轉染人TBX18基因的大鼠心臟成纖維細胞中HCN4蛋白表達、If及對心肌細胞搏動頻率的影響觀察
2016-04-05 14:37:50權大君曹政楊媚李莎
山東醫藥 2016年47期
權大君,曹政,楊媚,李莎
(1武漢大學人民醫院,武漢430060;2武漢大學心血管研究所;3湖北省心血管重點實驗室;4十堰市太和醫院)
轉染人TBX18基因的大鼠心臟成纖維細胞中HCN4蛋白表達、If及對心肌細胞搏動頻率的影響觀察
權大君1,2,3,曹政4,楊媚1,2,3,李莎1,2,3
(1武漢大學人民醫院,武漢430060;2武漢大學心血管研究所;3湖北省心血管重點實驗室;4十堰市太和醫院)
目的 觀察轉染人TBX18基因的大鼠心臟成纖維細胞HCN4蛋白表達、超極化激活內向離子電流(If)及對心肌細胞搏動頻率的影響。方法 取新生SD大鼠,斷頭處死后取心臟,分離培養成纖維細胞及心肌細胞。構建人TBX18基因的腺病毒載體Ad-GFP-TBX18,包裝并純化病毒滴度至1×1010TU/mL。取傳代2~5代時對數生長期新生大鼠成纖維細胞分為A、B、C組,培養24 h后 A、B組分別轉染Ad-GFP-TBX18、腺病毒載體pHBAd-MCMV-GFP(陰性對照),C組不做任何處理。采用RT-PCR法檢測各組細胞TBX18 mRNA。采用Western blotting法檢測各組超極化激活環核苷酸門控陽離子通道4(HCN4)蛋白。采用電壓鉗記錄各組細胞超極化激活內向離子電流(If)。將各組細胞與新生大鼠心肌細胞共培養,觀察新生大鼠心肌細胞搏動頻率。結果 A、B、C組TBX18 mRNA相對表達量分別為98.32±6.21、1.03±0.05、1.02±0.02。A組TBX18 mRNA相對表達量高于B、C組(P均<0.05);B、C組TBX18 mRNA相對表達量差異無統計學意義。A、B、C組HCN4蛋白相對表達量分別為0.49±1.21、0.12±0.02 、0.09±0.01。A組HCN4蛋白相對表達量高于B、C組(P均<0.05);B、C組HCN4蛋白相對表達量差異無統計學意義。A組約有30%的細胞可以檢測到If,而B、C組中沒有細胞檢測到If。同一時點A組新生大鼠心肌細胞搏動頻率高于B、C組(P均<0.05);B、C組新生大鼠心肌細胞搏動頻率差異無統計學意義。結論 轉染人TBX18基因的大鼠心臟成纖維細胞高表達HCN4蛋白,存在If電流,并能促進共培養新生大鼠心肌細胞的搏動。
TBX18基因;基因轉染;成纖維細胞;心肌細胞;超級化激活環核苷酸門控陽離子通道4;超極化激活內向離子電流;心臟起博;動物實驗
近年來,緩慢性心律失常的發病率逐年升高。安裝心臟起搏器可緩解癥狀,但起搏器不能隨患者情緒變化、激烈運動情況做出自主調節,且有需定期更換電池、易受電磁波干擾及不適合兒童使用等缺點,因此尋找替代療法生物起搏器逐漸成為了研究熱點。最近,跨系重編程方法的出現使得一種細胞直接轉變成另一種細胞成為可能,如成纖維細胞經不同轉錄因子修飾可轉變成多功能干細胞[1]、神經細胞[2]、心肌細胞[3]、紅系或粒系細胞[4],為生物起搏器的研究提供了新的方向。有研究證實人類促生長轉錄因子TBX18可使心肌細胞直接重編程為類竇房結樣細胞,增加細胞膜上超極化激活環核苷酸門控陽離子通道(HCN4)的表達,從而提高其搏動頻率,且在豚鼠[5]與豬[6]心臟上得到了證實,但是尚未見大鼠上的相關報道。在竇房結細胞和浦肯野細胞4期自動去極化中超極化激活內向離子電流(If)發揮著重要的作用,而HCN通道是產生If的特異性離子通道。本研究將人TBX18基因轉染入新生大鼠成纖維細胞,觀察轉染后HCN4蛋白表達、If,并將其與大鼠心肌細胞共培養觀察心肌細胞搏動頻率。現將結果報告如下。
1 材料與方法
1.1 實驗動物、試劑及儀器 出生1~3 d的SD大鼠10只,雌雄不等,由武漢大學實驗動物中心提供。DMEM/F12(1∶1)培養液(Gibco),胎牛血清(GIBCO),胰蛋白酶(Sigma),Ⅱ型膠原酶(Sigma),5-溴脫氧尿核苷(Sigma);載體pHBAd-MCMV-GFP(Hanbio),DH5 (Tiangen),質粒DNA抽提試劑盒(康為世紀),BamHⅠ酶,NotⅠ酶;超極化激活環核苷酸門控陽離子通道 4蛋白(HCN4)羊抗鼠(一抗,Abcam);波形蛋白(Vimentin)羊抗兔(一抗,CST)。熒光顯微鏡(德國Leica公司,TCS SP2MP),PCR儀(杭州博日科技),電泳儀(北京市六一儀器廠),掃描儀(Canon)。
