IL-21調節外周血中Treg細胞的表達在 Graves病發病機制中的研究①
2016-06-29 05:41:06黃小慶
中國免疫學雜志 2016年6期
黃小慶 劉 純
(重慶醫科大學附屬第一醫院內分泌科,重慶400016)
IL-21調節外周血中Treg細胞的表達在 Graves病發病機制中的研究①
黃小慶劉純
(重慶醫科大學附屬第一醫院內分泌科,重慶400016)
[摘要]目的:探究IL-21調節外周血Treg細胞的表達在Graves病發病機制中的研究。方法:收集28 例初發Graves病患者(GD)、27例經藥物治療后甲狀腺功能恢復正常的GD(eGD)患者和24例健康對照者(NC),電化學發光法檢測患者的血清 FT3、FT4、 uTSH、TgAb、TPOAb 和 TRAb 水平。分離研究對象外周血單個核細胞分為IL-21刺激組與未刺激組。實時熒光定量PCR法檢測Foxp3和IL-10 mRNA的表達水平;ELISA 法檢測培養上清液中IL-10蛋白的表達。結果:GD組甲功和自身抗體水平與eGD 組和NC組比較,差異均有統計學意義(P<0.05);但eGD組甲功和自身抗體水平與NC組比較差異無統計學意義(P>0.05)。IL-21刺激前,GD組的Foxp3、IL-10 mRNA和IL-10蛋白表達水平均較eGD組和NC組明顯升高,差異有統計學意義(P<0.05) ,但eGD組與NC組比較差異無統計學意義(P>0.05)。經 IL-21 刺激后,GD組的Foxp3、IL-10 mRNA和IL-10蛋白的表達水平均顯著低于刺激前,差異有統計學意義(P<0.05)。結論:IL-21可能通過抑制Treg細胞的分化及其效應分子IL-10的產生,使Treg細胞數量和功能下降,降低其對效應T細胞的抑制能力,從而參與GD的發病過程。
[關鍵詞]IL-21;Graves病;Treg細胞;Foxp3;IL-10
Graves病(Graves disease,GD)是一種以甲狀腺素合成增多的器官特異性自身免疫性疾病,是內分泌科常見的疾病之一。傳統理論認為,Th1細胞、Th2細胞是主要參與免疫反應的CD4+T細胞;Th1細胞通過分泌細胞因子IL-2、IFN-γ,Th2細胞通過分泌細胞因子IL-4、IL-13和IL-5參與免疫反應[1,2]。Th17細胞是CD4+T細胞的一個新分支,它參與許多自身免疫性疾病的發病過程[3]。免疫系統的主要功能是識別并清除外來抗原,產生免疫記憶,并產生免疫耐受;免疫平衡的維持依賴于許多因素,其中包括Th17細胞的激活和Treg細胞的抑制,當免疫平衡被打破時,免疫系統趨向于激活,此時機體更容易發生自身免疫性疾病[4]。Th17/Treg的失衡參與了許多自身免疫性疾病的發病過程,如1型糖尿病(DM1)、多發性硬化(MS)、系統性紅斑狼瘡(SLE)和銀屑病等[5-8]。Treg細胞是CD4+T細胞的一個新亞群,具有免疫應答低下和免疫抑制特性,在維持機體免疫耐受和免疫應答穩態方面具有非常重要的作用。Treg細胞主要通過產生細胞因子IL-10、TGF-β而發揮生物學效應。許多自身免疫性疾病都與外周血Treg細胞缺陷有關[9]。叉頭蛋白P3(Forkhead box protein P3,Foxp3)是近幾年來發現的控制Treg細胞發育和功能的關鍵轉錄因子,其主要表達在Treg細胞的細胞核上[10]。Foxp3作為一個轉錄調控因子,通過直接調控多種基因來調節Treg的活性,目前關于Foxp3通過何種方式調控這些基因表達的機制尚未闡明。
IL-21由一系列CD4+T細胞亞型及NKT細胞產生[11]。IL-21受體存在于CD4+T、CD8+T、NK細胞、巨噬細胞、DC細胞及角質細胞上;IL-21通過與IL-2、IL-4、IL-7、IL-9、IL-13同源受體及IL-21特異性受體結合而發揮作用[12]。Th17細胞和Treg細胞的分化均需要TGF-β的參與[13], 只有TGF-β存在的情況下,CD4+T細胞分化為Th17細胞,在TGF-β和IL-21共同存在的情況下,CD4+T細胞分化為Treg細胞[14]。我們前期研究發現,IL-21可能通過調節Th17細胞分化而上調IL-17的表達水平,參與了GD的發病過程[15]。有研究表明IL-21不但不能誘導Treg的增殖[16],同時還會抑制Treg的抑制功能。本文通過檢測IL-21刺激前后外周血單個核細胞(PBMC)中Treg細胞特異性轉錄因子Foxp3及其分泌的細胞因子IL-10的水平,深入了解Treg細胞在GD中的作用和意義,進一步探討Graves病的發病機理。
