同步抑制棉葉螨TtAK與TtCHI基因表達的RNAi載體的構建
2016-06-06 03:33:34耿文超趙伊英汪小東李艷軍
西北農業學報 2016年4期
耿文超,趙伊英,劉 峰,汪小東,李艷軍,孫 杰
(石河子大學 農學院,新疆石河子 832000)
?
同步抑制棉葉螨TtAK與TtCHI基因表達的RNAi載體的構建
耿文超,趙伊英,劉峰,汪小東,李艷軍,孫杰
(石河子大學 農學院,新疆石河子832000)
摘要為在植物中表達針對棉葉螨精氨酸激酶基因TtAK與幾丁質酶基因TtCHI的雙鏈RNA,以改良植物對棉葉螨的抗性。采用PCR技術,分別擴增獲得靶標基因TtAK與TtCHI的功能區域片段453 bp與610 bp,通過PCR引物引入特定的酶切位點,采用DNA標準重組技術與Gateway技術相結合的方法,以RNA干擾載體pB7GWIWG2(Ⅱ)為基礎,成功構建出針對棉葉螨TtAK與TtCHI基因的RNAi載體。為遺傳轉化棉花,改良棉花對棉葉螨的抗性奠定基礎。
關鍵詞TtAK基因;TtCHI基因;Gateway技術;RNA干擾;載體構建
新疆棉花主要害螨為土耳其斯坦葉螨(Tetranychusturkestani),該螨屬蛛形綱蜱螨目葉螨科,廣泛存在于所有棉花種植區。其主要集中在棉花葉片背面,吸取汁液,自下而上造成棉株紅葉,抑制棉花的生長發育,并且引起蕾鈴大量的脫落。近年來,新疆棉區棉葉螨繁殖、擴散速度快,蟲口密度大,呈偏重發生態勢,危害嚴重時可造成棉田減產30%以上[1]。長期依賴化學藥劑防治,不但使棉葉螨的抗藥性增強,而且還殺死大量有害螨的天敵。因此,通過培育抗葉螨棉花品種來降低棉葉螨的危害程度,是新疆棉花生產亟待解決的一個重要問題。
近年來,通過轉基因的方法讓植物體內表達昆蟲基因的雙鏈RNA(dsRNA),當植食性昆蟲取食進入到昆蟲體內,從而抑制昆蟲相應靶標基因,表達的RNAi技術已應用于改良植物抗蟲性[2-7]。在昆蟲中,那些編碼重要蛋白功能的基因是RNAi很好的靶標;在植物中表達針對這些基因的dsRNA,通過昆蟲取食植物時,能夠特異地抑制相應昆蟲關鍵功能基因的表達,從而導致昆蟲的生長、發育以及繁殖的缺陷,甚至可以殺死昆蟲[2-7]。在無脊椎動物的生命活動過程中,精氨酸激酶(Arginine Kinase,AK)是參與能量代謝的關鍵酶[8];它僅存在于無脊椎動物的身體組織中,如腸上皮細胞、肌肉纖維等,是無脊椎動物體內唯一的磷酸原激酶;在其肌肉中,磷酸精氨酸是唯一有效形成ATP的磷酰基供體,能在一定時間內給劇烈運動的肌肉提供能量,同時又維持ATP的恒定水平,在細胞能量代謝中發揮著重要作用[9-10]。由于精氨酸激酶僅存在于無脊椎動物中,并且該酶及其磷酸原的能量代謝途徑與哺乳動物中的完全不同,因此針對該酶的抑制劑不會對人類造成危害。