1.2 新生大鼠成纖維細胞及心肌細胞的分離培養 取10只出生1~3 d的SD大鼠,斷頭處死后取心臟組織,0.125%胰蛋白酶37 ℃恒溫消化10 min,棄上清液,加入組織體積4~5倍的0.08%膠原酶Ⅱ和0.125%胰酶的混合酶再次消化8~9次。取上清用含有10%FBS的低糖DMED終止消化,1 000 r/min離心10 min,用含有10%FBS的F12懸浮,用200目不銹鋼篩網過濾所得全部細胞,后接種于10 cm培養皿中, 37 ℃、5%CO2的培養箱中差速貼壁90 min。貼壁細胞即為成纖維細胞,未貼壁細胞即為心肌細胞。取培養第2天的心肌細胞與傳代第3代的成纖維細胞制作細胞爬片,免疫熒光實驗鑒定細胞,熒光顯微鏡下,成纖維細胞Vimentin 抗原表達陽性細胞率為97%,心肌細胞α-actin抗原陽性細胞率90%。
1.3 Ad-GFP-TBX18腺病毒載體構建、包裝及滴度測定 用BamHⅠ和NotⅠ酶對pHBAd-MCMV-GFP腺病毒載體(上海Hanbio公司)進行雙酶切,同時用PCR擴增TBX18的開放閱讀框,使酶切產物與TBX18片段連接,構建成Ad-GFP-TBX18腺病毒載體。重組載體進行PCR測序分析,結果顯示插入片段序列正確。將Ad-GFP-TBX18腺病毒載體、Lipofectamine2000試劑與Opti-MEM 混合均勻后轉染至293細胞,當細胞變大變圓呈葡萄狀,出現明顯噬斑時開始收毒,并于液氮及37 ℃水浴中反復凍融3次,離心、過濾收集病毒濃縮液,此時Ad-TBX18滴度為1×1010PFU/mL,并于-80 ℃保存,已備后續實驗使用。
1.4 新生大鼠成纖維細胞成纖維細胞分組及基因轉染 取傳代2~5代時對數生長期新生大鼠成纖維細胞接種于到6孔板,5×105/孔。將細胞分為A、B、C組,每組6個復孔。細胞培養24 h后 A、B組分別轉染Ad-GFP-TBX18、腺病毒載體pHBAd-MCMV-GFP(陰性對照),C組為空白對照組,不做任何處理。72 h后在熒光顯微鏡下觀察細胞,檢測綠色熒光蛋白,挑選熒光強度大(代表轉染效率高)的細胞擴大培養,用于后續實驗。
1.5 各組細胞TBX18 mRNA檢測 采用RT-PCR法檢測上述各組細胞TBX18 mRNA。取各組對數生長期細胞,提取細胞總RNA,逆轉錄合成cDNA并進行熒光定量PCR檢測。所有操作均嚴格按照使用說明書進行。反應條件:95 ℃、1 min,95 ℃、15 s,58 ℃、20 s,72 ℃、45 s,退火60 ℃,每20 s升溫1 ℃至95 ℃。TBX18上游引物5′-GGAGACTTGGATGAGACAAGTGAT-3′,下游引物5′-TTGGCAAATGGATTCCTGTCT-3′;以GAPDH作為內參,上游引物5′-CGCTAACATCAAATGGGGTG-3′,下游引物5′-TTGCTGACAATCTTGAGGGAG-3′)。采用2-ΔΔCt表示各組TBX18 mRNA相對表達量。
1.6 各組細胞HCN4蛋白檢測 采用Western blotting法。轉染72 h后取各組對數生長期細胞,TBS潤洗2~3次,細胞裂解液混勻,冰浴30 min,獲取上清液。BCA法提取總蛋白,取40 μg總蛋白,12%SDS-PAGE電泳,將蛋白質條帶轉移至PVDF膜上,5%脫脂奶粉封閉,TBST洗膜,分別加入HCN4一抗( 1∶500)/GAPDH(1∶10 000),4 ℃過夜;TBST洗膜3次,5 min/次。二抗采用山羊抗兔-HRP / 兔抗大鼠-HRP (1∶10 000)室溫避光孵育30 min,TBST搖床上洗4次,5 min/次。顯影、曝光。以GADPH為內參,以Band-Scan軟件進行灰度掃描并進行定量分析。各組HCN4蛋白相對表達量用HCN4蛋白條帶灰度值和GAPDH條帶灰度值之比表示。
