文蛤C型凝集素基因(Mm-Lec1)的克隆與表達分析
2016-07-01 06:09:56張晶晶李宏俊秦艷杰劉敏葉晟
海洋學報 2016年6期
張晶晶,李宏俊, 秦艷杰, 劉敏,葉晟
(1. 大連海洋大學 水產與生命學院,遼寧 大連 116023;2. 國家海洋環境監測中心 海洋生態室,遼寧 大連 116023)
文蛤C型凝集素基因(Mm-Lec1)的克隆與表達分析
張晶晶1,2,李宏俊2*, 秦艷杰1, 劉敏1,2,葉晟1,2
(1. 大連海洋大學 水產與生命學院,遼寧 大連 116023;2. 國家海洋環境監測中心 海洋生態室,遼寧 大連 116023)
摘要:文蛤(Meretrix meretrix)是我國重要的灘涂養殖貝類,病害嚴重影響文蛤增養殖業,研究文蛤的免疫機制有助于解決文蛤的病害問題。C型凝集素(C-type lectin)參與先天免疫,在識別病原相關分子模式和激活體液免疫因子等方面發揮重要作用。本研究檢索已構建的文蛤全長cDNA 文庫,經過Blast比對得到了文蛤C型凝集素1(Mm-Lec1)基因的全長cDNA序列。Mm-Lec1序列全長586 bp,5`和3`非翻譯區(UTR)的長度分別為21 bp和79 bp,開放閱讀框長度為486 bp,編碼161個氨基酸,分子量為18.65 kD,理論等電點為4.98。預測的氨基酸序列中含有信號肽(Met1-Ser19)、糖識別結構域(CRD)和糖結合位點(QPN)。Mm-Lec1的三級結構是緊湊型,含有β片層結構。同源性分析結果表明,Mm-Lec1與其他物種C型凝集素相似度在20%~32%;鄰接法(Neighbor-Joining, NJ)進化樹分析結果表明,Mm-Lec1與紫貽貝CTL 6和櫛孔扇貝CTL A聚為一支。實時熒光定量分析結果顯示,Mm-Lec1在文蛤鰓、肝胰腺、閉殼肌、外套膜、性腺和血細胞中均表達,其中鰓表達量最高,血細胞次之,性腺中表達量最少;在鰻弧菌(Vibrio anguillarum)刺激實驗中,6 h時Mm-Lec1在血細胞中的表達量最低,48 h表達量最高,暗示Mm-Lec1參與文蛤抵御細菌入侵的免疫過程。
關鍵詞:文蛤;C型凝集素;表達序列標簽;三級結構;實時熒光定量PCR
1引言
文蛤(Meretrixmeretrix)主要分布于廣西北部灣、江蘇南部、山東蓬萊灣及遼寧遼河口等海區[1],是我國重要的灘涂經濟貝類,也是主要的出口鮮活水產品之一。近年來,病害問題嚴重制約我國文蛤增養殖業發展,文蛤病害暴發具有明顯的區域性、季節性和流行性[2],常常導致文蛤大規模死亡,成為文蛤養殖業遇到的最大難題[3—4]。導致貝類病害暴發的原因是多方面的,是外界病原、環境和貝類生物自身共同作用的結果。研究文蛤抵御病害機制,探討貝類自身免疫防御機理,可以為文蛤抗病選育提供理論基礎。
C型凝集素(C-type lectin, CTL)代表一個依賴于鈣離子(Ca2+)參與的與糖原結合的蛋白家族,含有1個或多個一級結構和二級結構同源的碳水化合物識別結構域。C型凝集素根據與其結合的配體不同可分為Man型配體和Gal型配體,前者與D-甘露糖、D-葡萄糖結合,在氨基酸序列中對應的三氨基分別是EPN和EPD;后者與D-半乳糖及其衍生物結合,在氨基酸中對應的三氨基分別是QPD和QPN[5—6]。在先天免疫應答中,C型凝集素配體與外來病原菌糖原結合,中和病原菌的致病毒性,通過補體途徑或者細胞吞噬途徑消滅病原菌[7]。CTL在貝類先天免疫中發揮重要作用,在櫛孔扇貝(Chlamysfarreri)[5],合浦珠母貝(Pinctadafucata)[8],海灣扇貝(Argopectenirradias)[9]和偏頂蛤(Modiolusmodiolus)[10]抵御細菌,海灣扇貝損傷愈合[9],凡納濱對蝦(Litopenaeusvannamei)[11]和斑節對蝦(Penaeusmonodon)[12]抵御病毒入侵和文蛤(M.meretrix)耐鹽和耐高溫[13]生理響應中發揮作用。
目前,已報道CTL基因的軟體動物包括紫貽貝(Mytilusgalloprovincialis)[14],櫛孔扇貝[15—16],海灣扇貝[9,17—18]和合浦珠母貝(P.fucata)[8,19],太平洋牡蠣(Crassostreagigas)[20—21]、美洲牡蠣(Crassostreavirginica)[22]、和皺紋盤鮑(Haliotisdiscus)[23—24]等。本研究克隆了文蛤Mm-Lec1基因的全長cDNA序列,分析了Mm-Lec1基因mRNA在各組織分布情況和病原菌刺激下的表達規律,這將為進一步研究CTL基因在軟體動物中的免疫功能提供基礎資料。
2材料與方法
2.1材料