土耳其斯坦葉螨精氨酸激酶基因TtAK可作為植物介導的RNAi控制其危害的一個很好的敏感靶標。此外,幾丁質酶(Chitinase,CHI)是昆蟲發育過程中用來確保蛻皮的完成及外骨骼圍食膜和氣管等再生的重要酶[11-12],是昆蟲表皮、氣管和中腸圍食膜的重要組成成分,約占昆蟲蛻干物質的40%[13],對昆蟲的生長發育起著至關重要的作用[12]。因此,通過植物介導的RNAi技術抑制葉螨的幾丁質酶基因TtCHI表達,從而破壞其正常的生長發育過程,進而起到防治棉葉螨的作用,具有重要的意義。
本研究以植物介導的RNAi技術為前提,對棉葉螨2個靶標基因TtAK與TtCHI進行功能區域序列分析和比較,利用PCR技術分別擴增獲得的TtAK與TtCHI的靶標片段,通過引物引入的特定酶切位點,采用DNA重組技術和Gateway 技術,構建出雙基因的RNAi載體,為進一步遺傳轉化棉花等作物,進而為改良作物對棉葉螨的抗性奠定基礎。
1材料與方法
1.1材料、菌株及主要試劑
棉葉螨基因TtAK與TtCHI由石河子大學農學院昆蟲實驗室提供[14-15];大腸桿菌菌株EscherichiacoliTOP10、RNAi載體pB7GWIWG2 ((II)、根瘤農桿菌菌株AgrobacteriumtumefaciensLBA4404均為新疆兵團綠洲生態農業重點實驗室保存。
ExTaq酶和高保真DNA聚合酶PrimeSTARTM、克隆載體pMD18-T simple vector均購于大連寶生物有限公司;T4DNA連接酶、限制性內切酶、DNA凝膠回收試劑盒和DNA分子量標準Marker購自TaKara公司;試劑盒pENTRTMDirectional TOPO○RCloning Kits 和Gateway LR ClonaseTMⅡEnzyme Mix購于Invitrogen公司;抗生素等購自北京索來寶生物有限公司;其他試劑均為國產和進口分析純。
1.2靶標基因功能區域片段的克隆
以前期已克隆獲得的TtAK與TtCHI基因序列為基礎,選取相應的RNAi靶標區域,分別設計出特異性引物,正向引物AK-F:5′-GCTCAATACAAAGAAATGGAAG-3′;反向引物AK-R:5′-CTTCTTGTCAGCAGCAAGTTTAG GT-3′;正向引物CHI-F:5′-AAGCTTGCATGCCTGCAGGTC-3′(劃線部分為Hind Ⅲ酶切位點);反向引物CHI-R:5′-TCGATTGGATA- GAGATATTTGG-3′。 引物由華大生物工程(北京)股份有限公司合成。分別以獲得的2個棉葉螨基因cDNA為模板進行PCR反應。擴增條件為94 ℃ 5 min,30個循環(94 ℃ 1 min,56 ℃ 1 min,72 ℃ 1 min),72 ℃ 10 min。膠回收操作步驟參照百泰克生物公司膠回收試劑盒說明書。并且參照TaKara公司試劑盒說明,將回收的目的片段連接到pMD18 vector上,分別獲得pMD18-AK載體與pMD18-CHI載體。轉化大腸桿菌菌株E.coliTOP 10感受態細胞,篩選抗Amp菌落,同時進行菌液PCR鑒定和酶切鑒定。