1.7 各組細胞If檢測 采用電壓鉗記錄各組細胞If。采用EPC-9放大器和Pulse軟件進行數據記錄和分析。將對數生長期各組細胞置于灌流槽內,經細胞外液(含氯化鈉 135 mmol/L、 氯化鉀 5.4 mmol/L、氯化鈣 1.8 mmol/L、 氯化鎂 1 mmol/L、 氯化鋇 2 mmol/L、4-羥乙基哌嗪乙磺酸緩沖液10 mmol/L、葡萄糖 10 mmol/L,pH值為7.4)持續恒溫灌流(2 mL/min)。電極阻抗2.5~5 MΩ并充灌細胞內液 (含K+-ATP 5 mmol/L、氯化鉀120 mmol/L、氯化鎂5 mmol/L、氯化鈣1 mmol/L、乙二醇雙2-氨基乙基醚四乙酸10 mmol/L,4-羥乙基哌嗪乙磺酸緩沖液10 mmol/L,pH值為7.3),串聯電阻4~8 MΩ。分別于室溫(22~25 ℃)和37 ℃條件下記錄If,維持電位-30 mV,測試電位逐漸降至-110 mV,階躍10 mV,鉗制時間6 000 ms引出。利用100 μmol/L氯化鋇去除IK1電流的干擾。
1.8 與各組細胞共培養的新生大鼠心肌細胞搏動頻率觀察 A、B組轉染48 h后,用0.025%胰蛋白酶消化成細胞懸液,以1∶5的比例與對數生長期的乳鼠心肌細胞共培養,并于37 ℃、含有5%CO2的培養箱中培養,2 d換一次培養基。培養第2、4、6、8、10、12、14天在倒置顯微鏡下觀察并記錄各組共培養細胞的搏動頻率。

2 結果
A、B、C組TBX18 mRNA相對表達量分別為B、C組的98.32±0.85、1.03±0.05、1.02±0.02。A組TBX18 mRNA相對表達量高于B、C組(P均 <0.05);B、C組TBX18 mRNA相對表達量差異無統計學意義。A、B、C組HCN4蛋白相對表達量分別為0.49±1.21、0.12±0.02、0.09±0.01,A組HCN4蛋白相對表達量高于B、C組(P均<0.05);B、C組HCN4蛋白相對表達量差異無統計學意義。A組約有30%的細胞可以檢測到If,而B、C組中沒有細胞檢測到If。不同培養時間各組新生大鼠心肌細胞搏動頻率比較見表1。同一時點A組新生大鼠心肌細胞搏動頻率高于B、C組(P均<0.05);B、C組新生大鼠心肌細胞搏動頻率差異無統計學意義。
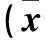
表1 不同培養時間各組新生大鼠心肌細胞搏動頻率比較±s)
注:與A組比較,#P<0.05。
3 討論
目前研究生物起搏的方法主要是通過干預最終使起搏細胞以外的其他細胞在保留自身表型的同時具有起搏細胞的特性和功能[5]。這些方法雖然可通過調節膜電位或使細胞去極化產生起搏電流,但存在電流不穩定、傳導阻滯或沒有自主調節等缺點。而最新研究發現不管在體外還是體內心肌細胞轉染TBX18基因后其表型與功能都向竇房結樣細胞轉變[6,7]。 TBX18是一種轉錄因子,主要負責胚胎向成熟心臟發育時竇房結頭部的形成,小鼠敲除TBX18基因后其竇房結發育較小但其功能變化不明顯[8]。乳鼠心肌轉染TBX18后可使其Cox43表達下降、因內向整流鉀通道表達減少從而提高其最大舒張電位、還可提高HCN4的表達且提高自發起搏頻率。還可提高cAMP表達水平調節“膜鐘”與“Ca2+鐘”,且觀察到自發LCR事件[5]。通過在大動物心室中直接注射TBX18基因發現心率可呈現晝夜變化且其心率受自主神經調節,搏位點起源于注射部位說明確實是TBX18在起作用。這種方法也不會引起的額外的心律失常,其他測量指標與正常動物相似[6]。說明TBX18可重編程心肌細胞從而發揮穩定的起搏功能。
成纖維細胞作為心臟中最主要的非心肌細胞,占到細胞總數的45%~75%[9],且心肌與成纖維細胞成交錯排列。生理狀態下,成纖維細可通過不斷合成和降解膠原蛋白與纖維連接蛋白起到支撐心臟結構的作用;還可通過與心肌形成縫隙連接蛋白直接影響心肌的電生理活動[10]。成纖維細胞是非興奮細胞,且其膜電阻較高、靜息電位成去極化相,且可產生機械敏感的電流。體外共培養時,心肌細胞與成纖維細間可形成Cox43和45供傳導沖動;成纖維細胞還能使比鄰的心肌細胞去極化從而降低其興奮性和傳導速率[11]。基于以上特點,成纖維細胞常被當做治療心律失常的靶點進行研究,如抑制成纖維細胞中Cox蛋白的表達提高傳導速率[12]或轉染MyoD可使成纖維細胞表達肌小管和心肌特異性蛋白從而促進傳導[13]。基于以上研究結果我們有理由提出如下假設:①在體動物心臟中注射TBX18基因,不僅心肌細胞可被轉染,成纖維細胞也可被轉染;②成纖維細胞經TBX18基因修飾后可改變其電生理特性,進而改變比鄰心肌的電生理特性。
