巴西橡膠樹PPCK基因家族成員的鑒定和表達分析
2016-07-09 18:39:14肖小虎隋金蕾戚繼艷陽江華朱晉恒唐朝榮
熱帶作物學報 2016年11期
肖小虎 隋金蕾 戚繼艷 陽江華 朱晉恒 唐朝榮
摘 要 以已發表的PPCK序列為探針,對橡膠樹和其他5種植物(木薯、水稻、楊樹、蓖麻和擬南芥)的轉錄組和基因組進行全面搜索,鑒定得到17個PPCK基因,其中包括3個橡膠樹PPCK基因,命名為HbPPCK1-3。序列分析發現,HbPPCK1-3和其他植物一樣在結構上非常保守,只有一個內含子和一個蛋白激酶結構域,并且相對位置也比較一致。在進化上,橡膠樹、木薯和蓖麻3種大戟科植物的PPCK蛋白具有較近的親緣關系。HbPPCK3在樹皮中表達豐度最高,其次是種子和雄花;HbPPCK2在膠乳和根中表達豐度相對較高,而HbPPCK1在各組織中的表達都比較低。另外,大部分家族成員的表達都受到真菌侵染、低溫和干旱脅迫處理的誘導,而乙烯利處理卻能抑制HbPPCK2的表達。
關鍵詞 巴西橡膠樹;PPCK;基因家族;結構和進化;表達分析
中圖分類號 S794.1 文獻標識碼 A
在植物中,磷酸烯醇式丙酮酸羧化酶(PEPC)以Mg2+或Mn2+為輔助因子, 催化磷酸烯醇式丙酮酸(PEP)和HCO3-生成草酰乙酸(OAA)和無機磷酸的不可逆性反應。PEPC廣泛存在于光合生物, 如植物、藻類、藍細菌和光合細菌中, 還存在于很多非光合細菌和原生動物中。PEPC催化的反應為細胞各種組分的生物合成提供四碳二羧酸, 參與維持檸檬酸循環,在初級代謝中有重要的補給作用,在C4植物和CAM植物的光合作用中催化大氣中CO2固定的第一步反應,是C4光合作用途徑中最重要的酶之一[1]。PEPC的活性主要是通過磷酸烯醇式丙酮酸羧化酶激酶(phosphoenolpyruvate carboxylase kinases,PPCKs)來調節[2]。PPCKs是鈣調控型蛋白激酶的一員,但卻缺少鈣依賴型蛋白激酶所特有的自抑區(auto-inhibitory)和EF-hands結構域,所以PPCK是一種非鈣依賴型蛋白激酶[2-3]。在景天酸代謝植物玉吊鐘(Kalanchoe fedtschenkoi)、C4植物玉米已有研究和C3植物擬南芥中已有相關研究[3-5],研究認為PPCK的表達主要通過轉錄水平進行調控。在橡膠樹中,橡膠的生產通過割膠來完成,割膠對橡膠樹來說是一種機械傷害,而作為一種傷害信號卻推動了橡膠的生物合成。人工使用乙烯利進行刺激可以顯著提高膠乳的產量,正是應用了橡膠樹樹皮對激素刺激的應答能力,進而使橡膠樹光合作用產物流向橡膠樹的生物合成[6-7]。本研究以已發表PPCK蛋白氨基酸序列為探針[3-5],對橡膠樹、木薯、蓖麻、擬南芥、楊樹和水稻6種植物的全基因組數據進行全面搜索,鑒定得到17個PPCK基因家族成員,其中橡膠樹中有3個。筆者利用相關軟件和數據庫,對橡膠樹PPCK基因家族成員的基因結構、進化和表達水平進行了初步分析。研究結果將有助于筆者深入了解橡膠樹PPCK基因在橡膠樹生長發育和逆境脅迫應答過程中的功能。
1 材料與方法
1.1 材料