1.3同步沉默雙基因的RNAi載體的構建
利用標準DNA重組技術與Gateway技術相結合的方法來構建質粒載體。用Hind Ⅲ酶切pMD18-CHI載體(pMD18 vector上帶有單一的Hind Ⅲ酶切位點,CHI擴增的正向引物設計1個Hind Ⅲ酶切位點),回收TtCHI靶標片段(610 bp)。同時,用Hind Ⅲ酶切pMD18-AK載體,回收線性大片段,為防止線性大片段產生自連現象,將收回的片段在磷酸酶作用下去磷酸化。然后將去磷酸化的大片段與回收TtCHI靶標片段(610 bp)在T4DNA連接酶進行連接,轉化E.coliTOP10感受態細胞后,挑取Kan抗性菌落,提取質粒并進行酶切鑒定,同時利用正向引物AK-F與反向引物CHI-R進行PCR擴增鑒定雙基因的連接方向,驗證后獲得pMD18-AK-CHI。
設計TtAK與TtCHI基因融合靶標片段的特異性引物,AK-CHI-F:5′-CACCGCTCAAT- ACAAAGAAATGG-3′;AK-CHI-R:5′-TCGA- TTGGATAGAGATATTTGG-3′。在正向引物前加上CACC接頭,利用PrimeSTARTMHS DNA聚合酶對重組質粒pMD18-AK-CHI進行PCR擴增,擴增反應體系為(總體系50 μL):5×PrimeSTAR 酶0.5 μL,5×PrimeSTAR buffer 10 μL,dNTP 4 μL,引物AK-CHI-F 1 μL,引物AK-CHI-R 1 μL,模版DNA(稀釋5倍)0.5 μL,加ddH2O補足至50 μL。擴增條件為94 ℃ 5 min,30個循環(94 ℃ 1 min,56 ℃ 1 min,72 ℃ 1 min),72 ℃ 10 min。回收擴增獲得的平末端PCR產物,與pENTR/D-TOPO載體進行TOPO定向克隆;轉化大腸桿菌菌株E.coliTOP10,篩選抗Kan菌落,同時進行菌落PCR鑒定和測序分析。利用Gateway技術,取測序結果正確的質粒進行LR反應,在0.5 mL離心管中加入以下反應體系(總體積4.5 μL):TOPO定向克隆獲得的入門載體2 μL,pB7GWIWG2(Ⅱ) 載體質粒2 μL,LR反應酶0.5 μL,瞬時離心將混合體系于25 ℃過夜后,加0.5 μL Proteinase K,37 ℃ 10 min 終止反應。轉化E.coliTOP10感受態細胞后,挑取Spec抗性的菌落進行PCR鑒定,獲得 pB7GWIWG2(Ⅱ)-AK-CHI載體(圖1)。通過電擊轉化法將pB7GWIWG2(Ⅱ)-AK-CHI載體導入到根瘤農桿菌菌株AgrobacteriumtumefaciensLBA4404中,挑取Spec抗性的菌落進行PCR鑒定。