1材料與方法
1.1材料
1.1.1研究對象收集2014年10月~2015年6月在重慶醫科大學附屬第一醫院內分泌科門診就診的Graves病患者,根據診斷標準分為GD 組和甲狀腺功能恢復正常的GD (eGD) 組。GD組:根據中華醫學會內分泌病分會2008年頒布的《中國甲狀腺疾病診治指南》[17]首次確診為GD 且未服用任何抗甲狀腺藥物的患者,有甲狀腺功能亢進的癥狀和體征;血漿游離 T3(FT3)或 T4(FT4)升高及敏感 TSH(sTSH)降低;均有不同程度彌漫性甲狀腺腫大;血漿 TRAb 抗體不同程度陽性;共28例,其中女性21例,男性7例。eGD組:明確診斷為GD的患者,經抗甲狀腺藥物治療時間≥1年且甲狀腺功能恢復正常,現甲巰咪唑(賽治)治療劑量5~10 mg/d的GD患者27例,其中女性21例,男性6例。正常對照組(NC):無甲狀腺疾病史及家族史,甲狀腺功能正常,甲狀腺自身抗體陰性,排除妊娠、其他自身免疫性疾病、過敏性疾病、炎癥性疾病等,年齡及性別匹配的24名健康志愿者設為正常對照組,其中女性16例,男性8例。所有入選對象均需排除患有1型糖尿病、系統性紅斑狼瘡、類風濕性關節炎、自身免疫性肝炎等常見的自身免疫性疾病、妊娠、急性感染及慢性炎癥性疾病等。
1.1.2實驗材料IL-21、anti-CD3和anti-CD28抗體(德國 MiltenyiBiotec 公司);RPMI1640 、胎牛血清(美國 Gibco 公司)淋巴細胞分離液(天津TBD生物技術發展中心);RNAiso Plus、PrimeScriptTMRT reagent Kit(Perfect Real Time)和SYBR?Premix Ex TaqTMⅡ(Tli RNaseH Plus)( 寶生物工程有限公司,中國);人IL-10 ELISA試劑盒(美國R&D公司);游離三碘甲狀腺原氨酸(Free triiodothyronine,FT3)、游離甲狀腺素(Free thyroxine,FT4)、超敏促甲狀腺激素(Ultra-sensitive thyrotropin,uTSH )、甲狀腺球蛋白抗體(Thyroglobulin antibody,TgAb )、甲狀腺過氧化物酶抗體(Thyroidperoxidase antibody,TPOAb)、促甲狀腺素受體抗體(Thyrotropin receptor antibody,TRAb)試劑盒及檢測儀器(美國貝克曼庫爾特),采用電化學發光法測定。
1.2方法
1.2.1樣本的采集和外周血單個核細胞(Peripheral blood mononuclear cells,PBMCs) 的分離及培養采集研究對象空腹靜脈10 ml于EDTA抗凝真空采血管內,Ficoll密度梯度離心法分離PBMC,調整PBMC細胞密度為2×106個/ml,懸浮于含10%胎牛血清的RPMI1640培養液中,分為IL-21刺激組和IL-21非刺激組,接種于24孔平底培養板中,兩組均加入1 μg/ml anti-CD3和1 μg/ml anti-CD28抗體,IL-21刺激組加入10 ng/ml IL-21,放于37℃、5%CO2的孵箱內孵育72 h,收集未貼壁細胞用于抽提RNA,收集培養上清液于-80℃下保存。活細胞數≥95%。
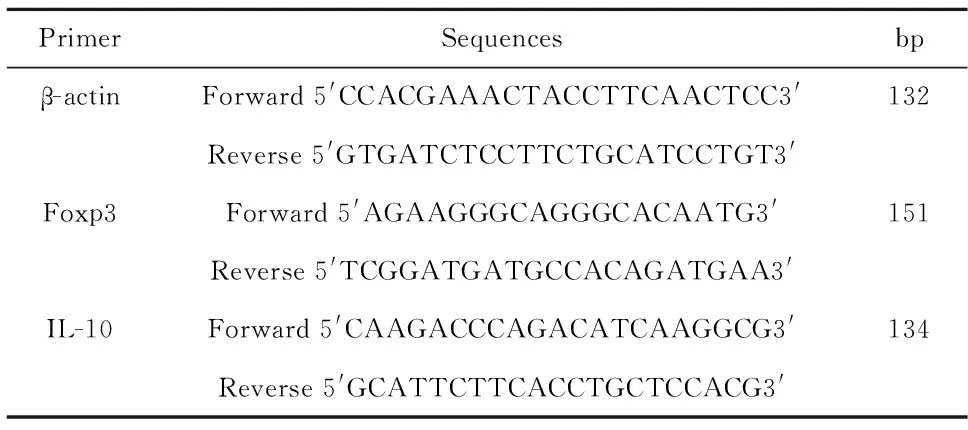
1.2.2熒光定量RT-PCR法測定Foxp3和IL-10 mRNA的表達細胞總RNA的提取按照RNAiso Plus的說明書進行。逆轉錄成cDNA按照PrimeScriptTMRT reagent Kit的說明書進行。聚合酶鏈反應,引物的設計采用 PrimerPremier 5.0。β-actin 、Foxp3和IL-10 mRNA的引物序列參見表1。使用熒光定量PCR 儀(CFX 96 Real-Time System,Bio-RAD,美國),反應條件為95℃,30 s;95℃,5 s;60℃,30 s;重復 39個循環。反應體系(10 μl):cDNA 1 μl,上游引物1 μl,下游引物1 μl,SYBE GREEN 5 μl,DEPC水2 μl。以未用IL-21干預的NC組為對照,計算各組2-△△Ct,獲得各組目的基因的相對表達量。