本實驗室Solexa測序所用的巴西橡膠樹(Hevea brasiliensis)熱研7-33-97材料,除了根來自于組培苗外,其他組織(包括膠乳、樹皮、樹葉、種子、雌花和雄花)均來自中國熱帶農業科學院試驗場三隊的正常割膠橡膠樹(開割2 a以上),根來自于中國熱帶農業科學院橡膠研究所種質資源圃華玉偉老師贈送的熱研7-33-9組培苗;不同發育時期的橡膠樹葉片來自于中國熱帶農業科學院橡膠所種質資源圃,為1年生的熱研7-33-97嫁接苗;乙烯利處理的材料主要來自于海南省儋州市中國熱帶農業科學院試驗場三隊,品系為熱研7-33-9,正常開割樹(3天1刀,不涂乙烯利刺激),用1.5%乙烯利在不同時間處理橡膠樹,相應4個時間點分別為0、3、12、24 h,具體實驗參照相關文獻中的方法進行[8]。
網上測序數據來自NCBI SRA數據庫,包括不同組織(PRJNA201084),真菌侵染(Corynespora cassiicola tolerance, PRJNA179126,葉片)、高低溫干旱脅迫(PRJNA182078,葉片)和乙烯利處理(PRJNA182079,膠乳),詳細信息參見數據庫中相關說明(http://www.ncbi.nlm.nih.gov/sra)。
1.2 方法
1.2.1 基因家族成員的鑒定與序列分析 為了全面鑒定橡膠樹、擬南芥、楊樹、水稻、木薯和蓖麻的PPCK基因家族成員,筆者從NCBI(http://www.ncbi.nlm.nih.gov/)和phytozome(https://phytozome.jgi.doe.gov/pz/portal.html)數據庫下載了6種植物的基因組和轉錄組數據。以已發表的PPCK序列為探針[3-5],對6種植物的基因組和轉錄組進行搜索,得到候選PPCK基因家族成員,利用InterProScan(http://www.ebi.ac.uk/Tools/pfa/iprscan/)對候選基因的結構域進行分析。再利用EMBOSS數據庫的在線軟件Pepstats(http://www.ebi.ac.uk/Tools/seqstats/emboss_pepstats/)對基因的等電點和分子量進行分析。
1.2.2 基因結構與進化分析 利用在線軟件GSDS2.0(http://gsds.cbi.pku.edu.cn/)對橡膠樹和其他5種植物PPCK基因家族成員的外顯子/內含子組織結構進行分析。利用MEGA 6.0軟件對橡膠樹和其他5種植物的PPCK氨基酸序列一起構建系統發育樹,采用Neighbor-Joining方法進行分子系統學分析,進行1 000次bootstrap統計學檢驗。
1.2.3 基因的表達模式分析 利用本實驗室和NCBI的solexa轉錄組數據對橡膠樹PPCK基因家族成員的表達模式進行分析[9]。首先,將NCBI的SRA數據庫(http://www.ncbi.nlm.nih.gov/sra/)中橡膠樹相關的solexa轉錄組數據下載到本地服務器,去除低質量序列,利用程序RSEM進行表達分析[10]。
2 結果與分析
2.1 PPCK基因家族成員的鑒定與序列分析
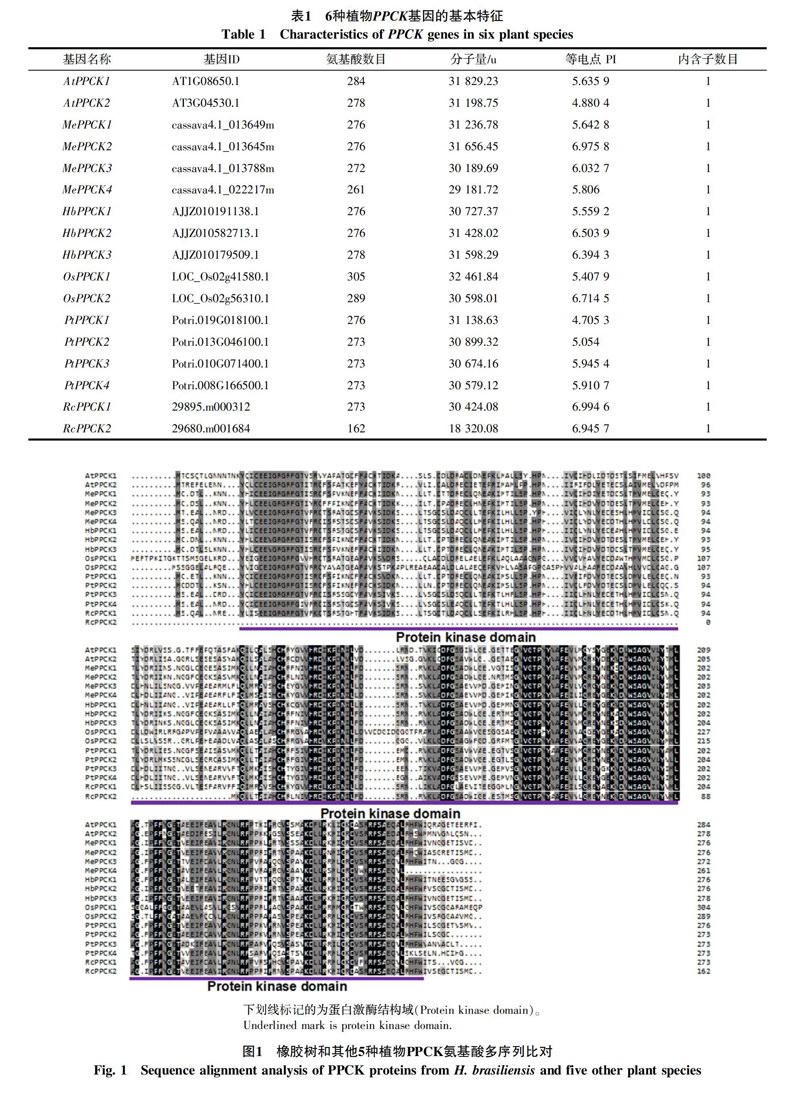