試驗用文蛤采自遼寧省盤山縣遼河口,在室內的60 L的塑料箱中暫養2周后,選取活力較好、殼長均勻(4.0~5.0 cm)的文蛤進行實驗。對于組織表達實驗,選取3只閉殼肌活力強的文蛤分別取血細胞、鰓、閉殼肌、性腺、外套膜和肝胰腺組織,保存于液氮中備用。對于鰻弧菌(virbrioanguillarum)浸染實驗,設置對照組和實驗組,實驗組海水鰻弧菌終濃度為4×107cfu/mL,在刺激0 h、6 h、12 h、24 h、48 h、72 h和96 h用1 mL注射器采集閉殼肌的血細胞,每個時間點設置3個重復,經4℃,5 500 r/min離心10 min處理后,保存于液氮中備用。
2.2文蛤C 型凝集素基因序列的獲得
本課題組已構建了文蛤3個組織的全長cDNA文庫,共得到3 129條高質量的EST序列[25—26],在NCBI (http://www.ncbi.nlm.nih.gov/)網站利用tblastx進行同源序列搜索比對,獲得文蛤C型凝集素的全長cDNA序列。
2.3文蛤C型凝集素基因序列的生物信息學分析
應用ORF Finder(http://www.ncbi.nlm.nih.gov/gorf/gorf.html)程序確定正確的開放閱讀框,并運用BioEdit軟件翻譯成氨基酸序列;采用InterProScan 5(http://www.ebi.ac.uk/Tools/pfa/iprscan5/)預測氨基酸序列中的疏水區;采用TMHMM在線服務器(http://www.cbs.dtu.dk/services/TMHMM/)預測蛋白跨膜區;用SignalP4.1 Server軟件 (http://www.cbs.dtu.dk/services/SignalP/)預測信號肽;采用SMART(http://smart.embl-heidelberg.de/)在線服務軟件預測功能域;采用DiANNA(http://clavius.bc.edu/~clotelab/DiANNA/)預測二硫鍵的個數;采用ExPASy SWISSMODEL軟件 (http://swissmodel.expasy.org/interactive)預測空間結構,采用RasTop軟件查看模型;運用在線軟件TargetP 1.1(http://www.cbs.dtu.dk/services/TargetP/)對蛋白質進行定位;通過NCBI (http://www.ncbi.nlm.gov)中的tblastx工具進行蛋白質序列相似性檢索;用ClustalX 2.1軟件與GeneDoc軟件聯合進行多序列比對;應用ClustalX 2.1軟件與MEGA 5.0軟件構建NJ系統進化樹;用在線ClustalW 2服務器(http://www.ebi.ac.uk/Tools/msa/clustalw2/)分析蛋白質序列的相似性。
2.4文蛤C型凝集素的組織分布表達與時序表達
按照天根的提取RNA的試劑盒說明書提取RNA,采用生工的反轉錄試劑盒獲得cDNA。依照寶生物的SYBR Premix Ex Taq kit試劑盒說明書,以cDNA為模板,配置20 μL的擴增體系,同時設置ABI7500系統的擴增程序。為了確保引物的特異性,我們增加了溶解曲線的擴增。為了減少實驗的誤差,每個組織或時間點重復3次實驗。利用Primer Premier 5.0設計C型凝集素和β-actin的特異性引物(表1)。采用2-△△Ct法計算Mm-Lec1基因的相對表達量,以β-actin為內參基因,使用One way ANOVA分析實驗組與對照組的Mm-Lec1基因的相對表達量的顯著性差異。
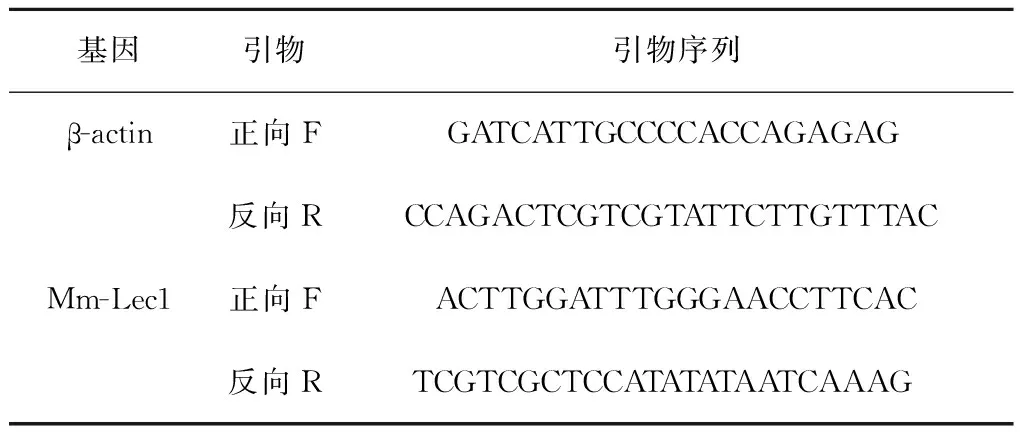
表1 β-actin和Mm-Lec1的特異性引物
3結果
3.1Mm-Lec1序列分析
Mm-Lec1基因的全長cDNA序列為586 bp(GenBank注冊號KT276515),3′端有polyA尾巴,5′和3′非翻譯區(UTR)的長度分別為21 bp和79 bp,開放閱讀框長度為486 bp,編碼161個氨基酸。Mm-Lec1氨基酸序列Met1-Ser19為信號肽序列,Ser19和Ala20之間存在剪切位點(圖1)。氨基酸序列包含1個糖識別結構域(CRD),CRD包含6個保守的半胱氨酸(Cys21,Cys32,Cys49,Cys125,Cys141和Cyst150)和1個糖結合位點QPN(Gln115,Pro116,Asn117)(圖1)。按照標準密碼子翻譯后的蛋白質推測分子式為C841H1227N217O245S11,分子量計算值為18.65 kD,理論等電點為4.98。極性與極性帶電氨基酸共99個(占61.5%),非極性氨基酸62個(占38.5%),極性氨基酸比較大,決定了Mm-Lec1氨基酸具有較好的水溶性。