1.2.3ELISA法測定IL-10蛋白的表達取上述凍存細胞培養上清,ELISA法測定IL-10蛋白的表達,具體操作嚴格按試劑盒說明書進行。
1.2.4甲狀腺功能及抗體的檢測電化學發光法檢測FT3、FT4、uTSH、TgAb、TPOAb、TRAb的水平。

2結果
2.1甲功及相關抗體水平GD組甲功與eGD 組和NC組比較,差異均有統計學意義(P<0.05);但eGD組甲功與NC組比較差異無統計學意義(P>0.05)。GD組自身抗體水平較 eGD組和NC組增高,差異均有統計學意義(P<0.05);但eGD組自身抗體水平與NC組比較差異無統計學意義(P>0.05)。見表2。
表1β-actin、Foxp3和IL-10 mRNA 的引物序列
Tab.1Primer sequences for β-actin,Foxp3 and IL-10 mRNA
PrimerSequencesbpβ-actinForward5'CCACGAAACTACCTTCAACTCC3'132Reverse5'GTGATCTCCTTCTGCATCCTGT3'Foxp3Forward5'AGAAGGGCAGGGCACAATG3'151Reverse5'TCGGATGATGCCACAGATGAA3'IL-10Forward5'CAAGACCCAGACATCAAGGCG3'134Reverse5'GCATTCTTCACCTGCTCCACG3'
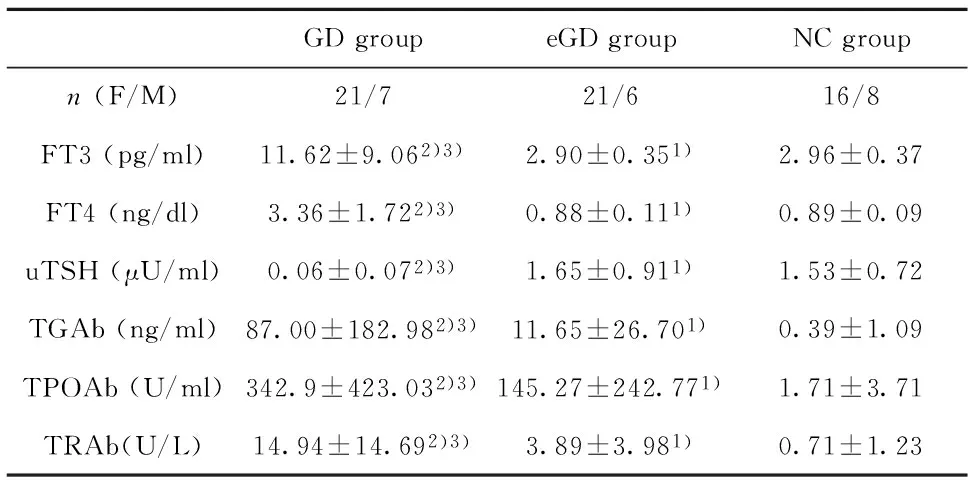
GDgroupeGDgroupNCgroupn(F/M)21/721/616/8FT3(pg/ml)11.62±9.062)3)2.90±0.351)2.96±0.37FT4(ng/dl)3.36±1.722)3)0.88±0.111)0.89±0.09uTSH(μU/ml)0.06±0.072)3)1.65±0.911)1.53±0.72TGAb(ng/ml)87.00±182.982)3)11.65±26.701)0.39±1.09TPOAb(U/ml)342.9±423.032)3)145.27±242.771)1.71±3.71TRAb(U/L)14.94±14.692)3)3.89±3.981)0.71±1.23
Note:1)P>0.05 vs.NC group;2)P<0.05 vs.NC group;3)P<0.05 vs.eGD group.
2.2Foxp3 mRNA表達水平未用IL-21處理時,GD組Foxp3 mRNA的表達量較eGD組和NC組增高,差異均有統計學意義(P<0.05);但eGD組與NC組比較差異無統計學意義(P>0.05)。經IL-21處理后,GD組、eGD組和NC組的Foxp3 mRNA的表達水平均有所降低,且較各組自身處理前有統計學意義(P<0.05)。見表3、圖1。
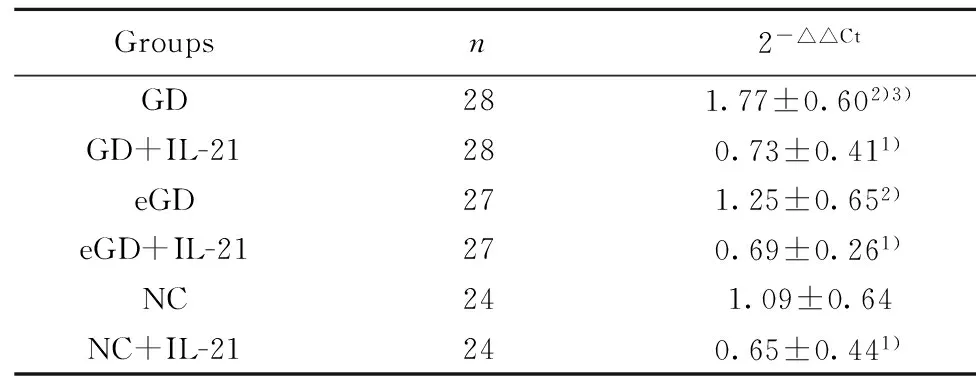
2.3IL-10 mRNA 表達水平未用IL-21處理時,GD組IL-10 mRNA的表達量較eGD組和NC組增高,差異均有統計學意義(P<0.05);但 eGD 組與NC組比較差異無統計學意義(P>0.05)。經IL-21處理后,GD組、eGD組和NC組的Foxp3 mRNA的表達水平均有所降低,且較各組自身處理前有統計學意義(P<0.05)。見表4、圖2。
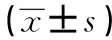
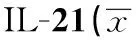

Groupsn2-△△CtGD281.62±0.702)3)GD+IL-21280.47±0.311)eGD271.11±0.592)eGD+IL-21270.73±0.321)NC240.94±0.44NC+IL-21240.51±0.181)
Note:Compared with the corresponding groups without IL-21,1)P<0.05;compared with NC group,2)P>0.05;compared with eGD group,3)P<0.05.
