinese)
[28] 崔旭紅, 謝明, 萬方浩. 短時高溫暴露對B型煙粉虱和溫室白粉虱存活以及生殖適應性的影響. 中國農業科學, 2008, 41(2): 424-430. CUI X H, XIE M, WAN F H. Effects of brief exposure to high temperature on survival and fecundity of two whitefly species: Bemisia tabaci B-biotype and Trialeurodes vaporariorum (Homoptera:Aleyrodidae). Scientia Agricultura Sinica, 2008, 41(2): 424-430. (in Chinese)
[29] MA C S, HAU B, POEHLING B H. Effects of pattern and timing of high temperature exposure on reproduction of the rose grain aphid,Metopolophium dirhodum. Entomologia Experimentalis et Applicata,2004, 110: 65-71.
[30] MAHROOF R, ZHU K Y, SUBRAMANYAM B. Changes in expression of heat shock proteins in Tribolium castaneum (Coleoptera:Tenebrionidae) in relation to developmental stage, exposure time, and temperature. Annals of the Entomological Society of America, 2005,98(1): 100-107.
[31] CUI X, WAN F H, XIE M, LIU T X. Effects of heat shock on survival and reproduction of two whitefly species, Trialeurodes vaporariorum and Bemisia tabaci biotype B. Journal of Insect Science , 2008, 8(24):Article 24.
[32] SCOTT M, BERRIGAN D, HOFFMANN A A. Costs and benefits of acclimation to elevated temperature in Trichogramma carverae. Entomologia Experimentalis et Applicata, 1997, 85: 211-219.
[33] 趙鑫, 傅建煒, 萬方浩, 郭建英, 王進軍. 短時高溫暴露對蓮草直胸跳甲生殖特性的影響. 昆蟲學報, 2009, 52(10): 1110-1114. ZHAO X, FU J W, WAN F H, GUO J Y, WANG J J. Effect of brief high temperature exposure on reproduction characteristics of Agasicles hygrophila (Coleoptera: Chrysomelidae). Acta Entomologica Sinica,2009, 52(10): 1110-1114. (in Chinese)
[34] ENKEGAARD A. Encarsia formosa parasitizing the Poinsettia-strain of the cotton whitefly, Bemisia tabaci, on Poinsettia: bionomics in relation to temperature. Entomologia Experimentalis et Applicata,1993, 69(3): 251-261.
[35] CHOWN S L, SORENSEN J G, TERBLANCHE J S. Water loss in insects: An environmental change perspective. Journal of Insect Physiology, 2011, 57(8): 1070-1084.
(責任編輯 岳梅)
Effects of Extreme High Temperature on Survival Rate,Reproduction, Trehalose and Sorbitol of Frankliniella occidentalis
JIANG Shan1, LI Shuai1, ZHANG Bin1, LI Hong-gang1,2, WAN Fang-hao1,3, ZHENG Chang-ying1
(1Key Laboratory for Insect Ecology of Shandong Province, College of Agronomy and Plant Protection, Qingdao Agricultural University, Qingdao 266109, Shandong;2Plant Protection Station of Shandong Province, Jinan 250100;3Key Laboratory for Biology of Plant Diseases and Insect Pests, Institute of Plant Protection,Chinese Academy of Agricultural Sciences, Beijing 100193)
Abstract:【Objective】The western flower thrips (Frankliniella occidentalis) is an important invasive and quarantine pest in China which created massive economic losses in vegetables and flowers. The objective of this study is to explore the influence of extreme high temperature (EHT) on survival rate, fecundity and content of trehalose and sorbital of F. occidentalis, and to provide a basis for the control of F. occidentalis. 【Method】After 2nd nymphs and adults exposed to 45℃ for 2 h twice (24 h interval under variable temperatures 22℃ (4 h)-25℃ (8 h)-28℃ (4 h)-25℃ (8 h) between two heat treatments), the survival rate, female adult longevity, offspring population dynamics, sex ratio and content of trehalose and sorbital were detected and analyzed. 【Result】In comparison with the control, the survival rate of each stage was less than 50% (0 for 3rd and 4th nymph, 41.38% for female adults,5% for 1st nymphs and 21.36% for 2nd nymphs), parental and offspring female adult longevity and fecundity significantly reduced (P<0.05), the content of trehalose in 2nd nymphs and adults significantly declined (P<0.05), and the content of sorbital in 2nd nymphs and female adults significantly increased after heat-shocked on parental 2nd nymphs and female adults (P<0.05). Meanwhile, higher female adult longevity and fecundity, lower content of trehalose, higher content of sorbital in F. occidentalis sufferred from twice 45℃ for 2 h heat shock on parental adults was expressed more than those in F. occidentalis sufferred from heat-shock on parental 2nd nymphs. Interestingly, the influence of twice 45℃ for 2 h heat shock not only occurred on parental generation but subsequent generations (F1and F2generations), and the trend of F1and F2generations was the same as the parental generation.【Conclusion】Twice 45℃ for 2 h exposes on parental F. occidentalis significantly affected the development, reproduction and physiology of present and offspring generations. The changes of trehalose and sorbital content had a correlation with the population dynamics of F. occidentalis.
Key words:extreme high temperature; Frankliniella occidentalis; fecundity; trehalose; sorbitol
收稿日期:2016-01-22;接受日期:2016-04-07
基金項目:國家自然科學基金(31372033)、山東省現代農業產業技術體系(SDAIT-02-021-11)、“泰山學者”建設工程專項