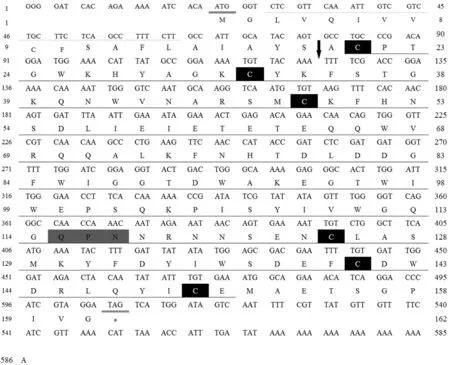
圖1 Mm-Lec1基因的核苷酸序列和推測的氨基酸序列Fig.1 The full cDNA and predicted amino acid sequence of Mm-Lec1 gene雙下劃線代表起始密碼子和終止密碼子;帶有下劃線的氨基酸序列是C型凝集素家族糖識別結構域(CRD)序列;帶有虛線下劃線的氨基酸是疏水氨基酸;黑色背景表示形成二硫鍵的半胱氨酸;灰色陰影代表糖結合位點;↓代表信號肽剪切位點The start and stop codons are double line underlined; the carbohydrate recognition domain (CRD) is underlined; hydrophobic amino acids are dotted line underlined; cysteines constituting disulfide bonds are in black box; the sugar binding sites are in grey shadow re-gion; ↓ is the token of signal peptide splice site
3.2蛋白亞細胞定位分析
TMHMM預測前60個氨基酸含有疏水氨基酸,存在跨膜區氨基酸的預測值較高(14.5>10),跨膜螺旋結構的預測值低(14.5<18)。InterProScan 5估測Leu3-Leu14為疏水區。TargetP 1.1在線服務器估算Mm-Lec1是分泌蛋白的概率是93.4%。
3.3同源比對、相似度和系統樹
從23條C型凝集素基因比對圖(圖2)發現6個位點的氨基酸是完全一樣的,其中5個是半胱氨酸。ClustalW 2在線服務器計算得出文蛤與紫貽貝C-type lectin 6的相似度最大(32%),其次是紫貽貝C-type lectin 4(31%),與斑節對蝦的相似度最小(20%)。利用MEGA 5.0軟件建立鄰接法(Neighbour-Joining,NJ)系統樹(圖3)。結果顯示Mm-Lec1先和紫貽貝(AJQ21497)聚為一支,然后和櫛孔扇貝(DQ209289)聚為一大支,和其他的軟體動物的C型凝集素的進化距離較遠。
3.4蛋白高級結構同源建模分析
運用DiANNA在線軟件,得出CRD結構域的6個半胱氨酸形成3個二硫鍵(分別是1-12:XXXXXCPSGWI-EFNQECFLFGA,29-130:DAEADCRRHSS-RYNYICEXXXX,104-122:NNSANCMSFYM-WRDDRCTARYN),連接方式是第1個和第2個,第3個和第6個,第4個和第5個半胱氨酸形成二硫鍵。將Mm-Lec1氨基酸序列提交到(http://swissmodel.expasy.org/)網站,在RCSB PCB數據庫(http://www.rcsb.org/pdb/home/home.do)中比對,得到1個與之相似度(similarity:37.50%)比較高的氨基酸序列的高級結構模板:小鼠清道夫受體C型凝集素Collectin placenta 1(2ox9.3.A)[27],其全局模型質量評估(Global Model Quality Estimation,GMQE)值為0.63(GMQE值在0~1之間,值越大,模板與目標基因的匹配度越高,得到的蛋白質的三級結構越可信)。根據此模板建立Mm-Lec1高級結構模型(圖4A),結果顯示蛋白分子空間結構是緊湊型,是由6個Helices,19個Strands,和11個Turns,其中3條β折疊形成1個β片層結構(圖4C)。
3.5文蛤Mm-Lec1組織表達規律
3.5.1C型凝集素在文蛤各個組織中的分布情況
采用QRT-PCR,我們分析了Mm-Lec1基因在健康文蛤各個組織中的相對表達量(圖5),鰓中Mm-Lec1的mRNA表達量是血細胞的2.6倍(P>0.05),閉殼肌中的表達量是血細胞的0.8倍(P>0.05),外套膜中的表達量是血細胞的0.3倍(P>0.05),肝胰腺中的表達量是血細胞的0.2倍(P<0.05),性腺中的表達量是血細胞0.09倍(P<0.05),所以鰓中表達量最高,血細胞次之,性腺中表達量最低。
3.5.2菌刺激之后Mm-Lec1的時間表達分析
在細菌刺激試驗中,我們分析了鰻弧菌(Vibrioanguillarum)刺激后0 h,6 h,12 h,24 h,48 h,72 h和96 h文蛤血細胞C型凝集素基因mRNA的相對表達量(圖6),6 h的表達量是0 h的0.4倍(P<0.05),12和24 h的表達量分別是0 h的1.1倍和1.9倍,48 h的相對表達量是0 h的3.3倍(P<0.05),72 h后Mm-Lec1的mRNA表達量恢復到0 h水平。C型凝集素的表達在時間上呈現先降低,再升高,在48 h時達到峰值,而后降低到基礎水平的趨勢。