HCN4是竇房結細胞在自動去極化時產生If的主要離子通道。胚胎發育早期心肌細胞與竇房結起搏細胞在結構與功能上相似,但在發育后期隨著HCN4的減少與If電流消失逐漸喪失其起搏功能。因此HCN4是維持竇房結細胞電生理功能的特異性蛋白。本研究結果顯示成纖維細胞轉染TBX18后HCN4蛋白表達增高,且通過膜片鉗記錄到電壓敏感的If,但并不能產生自發的動作電位,說明TBX18并不能像重編程心肌細胞一樣促使成纖維細胞轉變為起搏細胞。但用轉染TBX18的成纖維細胞共培養的心肌細胞搏動頻率比未轉染TBX18的成纖維細胞共培養的心肌細胞高20次/min,可能跟成纖維細胞膜上HCN4蛋白產生的去極化電流經縫隙連接流入比鄰的心肌細胞,促進心肌細胞動作電位4相自動去極化有關。
[1] Vierbuchen T, Ostermeier A, Pang ZP, et al. Direct conversion of fibroblasts to functional neurons by defined factors [J]. Nature, 2010,463(7284):1035-1041.
[2] Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors [J]. Cell, 2006,126(4):663-676.
[3] Ieda M, Fu JD, Delgado-Olguin P, et al. Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors [J]. Cell, 2010,142(3):375-386.
[4] Hu YF, Dawkins JF, Cho HC, et al. Biological pacemaker created by minimally invasive somatic reprogramming in pigs with complete heart block [J]. Sci Transl Med, 2014,6(245):245-294.
[5] Wiese C, Grieskamp T, Airik R, et al. Formation of the sinus node head and differentiation of sinus node myocardium are independently regulated by Tbx18 and Tbx3[J]. Circ Res, 2009,104(3):388-397.
[6] Szabo E, Rampalli S, Risueno RM, et al. Direct conversion of human fibroblasts to multilineage blood progenitors [J]. Nature, 2010,468(7323):521-526.
[7] Kapoor N, Liang W, Marban E, et al. Direct conversion of quiescent cardiomyocytes to pacemaker cells by expression of Tbx18[J]. Nat Biotechnol, 2013,31(1):54-62.
[8] Boink GJ, Robinson RB. Gene therapy for restoring heart rhythm [J]. J Cardiovasc Pharmacol Ther, 2014,19(5):426-438.
[9] Brown TR, Krogh-Madsen T, Christini DJ. Computational approaches to understanding the role of fibroblast-myocyte interactions in cardiac arrhythmogenesis[J]. Biomed Res Int, 2015,20(15):465-714.