2結果與分析
2.1靶標基因功能區域片段的克隆
以前期已克隆得到的TtAK與TtCHI基因序列為基礎,分別利用特異性引物PCR擴增,獲得453 bp 和610 bp的條帶(圖2)。將回收的目的片段分別連接到pMD18 vector上,轉化大腸桿菌菌株E.coliTOP10感受態細胞,篩選抗Amp菌落,提取質粒進行酶切鑒定。酶切目的片段與預期結果一致(圖3)。選取陽性克隆送往華大公司測序,結果表明,擴增的序列為TtAK與TtCHI基因靶標片段(圖4、圖5)。
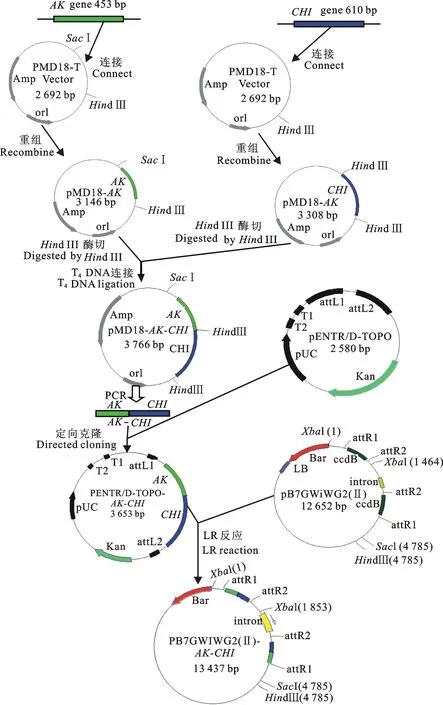
圖1 pB7GWIWG2(Ⅱ)-AK-CHI載體的構建示意圖
2.2同步沉默雙基因的RNAi載體的構建
用Hind Ⅲ酶切pMD18-CHI載體;由于pMD18 vector上帶有單一的Hind Ⅲ酶切位點,同時CHI擴增的正向引物設計了1個Hind Ⅲ酶切位點,所以酶切后回收pMD18-CHI的610 bp小片段;同時用Hind Ⅲ酶切pMD18-AK載體,回收線性大片段后去磷酸化。采用標準DNA重組技術,用T4DNA連接酶連接2個回收的片段。將其轉化大腸桿菌菌株感受態細胞,挑取抗性菌落,利用融合靶標片段的特異性引物進行PCR擴增。提取陽性質粒,用Hind Ⅲ等酶切進行分析。PCR與酶切驗證獲得pMD18-AK-CHI載體(圖6、圖7)。
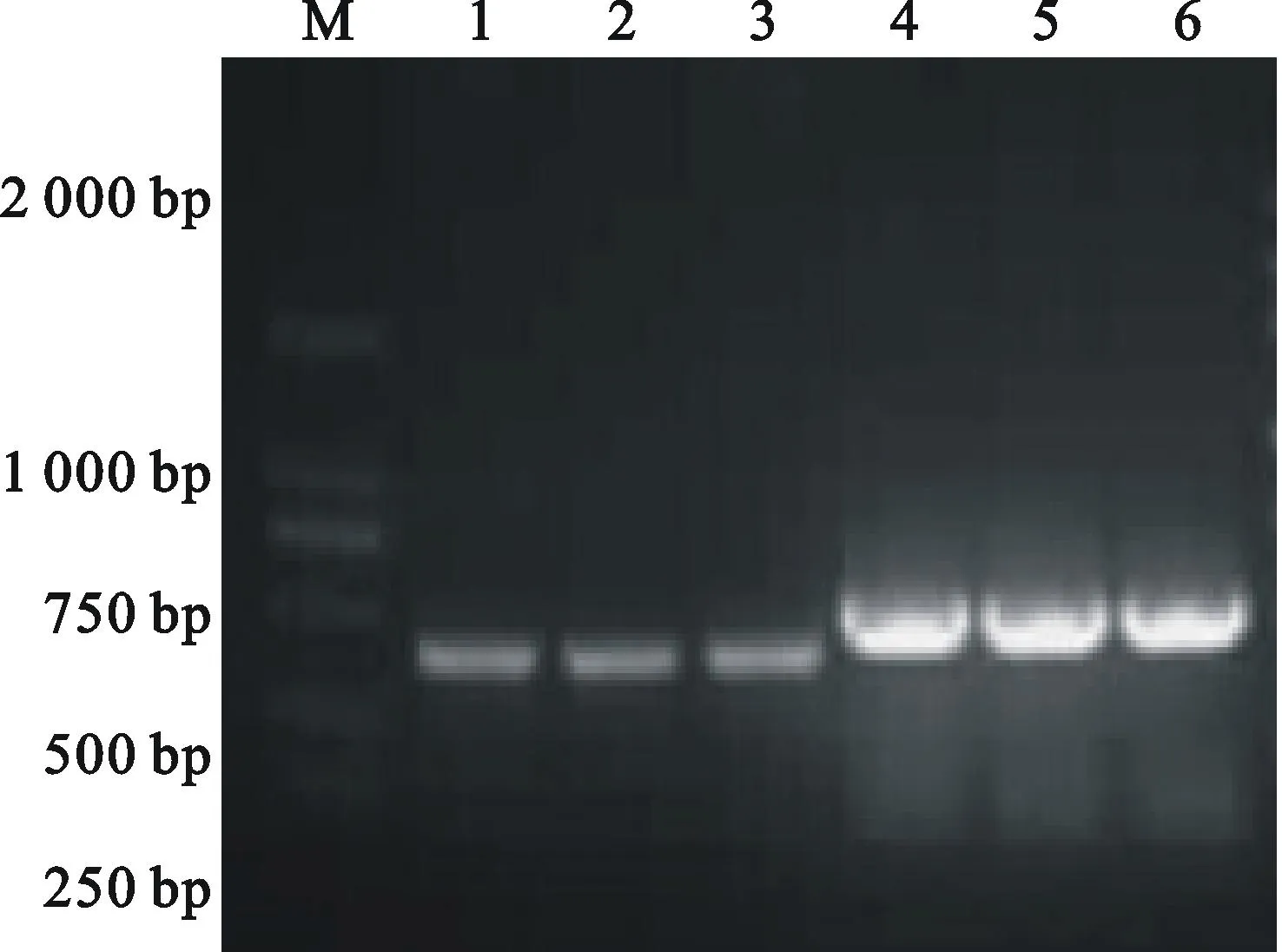
圖2PCR擴增靶標基因功能區域片段
Fig.2PCR amplification of target genes
利用Gateway重組技術,通過 PrimeSTARTMHS DNA聚合酶對重組質粒pMD18-AK-CHI的融合靶標片段進行PCR擴增,獲得的平末端PCR產物,與pENTR/D-TOPO載體進行TOPO定向克隆;轉化大腸桿菌菌株E.coliTOP10,篩選抗Kan菌落,同時進行菌落PCR鑒定和測序分析。將測序結果正確的入門克隆與RNAi載體pB7GWIWG2(Ⅱ)的重組位點在LR cloneⅡenzyme的催化下,進行LR反應。將AK-CHI的融合靶標片段以反向重復的形式插入到pB7GWIWG2(II)載體上,轉化大腸桿菌感受態細胞,挑Spec抗性單克隆進行菌落PCR 檢測(圖8)。提取陽性單克隆的質粒。通過XbaI 與SacIM.Marker DL 2 000; 1~3.TtAK基因的PCR產物PCR products ofTtAKgene; 4~6.TtCHI基因的PCR產物PCR products ofTtCHIgene