4討論
文蛤是我國沿海常見的一種重要經濟貝類,也是出口創匯的重要水產品之一,但是由于病害流行和養殖水域污染,文蛤養殖業受到極大損失,因此研究文蛤免疫機制顯得尤為重要。本研究報道了文蛤重要免疫基因C型凝集素Mm-Lec1的cDNA序列,運用InterProScan 5推測Mm-Lec1氨基酸序列Leu3-Leu14為疏水區,猜測這段氨基酸序列存在信號肽或者跨膜區。由TMHMM預測前60個氨基酸含有疏水氨基酸,存在跨膜區的氨基酸預測值相對比較高(14.5>10),跨膜螺旋結構的預測值低(14.5<18),推測Mm-Lec1中不含有跨膜螺旋結構,可能含有1個N端信號肽序列[28]。利用軟件SignalP4.0發現文蛤Mm-Lec1氨基酸序列Met1-Ser19是信號肽序列,TargetP 1.1預測得出C型凝集素是分泌蛋白的概率是93.4%,猜測文蛤Mm-Lec1是分泌蛋白。Mm-Lec1氨基酸序列的糖結合位點是QPN,具有D-半乳糖結合特性,說明Mm-Lec1是Gal型配體。從多序列比對圖中我們發現,糖結合位點是EPN和QPD的物種數最多,EPD其次,QPN、VPN、YPD和EPS最少。在糖結合位點中,EPN和QPD是高度保守的,其他都是突變型,這種高度保守的堿基被替換說明海洋生物C型凝集素序列的多樣性[16]。
用Swissmodel構建的Mm-Lec1模型與小鼠清道夫受體C型凝集素Collectin placenta 1(2ox9.3.A)的同源性較高,序列的氨基酸組成分析得出Mm-Lec1是水溶性蛋白質,推測Mm-Lec1屬于Collectin家族中的一員。Mm-Lec1的高級結構是緊湊型,運用DiANNA預測合浦珠母貝[8,19]和海灣扇貝[9]的CRD結構中的二硫鍵發現,都含有3個二硫鍵,半胱氨酸的結合方式都是第1個和第2個,第3個和第6個,第4個和第5個結合,但是合浦珠母貝是球形而海灣扇貝是長型(long-form),說明蛋白質的高級結構不僅僅只與二硫鍵的數量有關,但還需要更多的研究進行佐證。
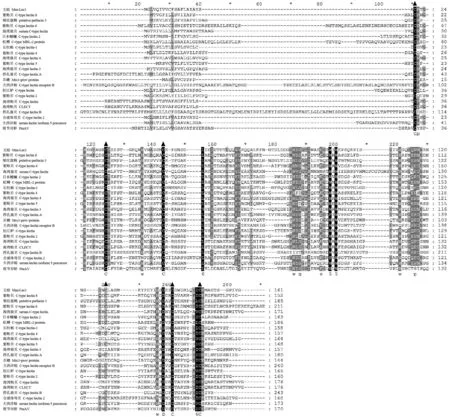
圖2 文蛤C型凝集素與其他已知的C型凝集素的多序列比對Fig.2 Multiple alignments of Mm-Lec1 with other known C-type lectin sequences黑色背景代表同源性,灰色陰影代表相似性;▲表示糖識別結構域中形成二硫鍵的半胱氨酸。其中文蛤Mm-Lec1 (M. meretrix, KT276515);紫貽貝C-type lectin 8(M. galloprovincialis, AJQ21499);皺紋盤鮑putative perlucin 3 (H. discus discus, ABO26595);紫貽貝C-type lectin 6(M. galloprovincialis, AJQ21497);海灣扇貝 serum C-type lectin (A. irradians, ACE80702);日本鰻鱺 C-type lectin 2(A. japonica, BAC54021);虹鱒 C-type MBL-2 protein(O. mykiss, CAJ14130);長牡蠣 C-type lectin-1(C. gigas, BAF75353);紫貽貝C-type lectin 4(M. galloprovincialis, AJQ21495);海灣扇貝 C-type lectin 4(A. irradians, ACS72239);紫貽貝C-type lectin 5(M. galloprovincialis, AJQ21496);海灣扇貝 C-type lectin 3(A. irradians, ACS72238);櫛孔扇貝C-type lectin A(C. farreri, DQ209289);爪蟾 Mrc1-prov protein(X. laevis, AAH77648);大西洋鮭 C type lectin receptor B(S. salar, AAT77221 );松江鱸 C-type lectin(T. fasciatus, AFW17073);紫貽貝C-type lectin 2(M. galloprovincialis, AJQ21493);海灣扇貝 C-type lectin(A. irradians, EU277646);海灣扇貝CLECT(A. irradians, ABZ89710);櫛孔扇貝 C-type lectin