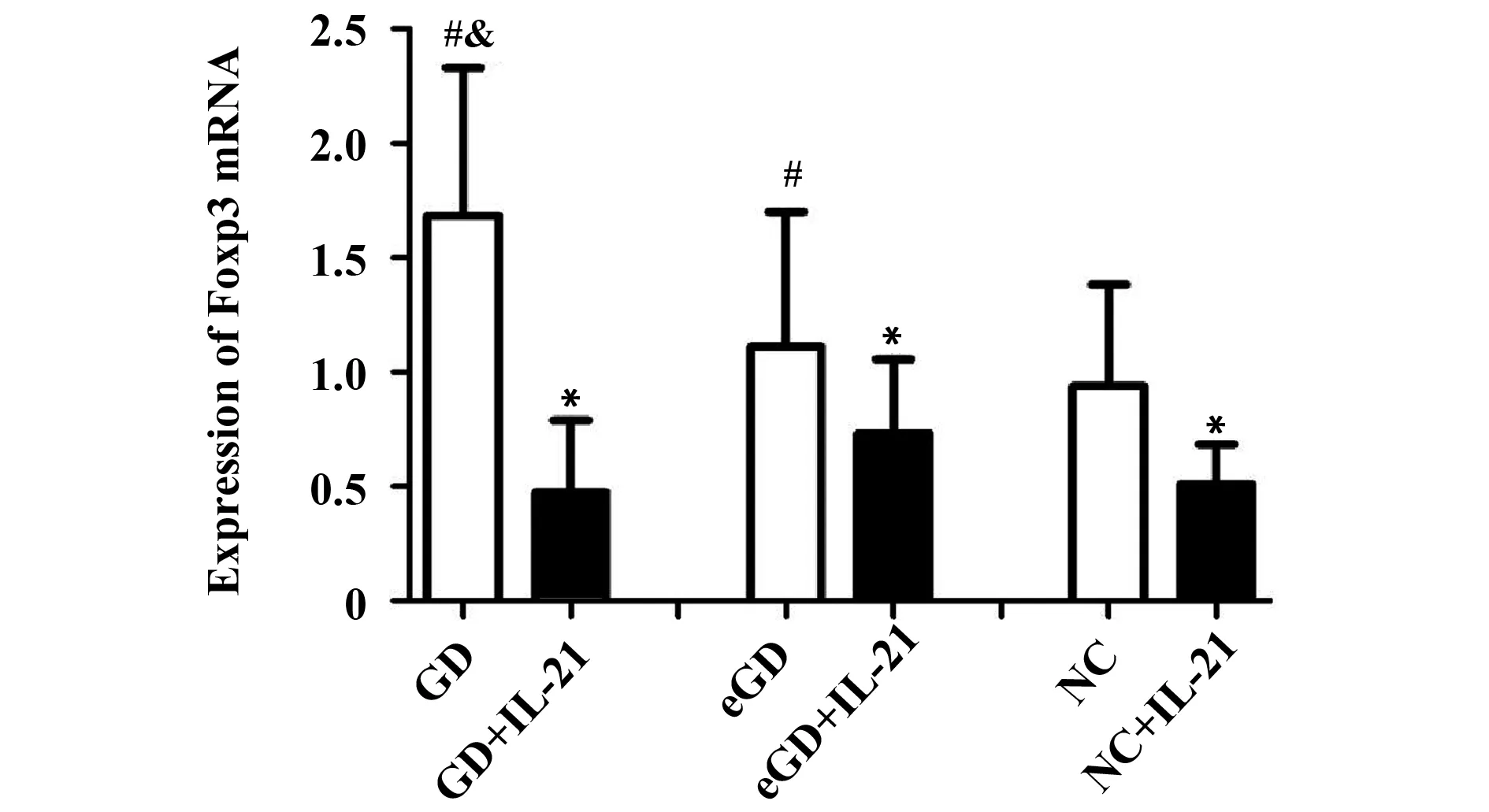
圖1 IL-21處理前后Foxp3 mRNA 表達水平比較Fig.1 Expression of Foxp3 mRNA before and after intervention of IL-21Note: Compared with the corresponding groups without IL-21,*.P<0.05;compared with NC group,#.P>0.05;compared with eGD group,&.P<0.05.
Groupsn2-△△CtGD281.77±0.602)3)GD+IL-21280.73±0.411)eGD271.25±0.652)eGD+IL-21270.69±0.261)NC241.09±0.64NC+IL-21240.65±0.441)
Note:Compared with the corresponding groups without IL-21,1)P<0.05;compared with NC group,2)P>0.05;compared with eGD group,3)P<0.05.