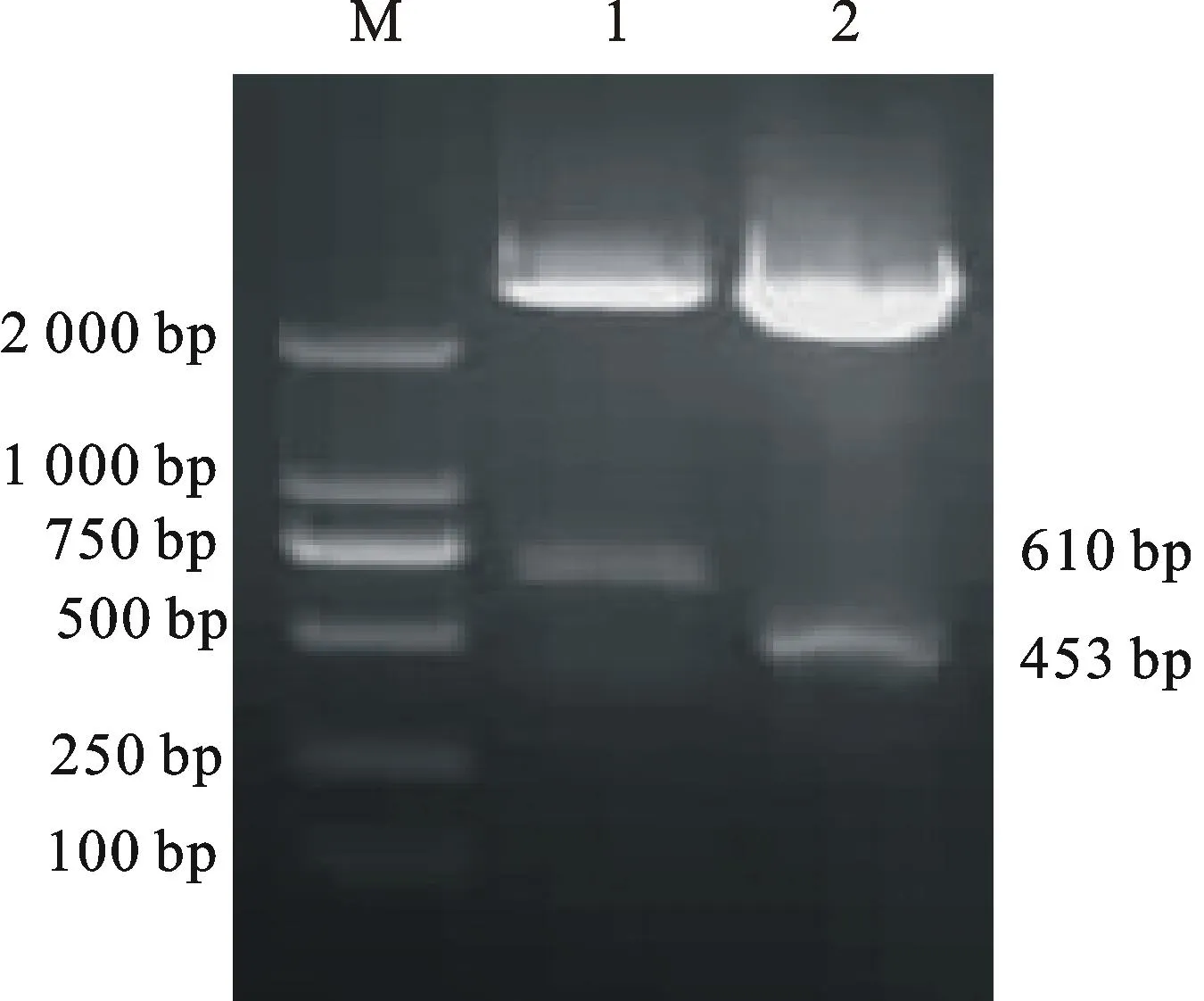
M.Marker DL 2 000; 1.HindⅢ與SacⅠ雙酶切pMD18-AK載體Fusion fragment PCR of pMD18-AKdigested byHindⅢ andSacⅠ; 2.HindⅢ酶切單pMD18-CHI載體Fusion fragment PCR of pMD18-CHIdigested byHindⅢ
圖3PCR擴增靶標基因功能區域片段
Fig.3PCR amplification of target gene

圖4 已克隆獲得 TtAK 靶標基因功能區域片段

圖5 已克隆獲得 TtCHI 靶標基因功能區域片段
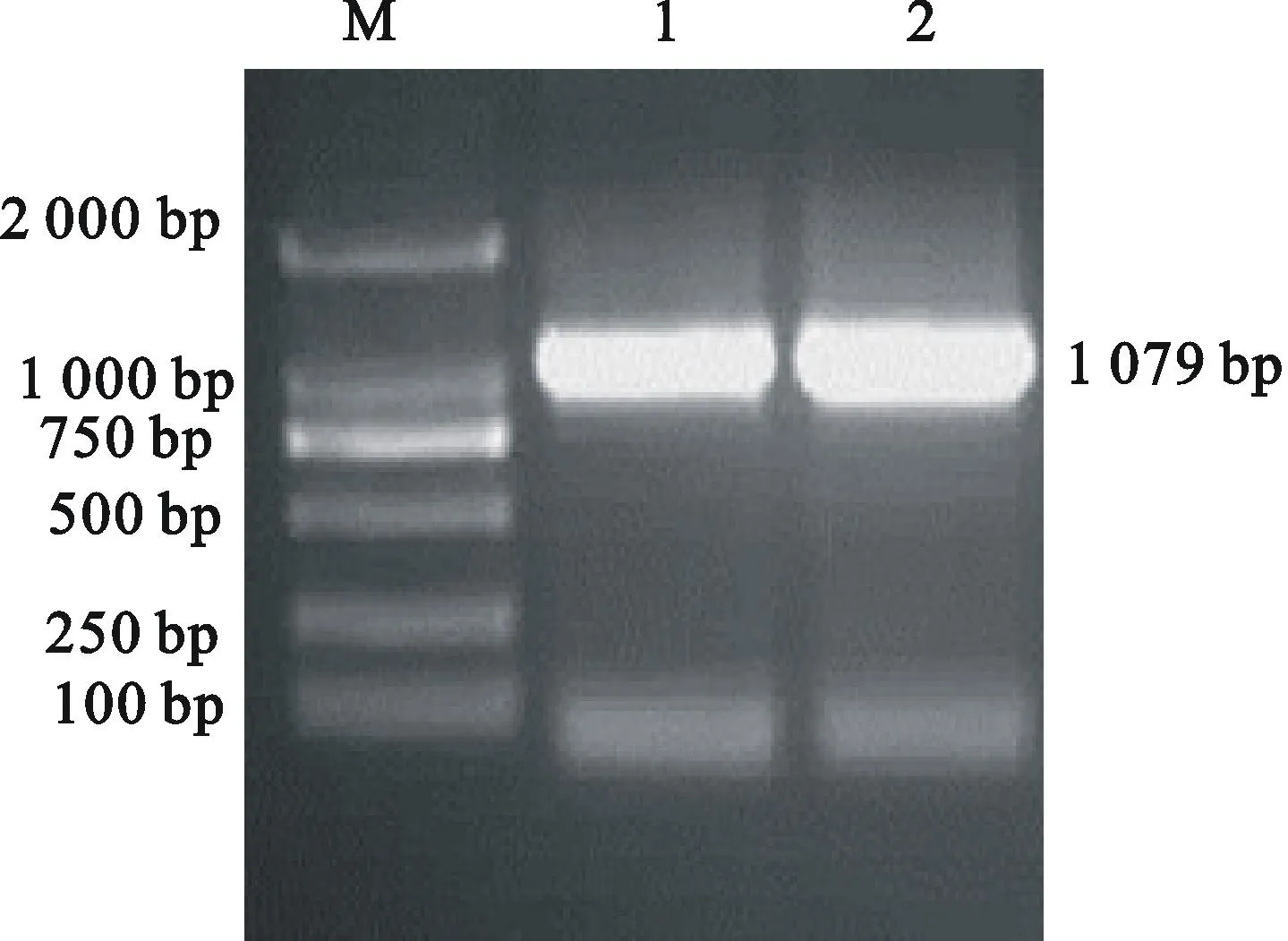
M.Marker DL 2 000; 1~2.pMD18-AK-CHIvector的PCR產物PCR products of pMD18-AK-CHIvector
圖6 pMD18-AK-CHI載體的PCR驗證
Fig.6 PCR identification of pMD18-AK-CHIvector
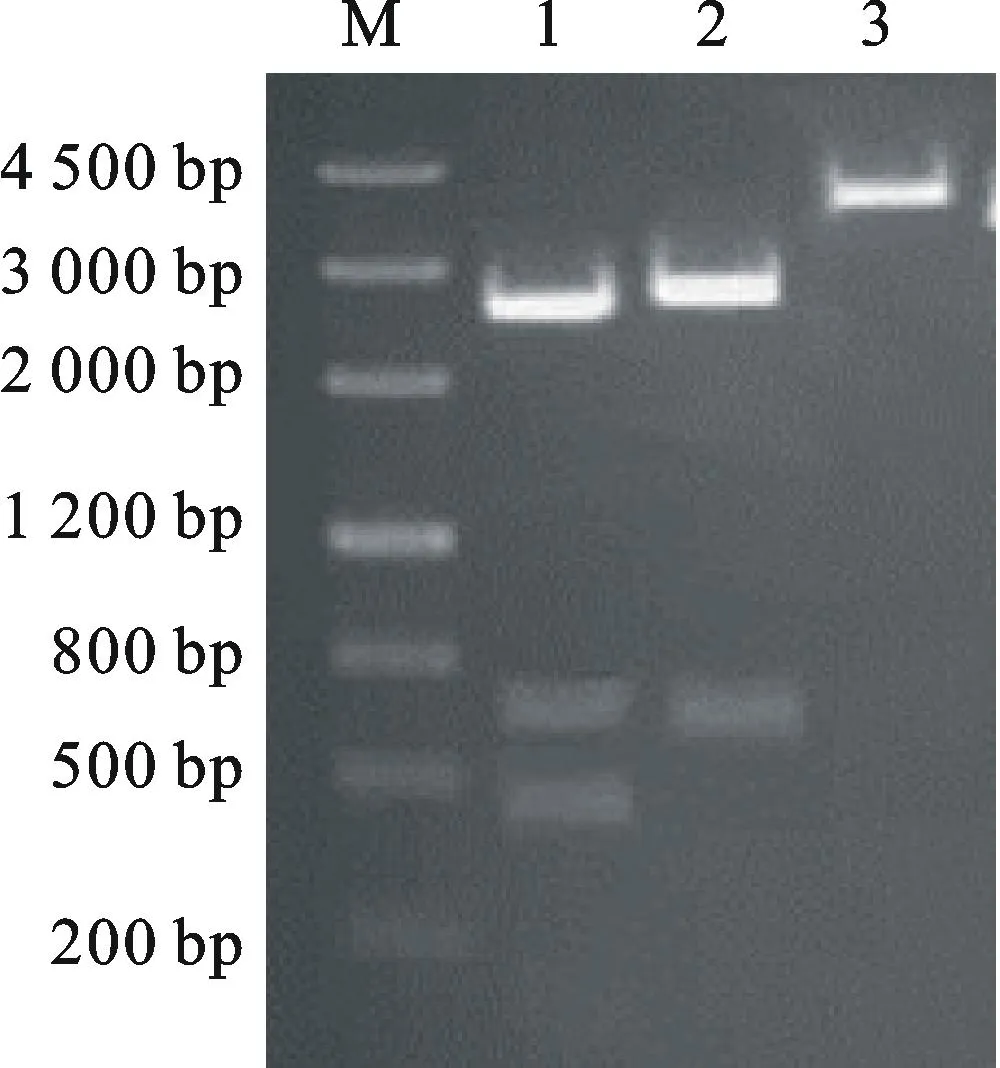
M.Marker Ⅲ; 1.Hind Ⅲ 與SacⅠ雙酶切 pMD18-AK-CHI載體Fusion fragment PCR of pMD18-AK-CHIdigested byHind Ⅲ andSacⅠ; 2.Hind Ⅲ 酶切 pMD18-AK-CHI載體Fusion fragment PCR of pMD18-AK-CHIdigested byHinⅢ; 3.SacⅠ酶切 pMD18-AK-CHI載體Fusion fragment PCR of pMD18-AK-CHIdigested bySacⅠ
圖7pMD18-AK-CHIvector載體的酶切驗證
Fig.7Restriction enzyme analysis of
pMD18-AK-CHIvector
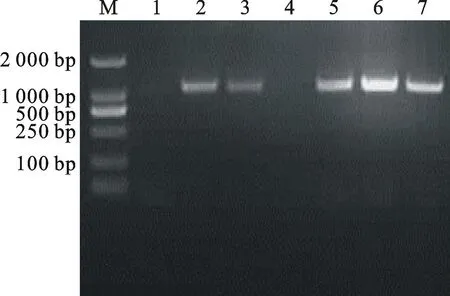
M.Marker DL 2 000; 1~5.pB7GWIWG2(Ⅱ)-AK-CHI的PCR產物PCR products of pB7GWIWG2(Ⅱ)-AK-CHIvector
圖8pB7GWIWG2(Ⅱ)-AK-CHI載體的PCR驗證
Fig.8PCR identification of pB7GWIWG2
(Ⅱ)-AK-CHIvector