B(C. farreri, DQ209290);合浦珠母貝C-type lectin 2(P. fucata, ACO36046);大西洋鮭 serum lectin isoform 5 precursor(S. salar, AAO43604);斑節對蝦PmAV(P. monodon, AAQ75589)Homology are in the black region; similarity are in the grey region; the acids under ▲ are cysteines constituting disulfide bonds in the structure domain. There include hard clam Mm-Lec1 (M. meretrix, KT276515), blue mussel C-type lectin 8 (M. galloprovincialis, AJQ21499), disk abalone putative perlucin 3 (H. discus discus, ABO26595), blue mussel C-type lectin 6 (M. galloprovincialis, AJQ21497), bay scallop serum C-type lectin (A. irradians, ACE80702), Japanese eel C-type lectin 2 (A. japonica, BAC54021), rainbow trout C-type MBL-2 protein (O. mykiss, CAJ14130), Pacific oyster C-type lectin-1 (C. gigas, BAF75353), blue mussel C-type lectin 4 (M. galloprovincialis, AJQ21495), bay scallop C-type lectin 4 (A. irradians, ACS72239), blue mussel C-type lectin 5 (M. galloprovincialis, AJQ21496), bay scallop C-type lectin 3 (A. irradians, ACS72238), Zhikong scallop C-type lectin A (C. farreri, DQ209289), clawed toad Mrc1-prov protein (X. laevis, AAH77648), Atlantic salmon C type lectin receptor B (S. salar, AAT77221 ), roughskin sculpin C-type lectin (T. fasciatus, AFW17073), blue mussel C-type lectin 2 (M. galloprovincialis, AJQ21493), bay scallop C-type lectin (A. irradians, EU277646), bay scallop CLECT (A. irradians, ABZ89710 ), Zhikong scallop C-type lectin B (C. farreri, DQ209290), Peal oyster C-type lectin 2 (P. fucata, ACO36046), Atlantic salmon serum lectin isoform 5 precursor (S. salar, AAO43604), black tiger shrimp PmAV (P. monodon, AAQ75589)
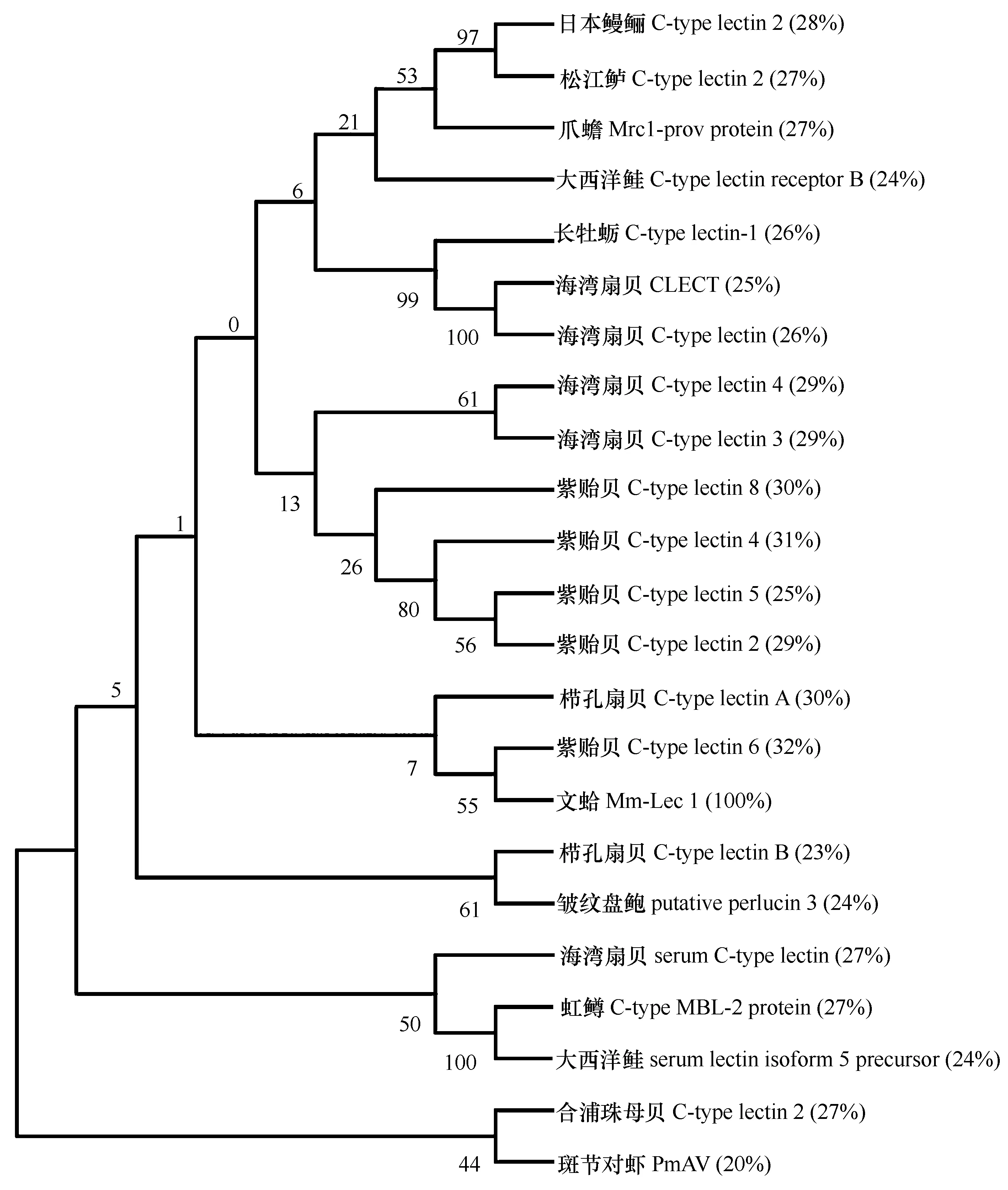
圖3 23條C型凝集素基因構建的NJ進化樹以及Mm-Lec1與其他已知CTL基因的相似度Fig.3 The NJ tree of 23 CTL genes and the similarity between other known CTL and Mm-Lec1

圖4 文蛤C型凝集素蛋白的高級結構不同角度觀測圖以及β片層結構展示圖Fig.4 The tertiary structure in different views of Mm-Lec1 and the show of β lamellar structureA.正視圖;B.仰視圖; C.在B基礎上Z軸旋轉逆時針旋轉60°得到的側視圖。藍色部分是β轉角,綠色部分是無規卷曲,黃色部分是β折疊,紅色部分是α螺旋A. Front view; B. Vertical view; C. Side view generated after B rotates 60 degrees CCW on Z axis. The blue, green, yellow and red is the token of β-turn, random coil, β folding and α helix, respectively