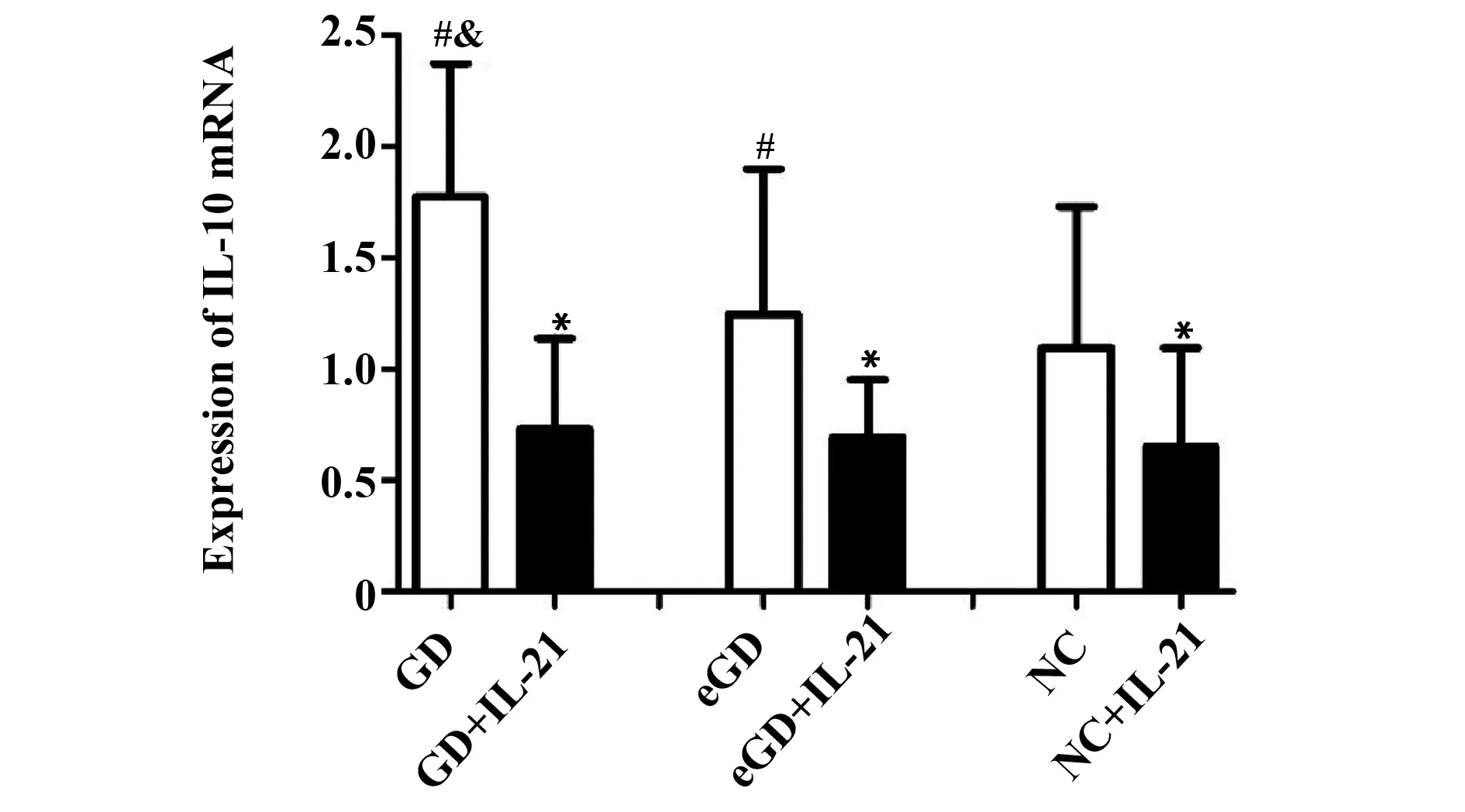
圖2 IL-21處理前后IL-10 mRNA表達水平比較Fig.2 Expression of IL-10 mRNA before and after intervention of IL-21Note: Compared with the corresponding groups without IL-21,*.P<0.05;compared with NC group,#.P>0.05;compared with eGD group,&.P<0.05.