雙酶切和XbaⅠ酶切驗證(圖9),獲得具有Spec抗性的RNAi載體pB7GWIWG2(Ⅱ)-AK-CHI載體。將構建好的RNAi載體通過電擊轉化法導入到根瘤農桿菌菌株AgrobacteriumtumefaciensLBA4404。經PCR鑒定,結果表明已成功將pB7GWIWG2(Ⅱ)-AK-CHI質粒轉化到LBA4404(圖10)。可用作于植物的遺傳轉化研究。
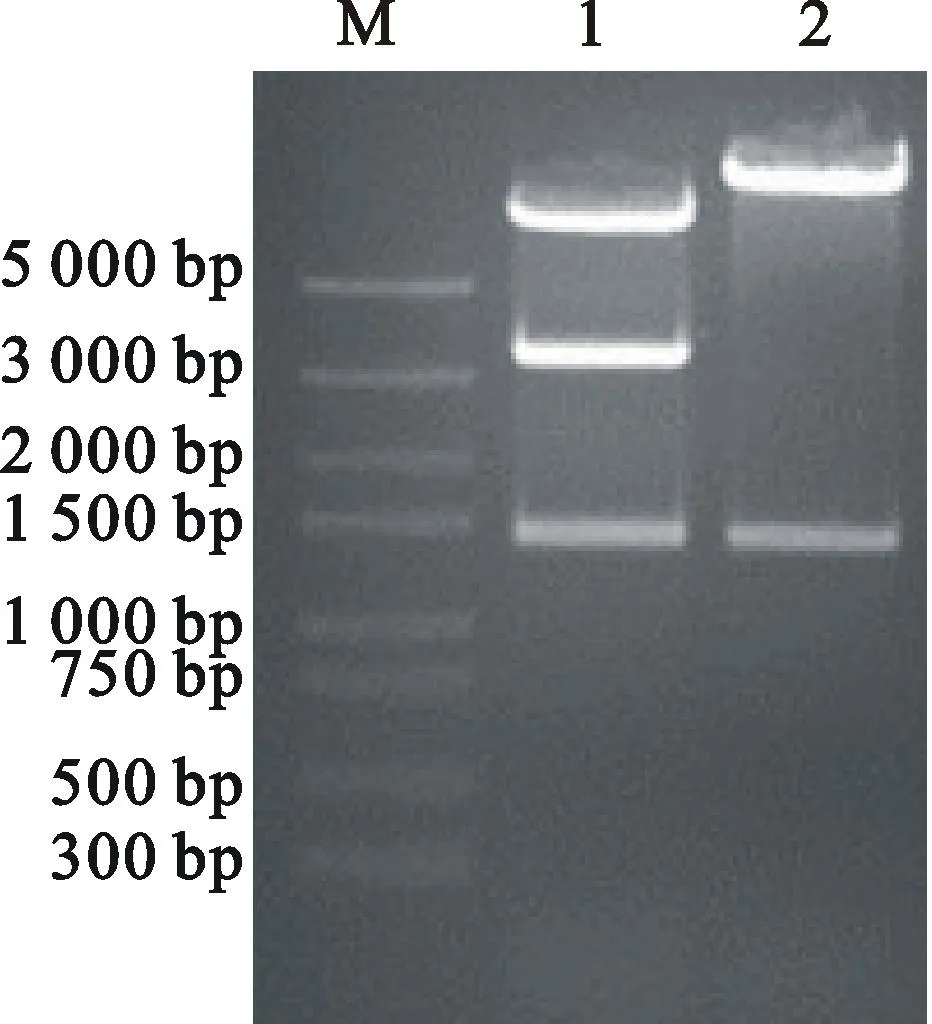
M.Trans5K DNA Marker; 1.XbalⅠ與SacⅠ雙酶切 pB7GWIWG2(Ⅱ)-AK-CHI載體Fusion fragment PCR of pB7GWIWG2(Ⅱ)-AK-CHIdigested byXbal Ⅰ andSacⅠ ;2.XbalⅠ 酶切pB7GWIWG2(Ⅱ)-AK-CHI載體Fusion fragment PCR of pB7GWIWG2(Ⅱ)-AK-CHIdigested byXbalⅠ
圖9pB7GWIWG2(Ⅱ)-AK-CHI載體的酶切驗證
Fig.9Restriction enzyme analysis
of pB7GWIWG2(Ⅱ)-AK-CHI
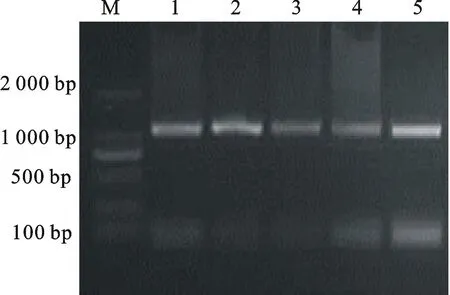
M.Marker DL 2 000; 1~5.pB7GWIWG2(Ⅱ)-AK-CHI載體的PCR擴增產物PCR products of pB7GWIWG2(Ⅱ)-AK-CHIvector
圖10 pB7GWIWG2(Ⅱ)-AK-CHI
載體導入農桿菌的PCR驗證
Fig.10PCR identification of pB7GWIWG2(Ⅱ)-AK-
CHIvector fromA.tumefaciens
3討論與結論
新疆由于特有的荒漠綠洲灌溉農業,已成為中國重要的棉花生產基地。近些年,隨著新疆棉花滴灌面積不斷擴大,棉田小生境和農田生態環境也在逐漸發生改變,致使近幾年螨害連年爆發,成為制約新疆棉花高產優質的重要因子[16]。有數據顯示,中國的二斑葉螨、朱砂葉螨已對多種的農藥產生抗性[17-18]。長期以來,中國農林作物害螨的防治一直以化學防治為主。化學農藥的廣泛使用,一方面殺傷大量天敵,使次要害蟲上升為主要害蟲和某些害蟲的再猖獗,另一方面導致害螨的抗藥性不斷增強,迫使農藥使用劑量增加或使用高毒農藥,嚴重污染自然環境,物種多樣性急劇減少,使生態系統變得十分脆弱,最終導致惡性循環。