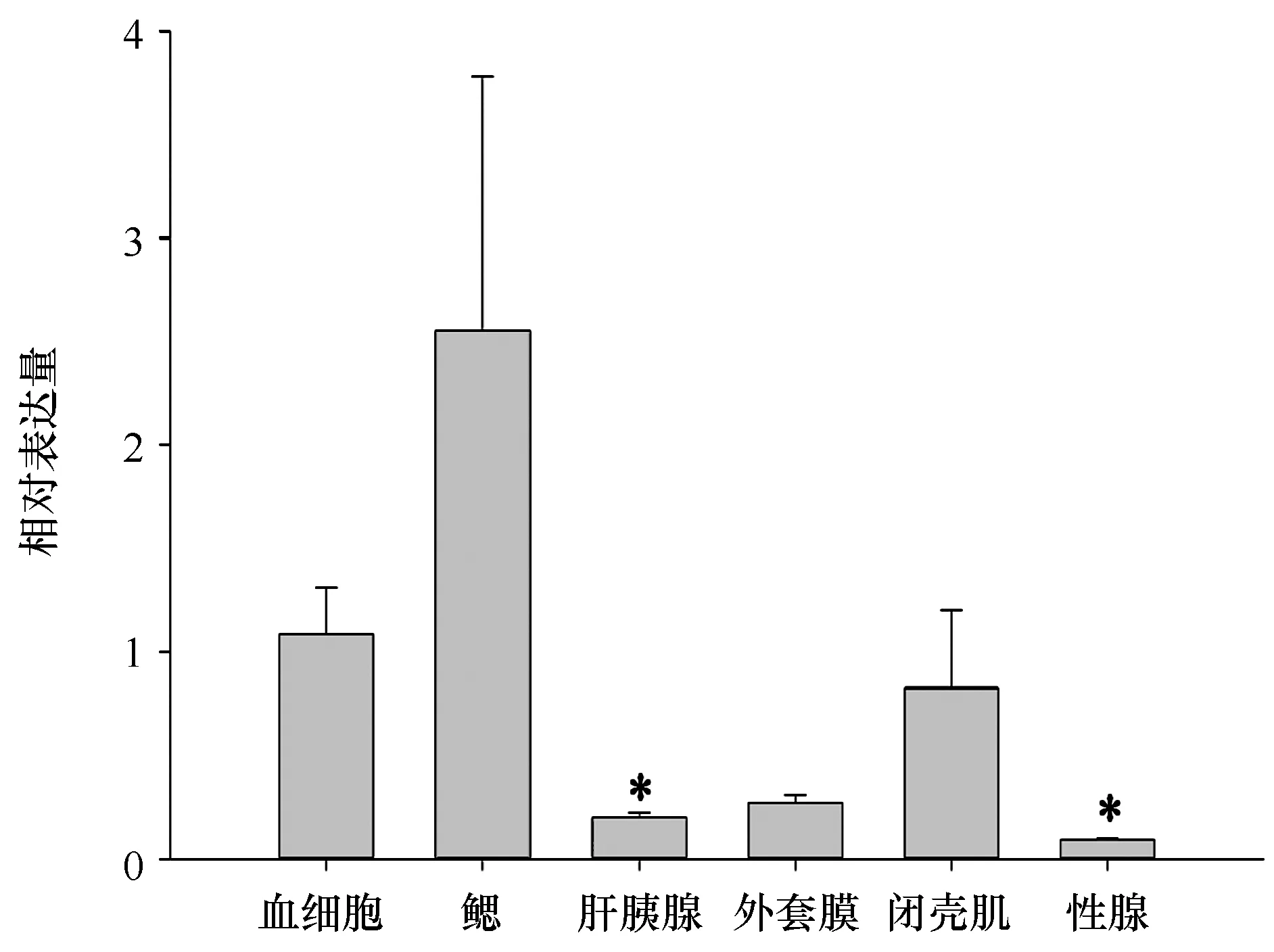
圖5 文蛤C型凝集素在組織中的表達量Fig.5 Relative expression of the Mm-Lec1 mRNA in tissues. Bars display mean ± S. D.*代表顯著性差異(P<0.05),柱狀圖代表的是相對表達量的平均值±方差* is the token of significance difference (P<0.05)

圖6 菌刺激后文蛤血細胞中C型凝集素基因mRNA的表達量Fig.6 Relative expression of the Mm-Lec1 gene mRNA in haemocytes after bacteria exposure*代表顯著性差異(P<0.05),柱狀圖代表的是相對表達量的平均值±方差* indicates significance difference (P<0.05). Bars display mean ± S. D.
一般低等生物中的C型凝集素只含有1個CRD,其他物種的則有兩個或多個CRD[29]。文蛤Mm-Lec1氨基酸含有1個C型凝集素結構域,這和海灣扇貝(A.irradias)[9,17—18,29]以及合浦珠母貝(P.fucata)[8,19]的C型凝集素結構一致,但和櫛孔扇貝(C.farreri)[15—16,30—31]結構不一致,其中Cflec-3[31]和Cflec-4[30]分別含有3個和4個CRD結構域。從23條序列的比對圖發現,比軟體動物在進化上高級很多的兩棲類爪蟾和魚類,都含有1個CRD結構域的情況。Mm-Lec1與其他物種的CTL的相似度在20%~32%之間(圖3),文蛤和紫貽貝 C-type lectin 6的相似度最高(32%),其次是紫貽貝C-type lectin 4(31%),與斑節對蝦PmAV的相似度最低(20%),與非同類的日本鰻鱺(28%),松江鱸(27%),爪蟾(27%)和虹鱒C-type MBL-2 protein(27%)卻有相對較高的相似度;此外,Mm-Lec1基因和最近發表的Mm-CTL基因堿基組成有很大的差別,相似度僅是36.5%。同時,NJ系統樹結果分析顯示不同物種及同一物種的CTL基因并沒有產生特別明顯的系統分化關系,比如,同屬于海灣扇貝的CTL基因和同屬紫貽貝的CTL基因并沒有各自獨立分為一支,甚至有一些基因的進化距離相對較遠;屬于兩棲類的爪蟾的C型凝集素基因卻與魚類的聚為一支;與Mm-Lec1同源性相差不多的紫貽貝 C-type lectin 6和紫貽貝C-type lectin 4卻與Mm-Lec1有不同進化距離。這些進化關系可能是由于凝集素基因家族成員不同的進化路徑和不同的功能造成的,也證實了C型凝集素是一個具有保守序列并且進化較快的基因[8]。
文蛤Mm-Lec1 mRNA在所檢測組織中都有表達,其中鰓中表達量最高,血細胞次之,性腺中表達量最少。胡鈺婷等[8]發現PoLEC1和PoLEC2在合浦珠母貝肝胰腺中表達量最高;Zhu等[9,29]發現AiCTL1和Ai Lec分別在海灣扇貝的血細胞和肝胰腺中表達量最高。鰓是軟體動物免疫系統的“第一道防線”,在貝類免疫方面起到重要的作用;血細胞和肝胰腺是無脊椎動物重要的免疫器官。但是在相同物種或不同物種不同的組織中C型凝集素基因的表達量不同,說明C型凝集素的種類很多,推測是不同C型凝集素基因的具體功能不同造成的[9]。血細胞不僅可以直接隔離并且殺死病原菌而且可以合成和分泌一些在免疫中有作用的活性物質,所以我們選擇血細胞為在不同時間的鰻弧菌刺激下的C型凝集素的實驗材料。6 h血細胞中的C型凝集素的表達量達到最低值,48 h達到最高值,72 h回到基礎水平,暗示C型凝集素在文蛤體內存在誘導表達的恢復模式,這與櫛孔扇貝[5]和合浦珠母貝[8]的C型凝集素研究結果一致,說明文蛤的C型凝集素有參與體內免疫和機體防御。
參考文獻:
[1]莊啟謙. 中國動物志, 軟體動物門, 雙殼綱, 簾蛤科[M]. 北京: 科學出版社, 2001: 230.
Zhuang Qiqian. Fauna sinica, phylum mollusca, bivalvia, veneridae[M]. Beijing: Science Press, 2001: 230.