2.4IL-10 蛋白表達水平未用IL-21處理時,GD組IL-10蛋白的表達量較eGD組和NC組增高,差異均有統計學意義(P<0.05);但eGD組與NC組比較差異無統計學意義(P>0.05)。經IL-21處理后,GD組、eGD組和NC組的IL-10蛋白的表達水平均有所降低,且較各組自身處理前有統計學意義(P<0.05)。見表5、圖3。
3討論
一類新的抑制性細胞Treg于1970年首次被Gershon和Kondo發現,這種細胞能夠在體外抑制免疫反應[18]。Treg細胞在維持免疫耐受方面發揮著至關重要的作用[19]。研究表明Treg細胞的功能缺陷或細胞數量減少參與了多種自身免疫性疾病的發病過程[20-27]。Foxp3是Treg細胞分化過程中的關鍵性轉錄因子,當Foxp3基因敲除,Treg細胞的數量減少,增加了自身免疫性疾病的發生率[28]。Sasaki等[29]的研究表明,在感染時IL-10 mRNA同IFN-γmRNA一樣都升高;這是因為機體免疫本身處于一個平衡狀態,促炎因子升高的同時相應的也會引起抗炎因子的升高,只是在疾病的不同階段促炎因子與抗炎因子所占的比重不同。
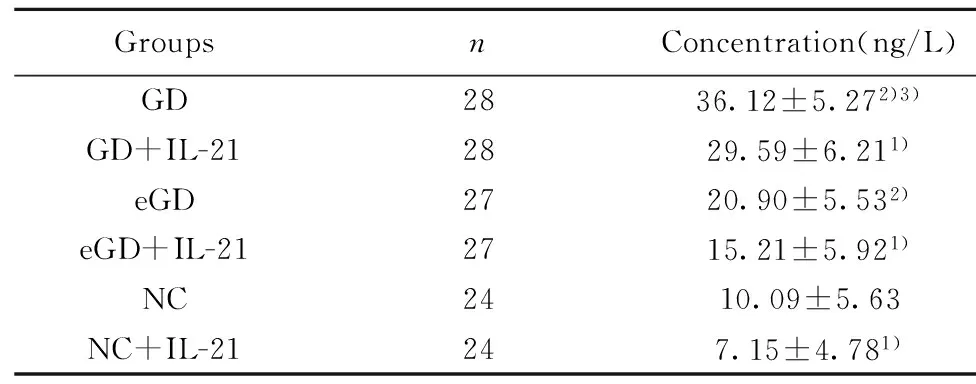
GroupsnConcentration(ng/L)GD2836.12±5.272)3)GD+IL-212829.59±6.211)eGD2720.90±5.532)eGD+IL-212715.21±5.921)NC2410.09±5.63NC+IL-21247.15±4.781)
Note:Compared with the corresponding groups without IL-21,1)P<0.05;compared with NC group,2)P>0.05;compared with eGD group,3)P<0.05.
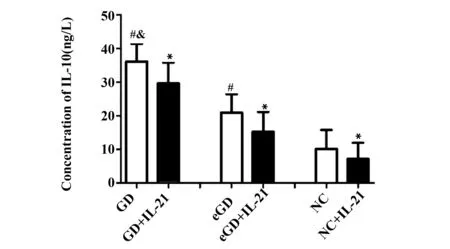
圖3 IL-21處理前后 IL-10蛋白表達水平比較Fig.3 Levels of IL-10 protein before and after intervention of IL-21Note: Compared with the corresponding groups without IL-21,*.P<0.05;compared with NC group,#.P>0.05;compared with eGD group,&.P<0.05.
本實驗結果中,GD患者外周血單個核細胞中的Foxp3 mRNA 和IL-10 mRNA表達水平顯著高于正常人群,而且其分泌的IL-10蛋白在GD患者中也明顯升高,這與以往研究結果相符[29]。這些結果說明在GD的發病過程中,Treg細胞的功能隨著免疫狀態的激活也受到一定程度的激活,且其通過增加分泌免疫抑制性細胞因子IL-10來使免疫達到平衡狀態。但Treg細胞具體通過何種途徑發揮作用仍不十分清楚。
IL-21是新近發現的IL-2細胞因子家族的新成員。IL-21主要由活化的各類CD4+T細胞亞型產生,其受體表達于多種免疫細胞,因此對多種免疫細胞有重要的生物調節作用。IL-21與其受體結合后通過激活Jak-STAT信號通路,從而引發一系列生物學效應[30]。研究表明,IL-21參與了多種自身免疫性疾病的發病過程,如系統性硬化、系統性紅斑狼瘡(SLE)、1型糖尿病、自身免疫性甲狀腺疾病[31-34]。在IL-21缺乏的情況下,IL-6誘導的CD4+T細胞中Foxp3+的表達量增加。Korn等[35]的研究發現,IL-6和TGF-β能夠誘導Th17細胞的分化;在IL-6缺失的情況下,IL-21協同TGF-β通過非IL-6依賴途徑使Th17細胞分化并抑制Treg細胞的表達。與細胞因子IL-2、IL-15和IL-7能誘導Treg細胞增殖的能力相反,IL-21非但不能誘導Treg細胞的增殖,反而還會抑制CD4+Treg細胞或CD8+Treg細胞的免疫抑制功能[16]。IL-21能通過活化CD4+T細胞亞型中的效應性T細胞來抑制調節性T細胞的活性[36]。以上研究表明IL-21可以作為一個獨立的因素在調節Treg細胞的分化過程中發揮著重要的作用。
本研究結果顯示,IL-21 刺激PBMCs后,GD組、eGD組和NC組的Foxp3 mRNA和IL-10 mRNA的表達水平顯著低于刺激前。這可能是因為IL-21可以通過下調轉錄因子Foxp3的表達,抑制了Treg細胞的活化[16],并抑制細胞因子IL-10的分泌[37]。IL-21 刺激PBMCs后,GD組較正常組Foxp3 mRNA和IL-10 mRNA降低趨勢更明顯,這提示在疾病狀態下,免疫系統被激活,機體更容易受炎癥因子的影響。因此,IL-21是促進GD發病的重要因素。
綜上所述,本研究表明IL-21可能通過抑制Treg細胞的分化及其效應分子IL-10的產生,使Treg細胞數量和功能下降,降低其對效應T細胞的抑制能力,從而參與GD的發病過程。由于本研究樣本量較小,所以IL-21在GD中的發病機制仍有待大樣本的研究資料進行驗證。
參考文獻:
[1]Noack M,Miossec P.Th17 and regulatory T cell balance in autoimmune and inflammatory diseases[J].Autoimmun Rev,2014,34(6):668-677.