利用tblastn搜索橡膠樹、木薯、蓖麻、擬南芥、楊樹和水稻6種植物的轉錄組和基因組數據,鑒定得到17個基因家族(表1),其中包括3個橡膠樹基因家族,命名為HbPPCK1-3。通過序列分析發現,橡膠樹PPCK蛋白為276~278個氨基酸,分子量為31~32 ku,等電點在5.6~6.5之間,內含子數目均為1。氨基酸序列多序列比對發現,橡膠樹PPCK蛋白都含有蛋白激酶保守結構域(圖1)。
2.2 基因結構和進化分析
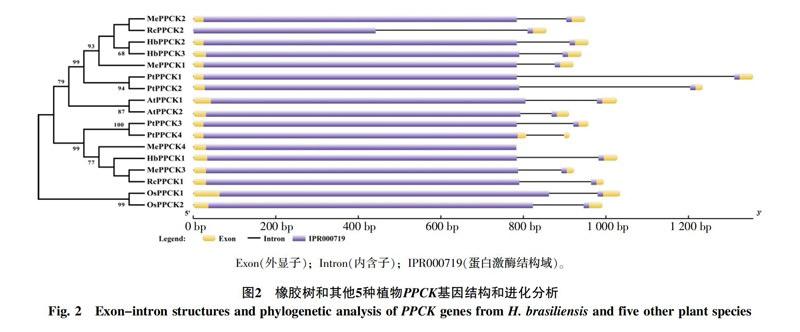
基因結構和進化分析發現(圖2),6種植物基因家族成員在內含子數目和相對位置,以及結構域的相對位置上都非常一致,均僅有1個內含子和蛋白激酶結構域。在進化上,單子葉植物和雙子葉植物分別聚在一起,而雙子葉植物又分出2個分支,其中HbPPCK1位于一個分支,HbPPCK2和HbPPCK3位于一個分支。有部分物種存在同源蛋白對,如HbPPCK2和HbPPCK3、AtPPCK1和AtPPCK2、PtPPCK3和 PtPPCK4等,說明這些同源對具有較近的親緣關系。橡膠樹基因家族成員主要和木薯、蓖麻2種大戟科植物PPCK聚在一起,這3種植物具有較近的親緣關系一致。
2.3 橡膠樹PPCK基因家族成員的表達分析
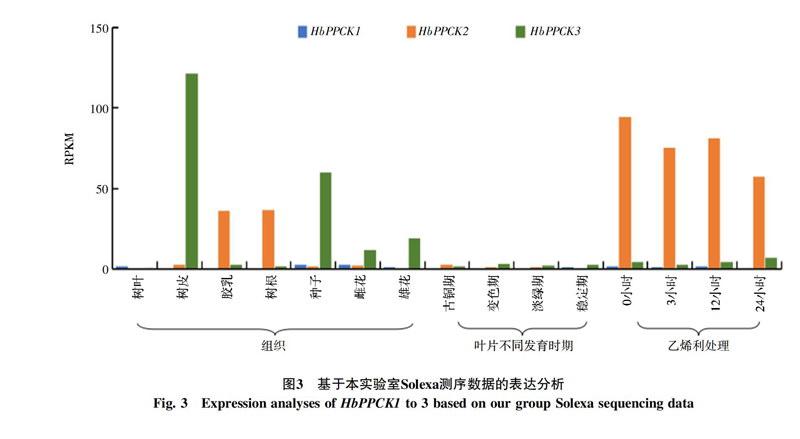
利用本實驗室現有的Solexa測序數據對3個橡膠樹PPCK基因在不同組織、不同葉片發育時期和乙烯利處理下的表達模式進行分析。結果表明:HbPPCK3在樹皮中表達豐度最高,其次是種子和雄花;HbPPCK2主要在膠乳和根中表達,在其他組織中的表達偏低,隨乙烯利刺激時間加長,表達呈下降趨勢。HbPPCK1在所檢測的所有組織中表達都很低(圖3)。另外,各基因家族成員在葉片中的表達豐度相對其他組織都非常低,在葉片發育過程中也沒有明顯變化趨勢。
筆者還利用NCBI的SRA數據庫中的轉錄組數據對橡膠樹PPCK的表達情況進行了分析。分析結果表明,HbPPCK3同樣在樹皮中表達豐度最高,C. cassiicola侵染、干旱、低溫和乙烯利刺激都會不同程度的誘導HbPPCK3表達;HbPPCK2主要在膠乳中表達,低溫處理后其表達豐度都會升高,而乙烯利刺激后表達豐度降低與本實驗室數據結果一致;HbPPCK1在各組織中表達都很低,C. cassiicola侵染和低溫處理會誘導其表達。
從以上兩部分實驗數據的結果來看,一個明顯的特征就是HbPPCK3在樹皮中高豐度表達,并且表達豐度要遠遠高于其他基因或其他組織,并且各種脅迫處理都會不同程度的誘導該基因的表達。這說明HbPPCK3可能在橡膠樹響應逆境脅迫應答方面具有重要功能。另外,NCBI和本實驗室數據結果的一致性,說明了本實驗分析結果的準確性。
3 討論
PEPC是C4光合作用途徑中最重要的酶之一,PPCK通過磷酸化調節PEPC活性。本研究充分利用網上數據庫資源,從橡膠樹、木薯、水稻、楊樹、蓖麻和擬南芥6種植物的全基因組數據中鑒定出17個PPCK基因家族成員,并從基因結構、進化和表達水平等方面進行了系統分析。在基因結構方面,和CDPK-SnRK超家族的其他成員相比,PPCK較為簡單,僅有一個蛋白酶結構域(圖2),除去激酶結構域以外的編碼區很短。