[2]王如才, 王昭萍. 海水貝類養殖學[M]. 青島: 中國海洋大學出版社, 2008: 396-408.
Wang Rucai, Wang Zhaoping. Marine shellfish culture[M]. Qingdao: Ocean University of China Press, 2008: 396-408.
[3]劉連生, 閆茂倉, 林志華, 等. 引起文蛤暴發性死亡病原菌的分離和鑒定[J]. 微生物學通報, 2009, 36(1): 71-77.
Liu Liansheng, Yan Maocang, Lin Zhihua, et al. Pathogens isolation and identification of eruptive epidemic disease inMeretrixmeretrix[J]. Microbiology, 2009, 36(1): 71-77.
[4]劉連生, 閆茂倉, 趙海泉, 等. 文蛤疾病學研究進展[J]. 水產科學, 2009, 28(4): 234-237.
Liu Liansheng, Yan Maocang, Zhao Haiquan, et al. Advancement of pathological researches on hard clamMeretrixmeretrix[J]. Fisheries Science, 2009, 28(4): 234-237.
[5]胥煒, 王昊, 宋林生, 等. 櫛孔扇貝C型凝集素基因的克隆與表達研究[J]. 高技術通訊, 2005, 15(1): 83-88.
Xu Wei, Wang Hao, Song Linsheng, et al. cDNA cloning and mRNA expression of C-type lectin from scallopChlamysFarreri[J]. Chinese High Technology Letters, 2005, 15(1): 83-88.
[6]Drickamer K. Engineering galactose-binding activity into a C-type mannose-binding protein[J]. Nature, 1992, 360(6400): 183-186.
[7]Weis W I, Taylor M E, Drickamer K. The C-type lectin superfamily in the immune system[J]. Immunological Reviews, 1998, 163(1): 19-34.
[8]胡鈺婷, 張殿昌, 崔淑歌, 等. 合浦珠母貝C-型凝集素基因的序列特征和功能分析[J]. 水產學報, 2011, 35(9): 1327-1336.
Hu Yuting, Zhang Dianchang, Cui Shuge, et al. Sequence features and functional analysis of the C-type lectin gene (PoLEC1) from pearl oysterPinctadafucata[J]. Journal of Fisheries of China, 2011, 35(9): 1327-1336.
[9]Zhu Ling, Song Linsheng, Xu Wei, et al. Molecular cloning and immune responsive expression of a novel C-type lectin gene from bay scallopArgopectenirradians[J]. Fish Shellfish Immunol, 2008, 25(3): 231-238.
[10]Tunkijjanukij S, Olafsen J A. Sialic acid-binding lectin with antibacterial activity from the horse mussel: further characterization and immunolocalization[J]. Dev Comp Immunol, 1998, 22(2): 139-150.
[11]Zhao Zhiying, Yin Zhixin, Xu Xiaopeng, et al. A novel C-type lectin from the shrimpLitopenaeusvannameipossesses anti-white spot syndrome virus activity[J]. J Virol, 2009, 83(1): 347-356.
[12]Luo Tian, Zhang Xiaobo, Shao Zongze, et al.PmAV, a novel gene involved in virus resistance of shrimpPenaeusmonodon[J]. Febs Letters, 2003, 551(1/3): 53-57.
[13]李猛, 周素明, 劉璐, 等. 文蛤(Meretrixmeretrix)C型凝集素基因的分子克隆及表達分析[J]. 海洋與湖沼, 2015, 46(5): 1186-1192.
Li Meng, Zhou Suming, Liu Lu, et al. Molecular clone and expression and expression of C-type lectin inMeretrixmeretrix[J]. Oceanlogia et Limnologia Sinica, 2015, 46(5): 1186-1192.
[14]Gerdol M, Venier P. An updated molecular basis for mussel immunity[J]. Fish Shellfish Immunol, 2015, 46(1): 17-38.
[15]黃海寧. 櫛孔扇貝C型凝集素基因的cDNA克隆和序列分析[J]. 福建水產, 2007(4): 6-10.
Huang Haining. cDNA cloning and sequence analysis on C-type lectin gene ofChlamysfarreri[J]. Journal of Fujian Fisheries, 2007(4): 6-10.
[16]Wang Hao, Song Linsheng, Li Chenghua, et al. Cloning and characterization of a novel C-type lectin from Zhikong scallopChlamysfarreri[J]. Mol Immunol, 2007, 44(5): 722-731.
[17]Zhang Huan, Song Xiaoyan, Wang Lingling, et al. AiCTL-6, a novel C-type lectin from bay scallopArgopectenirradianswith a long C-type lectin-like domain[J]. Fish Shellfish Immunol, 2011, 30(1): 17-26.
[18]宋小燕. 海灣扇貝凝集素基因和櫛孔扇貝親環素A基因的克隆與表達[D]. 咸陽: 西北農林科技大學, 2009.
Song Xiaoyan. Molecular cloning, characterization and expression of Lectins fromArgopectenirradiansand cyclophilin a fromChlamysfarreri[D]. Xianyang: Northwest A & F University, 2009.
[19]胡鈺婷, 張殿昌, 江世貴, 等. 合浦珠母貝C型凝集素2的序列特征和表達分析[J]. 湖北農業科學, 2011, 50(19): 4026-4030.
Hu Yuting, Zhang Dianchang, Jiang Shigui, et al. Sequence and expression analysis of a novel C-type lectin gene from pearl oysterPinctadafucata[J]. Hubei Agricultural Sciences, 2011, 50(19): 4026-4030.
[20]Yamaura K, Takahashi K G, Suzuki T. Identification and tissue expression analysis of C-type lectin and galectin in the Pacific oyster,Crassostreagigas[J]. Comp Biochem Phys B, 2008, 149(1): 168-175.
[21]Li Hui, Zhang Huan, Jiang Shuai, et al. A single-CRD C-type lectin from oysterCrassostreagigasmediates immune recognition and pathogen elimination with a potential role in the activation of complement system[J]. Fish Shellfish Immunol, 2015, 44(2): 566-575.