[2]Song X,Qian Y.The activation and regulation of IL-17 receptor mediated signaling[J].Cytokine,2013,62(2):175-182.
[3]Duhen R,Glatigny S,Arbelaez CA,etal.Cutting edge: the pathogenicity of IFN-γ-producing Th17 cells is Independent of T-bet[J].J Immunol,2013,190(9):4478-4482.
[4]Peck A,Mellins ED.Breaking old paradigms: Th17 cells in autoimmune arthritis[J].Clin Immunol,2009,132(3):295-304.
[5]Esendagli G,Kurne AT,Sayat G,etal.Evaluation of Th17-related cytokines and receptors in multiple sclerosis patients under interferon β-1 therapy[J].J Neuroimmunol,2013,255(1/2):81-84.
[6]Ryba-Stanistawowska M,Skrzypkowska M,Myliwiec M,etal.Loss of the balance between CD4(+)Foxp3(+) regulatory T cells and CD4(+)IL17A(+) Th17 cells in patients with type 1 diabetes[J].Hum Immunol,2013,74(6):701-707.
[7]Vincze K,Kovats Z,Cseh A,etal.Peripheral CD4+cell prevalence and pleuropulmonary manifestations in systemic lupus erythematosus patients[J].Respir Med,2014,108(5):766-774.
[8]Zhang L,Yang XQ,Cheng J,etal.Increased Th17 cells are accompanied by FoxP3(+) Treg cell accumulation and correlated with psoriasis disease severity[J].Clin Immunol,2010,135(1):108-117.
[9]Sakaguchi S,Miyara M,Costantino CM.FOXP3(+) regulatory T cells in the human immune system[J].Nat Rev Immunol,2010,10(7):490-500.
[10]Maruyama T,Kono K,Mizukami Y,etal.Distribution of Th17 cells and FoxP3(+) regulatory T cells in tumor-infiltrating lymphocytes,tumor-draining lymph nodes and peripheral blood lymphocytes in patients with gastric cancer[J].Cancer Sci,2010,101(9):1947-1954.
[11]Leonard WJ,Zeng R,Spolski R.Interleukin 21: a cytokine/cytokine receptor system that has come of age[J].J Leukoc Biol,2008,84(2):348-356.
[12]Spolski R,Leonard WJ.The yin and yang of interleukin-21 in allergy,autoimmunity and cancer[J].Curr Opin Immunol,2008,20(3):295-301.
[13]Bettelli E,Carrier Y,Gao W,etal.Reciprocal developmental pathways for the Generation of pathogenic effector TH17 and regulatory T cells[J].Nature,2006,441(790):235-238.
[14]Leung S,Liu X,Fang L,etal.The cytokine milieu in the interplay of pathogenic Th1/Th17 cells and regulatory T cells in autoimmune disease[J].Cell Mol Immunol,2010,7(3):182-189.
[15]盧永霞,劉純.IL-21上調IL-17的表達在Graves病中的意義[J].上海交通大學學報:醫學版,2014,34(1):65-69.
[16]Peluso I,Fantini MC,Fina D,etal.IL-21 counteracts the regulatory T cell-mediated suppression of human CD4+T lymphocytes[J].J Immunol,2007,178(2):732-739.
[17]滕衛平,曾正陪,李光偉,等.中國甲狀腺疾病診治指南[M].北京:中華醫學會內分泌學分會,2007:47.
[18]Gershon RK,Kondo K.Cell interactions in the induction of tolerance: the role of thymic lymphocytes[J].Immunology,1970,18(5):723-737.
[19]Sakaguchi S,Yamaguchi T,Nomura T,etal.Regulatory T cells and immune tolerance[J].Cell,2008,133(5):775-787.
[20]Grant CR,Liberal R,Mieli-Vergani G,etal.Regulatory T-cells in autoimmune diseases: challenges,controversies and--yet--unanswered questions[J].Autoimmun Rev,2015,14(2):105-116.
[21]Kessel A,Haj T,Peri R,etal.Human CD19(+)CD25(high) B regulatory cells suppress proliferation of CD4(+) T cells and enhance Foxp3 and CTLA-4 expression in T-regulatory cells[J].Autoimmun Rev,2012,11(9):670-677.
[22]Liston A,Gray DH.Homeostatic control of regulatory T cell diversity[J].Nat Rev Immunol,2014,14(3):154-165.
[23]Dwivedi M,Kemp EH,Laddha NC,etal.Regulatory T cells in vitiligo: Implications for pathogenesis and therapeutics[J].Autoimmun Rev,2015,14(1):49-56.
[24]Osnes LT,Nakken B,Bodolay E,etal.Assessment of intracellular cytokines and regulatory cells in patients with autoimmune diseases and primary immunodeficiencies-novel tool for diagnostics and patient follow-up[J].Autoimmun Rev,2013,12(10):967-971.
[25]Gertel-Lapter S,Mizrachi K,Berrih-Aknin S,etal.Impairment of regulatory T cells in myasthenia gravis: studies in an experimental model[J].Autoimmun Rev,2013,12(9):894-903.
[26]Fenoglio D,Bernuzzi F,Battaglia F,etal.Th17 and regulatory T lymphocytes in primary biliary cirrhosis and systemic sclerosis as models of autoimmune fibrotic diseases[J].Autoimmun Rev,2012,12(2):300-304.