[10] Goldsmith EC, Hoffman A, Morales MO, et al. Organization of fibroblasts in the heart[J]. Dev Dyn, 2004,230(4):787-794.
[11] Ongstad E, Kohl P. Fibroblast-myocyte coupling in the heart: Potential relevance for therapeutic interventions [J]. J Mol Cell Cardiol, 2016,91(4):238-246.
[12] Kizana E, Ginn SL, Allen DG, et al. Fibroblasts can be genetically modified to produce excitable cells capable of electrical coupling[J]. Circulation, 2005,111(4):394-398.
[13] Kizana E, Ginn SL, Smyth CM, et al. Fibroblasts modulate cardiomyocyte excitability: implications for cardiac gene therapy [J]. Gene Ther, 2006,13(22):1611-1615.
[14] Yang XJ, Zhou YF, Li HX, et al. Mesenchymal stem cells as a gene delivery system to create biological pacemaker cells in vitro [J]. J Int Med Res, 2008,36(5):1049-1055.
[15] Fahrenbach JP, Mejia-Alvarez R, Banach K. The relevance of non-excitable cells for cardiac pacemaker function [J]. J Physiol, 2007,585(2):565-578.
Expression of HCN4 protein and If in cardiac fibroblasts of rats transfected with human TBX18 gene and its effect on cardiomyocyte pulsation frequency
QUANDajun1,CAOZheng,YANGMei,LISha
(1RenminHospitalofWuhanUniversity,Wuhan430060,China)
Objective To observe the expression of HCN4 protein, If and its effect on the pulsation frequency of cardiomyocytes in cardiac fibroblasts transfected with human TBX18 gene.Methods The hearts were extracted after decapitation of the newborn SD rats, and the fibroblasts and cardiomyocytes were isolated and cultured. Adenovirus vector Ad-GFP-TBX18 was constructed and the virus titer was purified to 1×1010TU/mL. The fibroblasts were divided into groups A, B and C at the time of passage 2 to 5. After 24 hours of culture, groups A and B were transfected with Ad-GFP-TBX18 and pHBAd-MCMV- GFP, respectively, and group C without any treatment. The mRNA expression of TBX18 mRNA was detected by RT-PCR. The hyperpolarization-activated cyclic nucleotide-gated cation channel 4 (HCN4) protein was detected by Western blotting. The hyperpolarization activated inward current (If) was recorded by voltage clamp. The fibroblasts of each group were co-cultured with neonatal rat cardiomyocytes to observe the frequency of cardiomyocytes in neonatal rats.Results The relative expression levels of TBX18 mRNA in the groups A, B and C were 98.32±6.21, 1.03±0.05 and 1.02±0.02, respectively. The relative expression of TBX18 mRNA in the group A was higher than that in the groups B and C (allP<0.05). There was no significant difference in the relative expression of TBX18 mRNA between group B and group C. The relative expression of HCN4 protein in the groups A, B and C were 0.49 ± 1.21, 0.12 ± 0.02, and 0.09 ± 0.01, respectively. The relative expression of HCN4 protein in the group A was higher than that in the groups B and C (P<0.05). There was no significant difference in the relative expression of HCN4 protein between group B and group C. About 30% of the cells in group A were able to detect If, whereas in the groups B and C we did not detect If. At the same time, the frequency of cardiomyocyte pulsation in group A was higher than that in the groups B and C (allP<0.05). There was no significant difference in the frequency of cardiomyocytes between groups B and C.Conclusion HCN4 protein is highly expressed in rat cardiac fibroblasts transfected with human TBX18 gene and the If currents are present, which may promote the pulsation of neonatal rat cardiomyocytes.
TBX18 gene; gene transfection; fibroblast; cardiomyocyte; hyper-activated cyclic nucleotide-gated cation channel 4; hyperpolarization-activated inward current; heart pacing; animal experiment
國家自然科學基金面上項目(81270249);國家自然科學基金面上項目(81570306);湖北省科技支撐計劃項目(2013BCB013)。
權大君(1991-),男,碩士研究生在讀,主要研究方向為心電生理。E-mail: 411108619@qq.com
簡介:曹政(1977-),男,醫學博士,教授,副主任醫師,主要研究方向為冠心病與電生理。E-mail: caozheng908@163.com
10.3969/j.issn.1002-266X.2016.47.006
R54
A
1002-266X(2016)47-0021-04
2016-05-05)