[22]Jing Xing, Espinosa E P, Perrigault M, et al. Identification, molecular characterization and expression analysis of a mucosal C-type lectin in the eastern oyster,Crassostreavirginica[J]. Fish Shellfish Immunol, 2011, 30(3): 851-858.
[23]Wang Ning, Whang I, Lee J. A novel C-type lectin from abalone,Haliotisdiscusdiscus, agglutinatesVibrioalginolyticus[J]. Dev Comp Immunol, 2008, 32(9): 1034-1040.
[24]Zhang Jian, Qiu Reng, Hu Yonghua. HdhCTL1 is a novel C-type lectin of abaloneHaliotisdiscushannaithat agglutinates Gram-negative bacterial pathogens[J]. Fish Shellfish Immunol, 2014, 41(2): 466-472.
[25]高祥剛, 李云峰, 宋文濤, 等. 文蛤腸、外套膜和肝胰臟組織CDNA文庫構建及其ESTs序列分析[J]. 中國水產科學, 2010, 17(2): 344-350.
Gao Xianggang, Li Yunfeng, Song Wentao, et al. Construction of cDNA libraries from intestine, mantle and hepatopancreas of hard clam (Meretrixmeretrix) and ESTs analysis[J]. Journal of Fishery Sciences of China, 2010, 17(2): 344-350.
[26]Li Hongjun, Liu Weidong, Gao Xianggang, et al. Identification of host-defense genes and development of microsatellite markers from ESTs of hard clamMeretrixmeretrix[J]. Mol Biol Rep, 2011, 38(2): 769-775.
[27]Feinberg H, Taylor M E, Weis W I. Scavenger receptor C-type lectin binds to the leukocyte cell surface glycan Lewisx by a novel mechanism[J]. J Biol Chem, 2007, 282(23): 17250-17258.
[28]Sonnhammer E L L, Von Heijne G, Krogh A. A hidden Markov model for predicting transmembrane helices in protein sequences[C]//Ismb. 1998, 6: 175-182.
[29]Zhu Ling, Song Linsheng, Xu Wei, et al. Identification of a C-type lectin from the bay scallopArgopectenirradians[J]. Mol Biol Rep, 2009, 36(5): 1167-1173.
[30]Zhang Huan, Wang Hao, Wang Lingling et al. Cflec-4, a multidomain C-type lectin involved in immune defense of Zhikong scallopChlamysfarreri[J]. Deve Comp immunol, 2008, 149(1): 168-175.
[31]Zhang Huan, Wang Hao, Wang Lingling, et al. A novel C-type lectin (Cflec-3) fromChlamysfarreriwith three carbohydrate-recognition domains[J]. Fish Shellfish Immunol, 2009, 26(5): 707-715.
Cloning and expression analysis of a C-type lectin gene (Mm-Lec1) in hard clam Meretrix meretrix
Zhang Jingjing1,2, Li Hongjun2, Qin Yanjie1, Liu Min1,2, Ye Sheng1,2
(1.CollegeofFisheriesandLifeScience,DalianOceanUniversity,Dalian116023,China; 2.MarineEcologyDepartment,NationalMarineEnvironmentalMonitoringCenter,Dalian116023,China)
Abstract:The hard clam (Meretrix meretrix) is an economically important bivalve species in China. Aquaculture of M. meretrix is seriously affected by the epidemic diseases. The study of the immune system of M. meretrix is an important approach to solve disease problems. C-type lectins (CTLs) play important roles in the identification of pathogen associated molecular patterns and activation of humoral immunity. In the present study, a C-type lectin gene of M. meretrix(denoted as Mm-Lec1) was obtained through sequencing full-length cDNA library. The full-length cDNA of Mm-Lec1 was 586 bp with a 486 bp open reading frame, encoding 161 amino acids. The length of 5′ and 3′ untranslated region was 21 and 79 bp, respectively. The molecular weight of predicted protein was 18.65 kD, and the theoretical isoelectric point was 4.98. The predicted amino acid sequence had a signal peptide (Met1-Ser19), a sugar-binding site (QPN) and a carbohydrate recognition domain (CRD). The tertiary structure of Mm-Lec1 was predicted as a compact type and had a β sheet structure with three beta sheet layers. The similarity between Mm-Lec1 and the other species mentioned in this study were 20%-32%. Mm-Lec1, Mytilus galloprovincialis CTL 6 and Chlamys farreri CTL A were clustered in one branch in the neighbor-joining (NJ) tree. Mm-Lec1 mRNA was expressed in all tested tissues, including gill, haemocytes, hepatopancreas, mantle, adductor muscle and gonad, with the highest expression level in gill, the second in haemocytes, and the least in gonad. In the bacteria exposure test, the mRNA expression level was lowest at 6 h, and highest at 48 h, suggesting that Mm-Lec1 play a role in defensing against bacterial invasion in M. meretrix.
Key words:Meretrix meretrix; C-type lectin; EST; tertiary structure; real-time PCR
收稿日期:2015-10-15;
修訂日期:2016-01-05。
基金項目:國家自然科學基金(31101899,31572595);國家海洋局近岸海域生態環境重點實驗室開放基金(201511)。
作者簡介:張晶晶(1989—),女,河南省周口市人,研究方向為分子生態學。E-mail:liutian.3090@163.com *通信作者:李宏俊,博士,副研究員,研究方向為分子生態學。E-mail:hjli@nmemc.org.cn
中圖分類號:S917.4
文獻標志碼:A
文章編號:0253-4193(2016)06-0110-09
張晶晶, 李宏俊, 秦艷杰, 等. 文蛤C型凝集素基因(Mm-Lec1)的克隆與表達分析[J]. 海洋學報, 2016, 38(6): 110—118, doi:10.3969/j.issn.0253-4193.2016.06.012
Zhang Jingjing, Li Hongjun, Qin Yanjie, et al. Cloning and expression analysis of a C-type lectin gene (Mm-Lec1) in hard clamMeretrixmeretrix[J]. Haiyang Xuebao, 2016, 38(6): 110—118, doi:10.3969/j.issn.0253-4193.2016.06.012