[27]Kornete M,Piccirillo CA.Critical co-stimulatory pathways in the stability of Foxp3+Treg cell homeostasis in Type I diabetes[J].Autoimmun Rev,2011,11(2):104-111.
[28]Zheng Y,Rudensky AY.Foxp3 in control of the regulatory T cell lineage[J].Nat Immunol,2007,8(5):457-462.
[29]Sasaki S,Nishikawa S,Miura T,etal.Interleukin-4 and interleukin-10 are involved in host resistance to Staphylococcus aureus infection through regulation of gamma interferon[J].Infect Immun,2000,68(5):2424-2430.
[30]Zeng R,Spolski R,Casas E,etal.The molecular basis of IL-21-mediated proliferation[J].Blood,2007,109(10):4135-4142.
[31]Lei L,He ZY,Zhao C,etal.Elevated frequencies of CD4(+) IL-21(+) T,CD4(+) IL-21R(+) T and IL-21(+) Th17 cells,and increased levels of IL-21 in bleomycin-induced mice May be associated with dermal and pulmonary inflammation and fibrosis[J].Int J Rheum Dis,2016,19(4):392-404..
[32]Wang HX,Chu S,Li J,etal.Increased IL-17 and IL-21 producing TCRαβ+CD4-CD8-T cells in Chinese systemic lupus erythematosus patients[J].Lupus,2014,23(7):643-654.
[33]Ferreira RC,Simons HZ,Thompson WS,etal.IL-21 production by CD4+effector T cells and frequency of circulating follicular helper T cells are increased in type 1 diabetes patients[J].Diabetologia,2015,58(4):781-790.
[34]Guan LJ,Wang X,Meng S,etal.Increased IL-21/IL-21R expression and its proinflammatory effects in autoimmune thyroid disease[J].Cytokine,2015,72(2):160-165.
[35]Korn T,Bettelli E,Gao W,etal.IL-21 initiates an alternative pathway to induce proinflammatory T(H)17 cells[J].Nature,2007,448(7152):484-487.
[36]Chevalier N,Thorburn AN,Macia L,etal.Inflammation and lymphopenia trigger autoimmunity by suppression of IL-2-controlled regulatory T cell and increase of IL-21-mediated effector T cell expansion[J].J Immunol,2014,193(10):4845-4858.
[37]Spolski R,Kim HP,Zhu W,etal.IL-21 mediates suppressive effects via its induction of IL-10[J].J Immunol,2009,182(5):2859-2867.
[收稿2015-10-11修回2015-10-27]
(編輯倪鵬)
Effect of IL-21 on Treg cells in peripheral blood mononuclear cells in pathogenesis of Graves disease
HUANG Xiao-Qing,LIU Chun.
Department of Endocrinology,The First Affiliated Hospital of Chongqing Medical University,Chongqing 400016,China
[Abstract]Objective:To explore the role of IL-21 in regulating the expression of peripheral blood Treg cells in the pathogenesis of Graves disease (GD).Methods: Electrochemical luminescence detection was used to determine levels of thyroid function indexes and autoantibodies.Peripheral blood mononuclear cells (PBMCs) were extracted,then,cultured in the presence or absence of IL-21 in vitro.The level of IL-10 protein was measured by enzyme-linked immunosorbent assay (ELISA),and expressions of Foxp3 and IL-10 mRNA were examined by Real-time PCR.Results: Compared to eGD and control group,there were significant differences in the levels of thyroid function indexes in GD group (P<0.05),whereas there was no difference between control group and eGD group (P>0.05).Before IL-21 stimulation,compared to eGD and control groups,expressions of Foxp3 and IL-10 mRNA and level of IL-10 protein were significantly higher in GD group(P<0.05);but there were no significant differences between eGD group and control group (P>0.05).After IL-21 stimulation,expressions of Foxp3,IL-10 mRNA and IL-10 protein levels were decreased markedly in all three groups,among which GD group showed the greatest change(P<0.05).Conclusion: IL-21 may inhibit the differentiation of Treg cells and the production of IL-10,therefore decreasing the number of Treg cells and the ability of suppressing effector T cells ,thus should be involved in the pathogenesis of GD.
[Key words]IL-21;Graves disease;Regulatory T cells;Foxp3;IL-10
doi:10.3969/j.issn.1000-484X.2016.06.018
作者簡介:黃小慶(1989年-),女,碩士,主要從事甲狀腺疾病免疫發病機制研究,E-mail:1548511929@qq.com。 通訊作者及指導教師:劉純(1963年-),女,主任醫師,碩士生導師,主要從事甲狀腺疾病免疫發病機研究,E-mail:liuchun200157@163.com。
中圖分類號R581.1
文獻標志碼A
文章編號1000-484X(2016)06-0853-06
①本文受重慶市渝中區科技計劃項目(20140127) 和 國家臨床重點專科建設項目資助(2011)。
猜你喜歡
新少年(2022年9期)2022-09-17 07:10:54
音樂探索(2022年2期)2022-05-30 21:01:37
美與時代·美術學刊(2022年3期)2022-04-27 01:18:15
小天使·一年級語數英綜合(2020年6期)2020-12-16 02:56:41
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
人大建設(2019年12期)2019-05-21 02:55:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
北極光(2014年8期)2015-03-30 02:50:51
鄭州大學學報(醫學版)(2015年2期)2015-02-27 14:50:46
中國火炬(2010年8期)2010-07-25 11:34:30