自利用轉基因植物介導的RNAi抑制棉鈴蟲、玉米根蟲等危害取得成功以來,利用RNAi 控制害蟲迅速成為國際研究熱點[2-7]。RNAi 防治害蟲的效率依賴于高效的干擾片斷和簡便的導入方法。在昆蟲中,那些編碼重要蛋白的功能基因是RNAi 很好的靶標。本研究選擇棉葉螨的生長發育中2個關鍵酶基因精氨酸激酶基因TtAK與幾丁質酶基因TtCHI,通過標準的DNA重組技術構建這2個靶標基因的融合片段;進而利用Gateway技術,依據DNA重組與λ噬菌體位點的特異性重組原理,通過BP和LR兩步反應成功得獲得植物干擾載體pB7GWIWG2(II)-AK-CHI,為進一步遺傳轉化棉花,改良棉花對棉葉螨的抗性奠定基礎。
參考文獻Reference:
[1]尹奉諄.中國植物保護研究進展[M].北京:中國科學技術出版社,1996:120-123.
YIN F ZH.China Plant Protection Research[M].Beijing:China Science and Technology Press,1996:120-123(in Chinese).
[2]MAO Y B,CAI W J,WANG J W,etal.Silencing a cotton bollworm P450 monooxygenase gene by plant-mediated RNAi impairs larval tolerance of gossypol[J].NatureBiotechnology,2007,25(11):1307-1313.
[3]BAUM J A,BOGAERT T,CLINTON W,etal.Control of coleopteran insect pests through RNA interference[J].NatureBiotechnology,2007,25(11):1322-1326.
[4]GORDON K H,WATERHOUSE P M.RNAi for insect-proof plants[J].NatureBiotechnology,2007,25(11):1231-1232.
[5]KUMAR P,PANDIT S S,BALDWIN I T.Tobacco rattle virus vector:a rapid and transient means of silencing manduca sexta genes by plant mediated RNA interference[J].PLoSOne,2012,7(2):e31347.
[6]XIONG Y,ZENG H,ZHANG Y,etal.Silencing the HaHR3 gene by transgenic plant-mediated RNAi to disruptHelicoverpaarmigeradevelopment[J].InternationalJournalofBiologicalSciences,2013,9(4):370-381.