相對CDPK等其他CDPK-SnRK成員,PPCK缺少鈣依賴型蛋白激酶所特有的自抑區(auto-inhibitory)和EF-hands 結構域,所以PPCK是一種非鈣依賴型蛋白激酶[2-3]。在進化方面,PPCK主要分為2個較大的分支,單子葉植物和雙子葉植物分別聚在一起,雙子葉植物又分出2個分支,其中HbPPCK2和 HbPPCK3位于一個分支,HbPPCK1位于一個分支,結合表達分析的結果可以看出,HbPPCK1在各組織中表達豐度都比較低,而HbPPCK2和HbPPCK3分別在不同組中高豐度表達,另外擬南芥的2個PPCK成員也和HbPPCK2、HbPPCK3位于同一分支,因而可以初步推測HbPPCK2和HbPPCK3所處的分支為PPCK進化的主要方向。已有研究認為,PPCK基因的表達主要通過轉錄水平進行調控[3-5],在本研究中HbPPCK1在各組織中表達豐度都比較低或不表達,HbPPCK2和HbPPCK3的表達具有一定的組織特異性,并且受不同脅迫處理的調控,如HbPPCK3主要在樹皮和種子中表達,真菌侵染、干旱、低溫和乙烯利刺激都會不同程度的誘導其表達,HbPPCK2主要在膠乳中表達,低溫處理可以誘導其在葉片中的表達,而乙烯利處理會降低其在膠乳中的表達,由此推測逆境條件能誘導PPCK基因的表達,進而對磷酸烯醇式丙酮酸羧化酶的活性進行調節。本實驗室的高通量數據已成功應用于橡膠樹基因組數據分析以及橡膠樹蔗糖合成酶基因家族相關分析,相關結果已發表在Nature Plants[11]和 Febs Journal[9],這進一步證實了本文數據結果的可靠性。本研究利用相關軟件和數據庫,對橡膠樹和其他植物PPCK基因家族成員的結構、進化和表達進行了初步分析,研究結果將為進一步研究PPCK基因在橡膠樹光合作用和逆境脅迫應答等方面的功能奠定基礎。
參考文獻
[1] Hatch M D. C 4 photosynthesis: a unique elend of modified biochemistry, anatomy and ultrastructure[J]. Biochimica et Biophysica Acta(BBA)-Reviews on Bioenergetics, 1987, 895(2): 81-106.
[2] Hrabak E M, Chan C W M, Michael G, et al. The Arabidopsis CDPK-SnRK superfamily of protein kinases.[J]. Plant Physiology, 2003, 132(2): 666-680.
[3] Hartwell J, Gill A, Nimmo G A, et al. Phosphoenolpyruvate carboxylase kinase is a novel protein kinase regulated at the level of expression[J]. Plant Journal for Cell & Molecular Biology, 1999, 20(3): 333-42.