[7]WURIYANGHAN H,FALK B W.RNA interference towards the potato psyllid bactericera cockerelli,is induced in plants infected with recombinant tobacco mosaic virus(TMV)[J].PLoSOne,2013,8(5):e66050.
[8]GATTIS J L,RUBEN E,FENLEY M O,etal.The active site cysteine of arginine kinase structural and functional analysis of partially active mutants[J].Biochemistry,2004,43(27):8680-8689.
[9]PEREIRA C A,ALONSO G D,PAVETO M C,etal.Trypanosomacruziarginine kinase characterization and cloninga novel energetic pathway in protozoan parasites[J].JournalofBiologicalInorganicChemistry,2000,275(2):1495-1501.
[10]TANAKA K,ICHINARI S,IWANAMI K,etal.Arginine kinase from the beetleCissitescephalotes(Olivier).Molecular cloning,phylogenetic analysis and enzymatic properties[J].InsectBiochemistryandMolecularBiology,2007,37(4):338-345.
[11]BROWN A E,GROSSMAN S H.The mechanism and modes of inhibition of arginine kinase from the cockroach(Periplanetaamericana)[J].ArchivesofInsectBiochemistryandPhysiology,2004,57(4):166-177.
[12]KRAMER K J,HOPKINS T L,SCHAEFER J.Applications of solids NMR to the analysis of insect sclerotized structures[J].InsectBiochemistryandMolecularBiology,1995,25(10):1067-1080.
[13]ANDERSEN S O,HOJRUP P,ROEPSTORFF P.Insect cuticular proteins[J].InsectBiochemistryandMolecularBiology,1999,25(2):153-176.
[14]趙伊英,武萬峰,汪小東,等.精氨酸激酶基因的蛋白表達及實時熒光定量分析[J].石河子大學學報(自然科學版),2013,31(1):6-9.
ZHAO Y Y,WU W F,WANG X D,etal.Protein expression of arginine kinase and real-time PCR analysis[J].JournalofShiheziUniversity(NaturalScienceEdition),2013,31(1):6-9(in Chinese with English abstract).
[15]汪小東,劉峰,袁秀萍,等.二斑葉瞞幾丁質合成酶(CHS)基因的克隆及序列分析[J].西北農業學報,2014,23(5):39-44.
WANG X D,LIU F,YUAN X P,etal.Cloning and sequence analysis of chitin synthase(CHS) from spider mite[J].ActaAgriculturaeBoreali-occidentalisSinica,2014,23(5):39-44(in Chinese with English abstract).
[16]徐金虹.棗棉間作棉葉瞞猖獗發生的原因及綜防措施[J].新疆農業科技,2011(5):33.
XU J H.Causes and controlling measures of cotton apider mites’ rampant occurence in the conditions of jujube and cotton intercropping[J].XinjiangAgriculturalScienceandTechnology,2011(5):33(in Chinese).
[17]匡海源.我國重要農業害螨的發生與防治[J].農藥,1996,35(8):6-11.
KUANG H Y.The occurence and control of important agricultural mites in China[J].Pesticide,1996,35(8):6-11(in Chinese with English abstract).
[18]董順文,王朝生.棉花抗螨機理的研究進展[J].中國棉花,1999,26(12):2-5.
DONG SH W,WANG CH SH.Research progress of the mechanism in resistant cotton spider mites[J].ChinaCotton,1999,26(12):2-5(in Chinese).
Construction of RNAi for Synchronously Inhibiting Genes Vector ofTtAK
Received 2015-03-27Returned2015-04-27
Foundation itemNational Natural Science Foundation of China (No.31201521).
First authorGENG Wenchao,male,master student.Research area:seed biology and germplasm enhancement.E-mail:562517871@qq.com
(責任編輯:史亞歌Responsible editor:SHI Yage)
andTtCHIGenes Expression inTetranychusturkestani
GENG Wenchao,ZHAO Yiying,LIU Feng,WANG Xiaodong,LI Yanjun and SUN Jie
(College of Agriculture,Shihezi University,Shihezi Xinjiang832000,China)
AbstractIt’s necessary to produce transgenic plants which express the transcript of a double-stranded RNA (dsRNA) of Tetranychus turkestani AK and CHI gene for the purpose of improving plant resistance to cotton spider mites.The 453 bp and 610 bp fragments of dual-target gene TtAK and TtCHI were respectively amplified by PCR technology in this study firstly.Combining DNA recombinant technique with Gateway technology,the dual-target gene silencing RNAi vector pB7GWIWG2(Ⅱ)-AK-CHI based on RNAi vector pB7GWIWG2(Ⅱ) was successfully constructed.The study has laid the foundation for genetic transformation of cotton and improving plant resistance to cotton spider mites.
Key wordsTtAK gene; TtCHI gene; Gateway technology; RNA interference;Vector construction
收稿日期:2015-03-27修回日期:2015-04-27
基金項目:國家自然科學基金(31201521)。
通信作者:劉峰,男,副教授,研究方向為作物遺傳育種與分子生物學。E-mail:liufeng@shzu.edu.cn
中圖分類號Q966
文獻標志碼A
文章編號1004-1389(2016)04-0554-07
Corresponding authorLIU Feng,male,associate professor.Research area:crop genetics breeding and molecular biology.E-mail:liufeng@shzu.edu.cn
網絡出版日期:2016-04-02
網絡出版地址:http://www.cnki.net/kcms/detail/61.1220.S.20160402.1117.022.html
第一作者:耿文超,男,碩士研究生,研究方向為種子生物學與種質創新。E-mail:562517871@qq.com