[4] Shenton M. Distinct patterns of control and expression amongst members of the PEP carboxylase kinase gene family in C 4 plants[J]. Plant Journal, 2006, 48(1): 45-53.
[5] Fontaine V, Hartwell J, Jenkins G I, et al. Arabidopsis thaliana contains two phosphoenolpyruvate carboxylase kinase genes with different expression patterns[J]. Plant Cell & Environment, 2002, 25(1): 115-122.
[6] d'Auzac J, Jacob J L, Prévot J C, et al. The regulation of cis-polyisoprene production(natural rubber)from Hevea brasiliensis, In Recent research developments in plant physiology(ed S.G. Pandalai)[R]. Research Singpost, Trivandrum, India. 1997: 273-332.
[7] Hao B Z, Wu J L. Laticifer Differentiation in Hevea brasiliensis: Induction by Exogenous Jasmonic Acid and Linolenic Acid[J]. Annals of Botany, 2000, 85(1): 37-43.
[8] 肖小虎. 巴西橡膠樹蔗糖代謝相關基因家族的克隆、 結構進化和表達分析[D]. 海南大學, 2013.
[9] Xiao X H, Tang C R, Fang Y, et al. Structure and expression profile of the sucrose synthase gene family in the rubber tree: indicative of roles in stress response and sucrose utilization in the laticifers[J]. Febs Journal, 2014, 281(1): 291-305.
[10] Li B, Dewey C N. Li B, etal. RSEM: accurate transcript quantification from RNA-Seq data with or without a reference genome [J]. BMC Bioinformatics, 2011, 12(1): 93-99.
[11] Tang C, Meng Y, Fang Y, et al. The rubber tree genome reveals new insights into rubber production and species adaptation[J]. Nature Plants, 2016, 2: 16073.
