青蝦幼蝦發育時期性腺發育組織學研究
2016-02-07 12:40:22金舒博張穎關海紅傅洪拓蔣速飛熊貽偉喬慧張文宜龔永生吳滟
水產學雜志 2016年4期
金舒博,張穎,關海紅,傅洪拓,蔣速飛,熊貽偉,喬慧,張文宜,龔永生,吳滟
(1.南京農業大學無錫漁業學院,江蘇 無錫 214081;2.中國水產科學研究院淡水漁業研究中心,農業部淡水漁業與種質資源利用重點實驗室,江蘇 無錫 214081;3.中國水產科學研究院黑龍江水產研究所,黑龍江 哈爾濱 150070)
青蝦幼蝦發育時期性腺發育組織學研究
金舒博1,2,張穎3,關海紅3,傅洪拓1,2,蔣速飛2,熊貽偉2,喬慧2,張文宜2,龔永生2,吳滟2
(1.南京農業大學無錫漁業學院,江蘇 無錫 214081;2.中國水產科學研究院淡水漁業研究中心,農業部淡水漁業與種質資源利用重點實驗室,江蘇 無錫 214081;3.中國水產科學研究院黑龍江水產研究所,黑龍江 哈爾濱 150070)
采用組織切片及類固醇激素含量測定等方法,研究青蝦Macrobrachium nipponense幼蝦發育第1~31d的精巢、卵巢及促雄腺發育的起始時間、發育過程及成熟時間。結果表明:青蝦幼蝦發育到第10d(PL10)時促雄腺呈索狀結構開始發育,隨后經歷增殖期(PL10)、合成期(PL13)和分泌期(PL19)3個發育階段發育成熟,形成完整的促雄腺。青蝦精巢和卵巢均在幼蝦發育第13d(PL13)開始發育,精巢形成不規則排列的精原細胞,而卵巢生殖上皮開始分化為橢圓或多邊形的卵原細胞。精巢經精原細胞期(PL13)、精母細胞期(PL16)、精細胞期(PL19)和精子期(PL22)4個發育階段成熟,此時成熟的精巢生精小管內充滿成熟的精子。卵巢經卵原細胞期(PL13)、初級卵母細胞期(PL16)、次級卵母細胞期(PL16)和卵子期(PL19)4個時期發育成熟,此時卵巢充滿成熟卵子。從PL1到PL22,睪酮分泌量逐漸增加,維持在穩定水平至PL25后逐漸下降;從PL1到PL19,17β-雌二醇的分泌量逐漸增加,維持在穩定水平至PL25后逐漸下降。本研究顯示:促雄腺與精巢發育過程均持續10d,但促雄腺早于精巢3d開始發育和成熟,這可為性別和生殖相關基因的篩選及性別調控機制的研究提供參考。
青蝦;組織學觀察;類固醇激素測定;性腺發育;性早熟
青蝦Macrobrachium nipponense(日本沼蝦)是我國重要的淡水經濟蝦類,養殖范圍廣泛。青蝦的雌雄個體生長差異顯著,雄性個體的生長速度明顯快于雌性,同齡雄蝦的大小約為雌蝦的2~2.5倍[1]。青蝦全雄群體養殖具有巨大的經濟價值。
促雄腺(androgenic gland)又稱為促雄性腺,與性別分化、雄性發育及維持有關,在甲殼類性別調控中發揮關鍵作用[2],可促使甲殼動物向雄性方向發育[3]。在性別分化的早期,植入促雄腺或注射促雄腺提取物可導致雌性甲殼動物出現雄性第二性征而雄性化[4];性別已經分化的個體,人為摘除或植入促雄腺可使性逆轉,改變生理性別。圍繞促雄腺開展的研究已成為近些年來甲殼動物性別決定相關研究的重點。目前已研究了青蝦[5]、凡納濱對蝦Litopenaeus cannamei[6]、鋸緣青蟹Scylla serrata[7]、三疣梭子蟹Portunus trituberculatus[8]、中華絨螯蟹Eriocheir sinensis[9]等甲殼動物成體促雄腺的組織學。但尚未開展有關幼蝦發育過程中促雄腺的組織學研究,如青蝦促雄腺分化的起始時間、發育過程及成熟時間等。
性早熟是青蝦養殖過程中的嚴重問題。在繁殖季節,雌蝦卵巢發育周期非常短,新繁育的雌蝦性腺成熟早,可以在發育45d后與雄蝦交配產卵,導致多代同塘、種質退化、養殖密度增大等問題,嚴重制約青蝦養殖業的可持續發展[10]。因此,開展青蝦性腺發育的研究對性腺發育調控至關重要。
本實驗采用組織學切片方法研究青蝦幼蝦發育過程中促雄腺及性腺發育,以確定促雄腺及性腺分化的起始時間、發育過程及成熟時間;測定睪酮和17β-雌二醇的分泌量,探討類固醇激素分泌對青蝦精巢及卵巢發育的影響。
1 材料與方法
1.1 材料
野生健康的抱卵雌蝦購于無錫太湖邊,體質量為3.78~5.26g,養殖于實驗室中,24h充氣,水溫維持在30℃。日換水量約為總水量的1/2,每天投喂2次,直到受精卵進入幼蝦發育期。
1.2 方法
組織學觀察:從幼蝦發育第1~31d,每隔3d取樣一次,每次取8只幼蝦固定于Bouin氏固定液中,用于組織學觀察。實驗蝦的體質量見表1。常規石蠟包埋,連續切片,切片厚6μm。Ehrlich氏蘇木精與伊紅對比染色,Olympus SZX16顯微鏡下觀察。
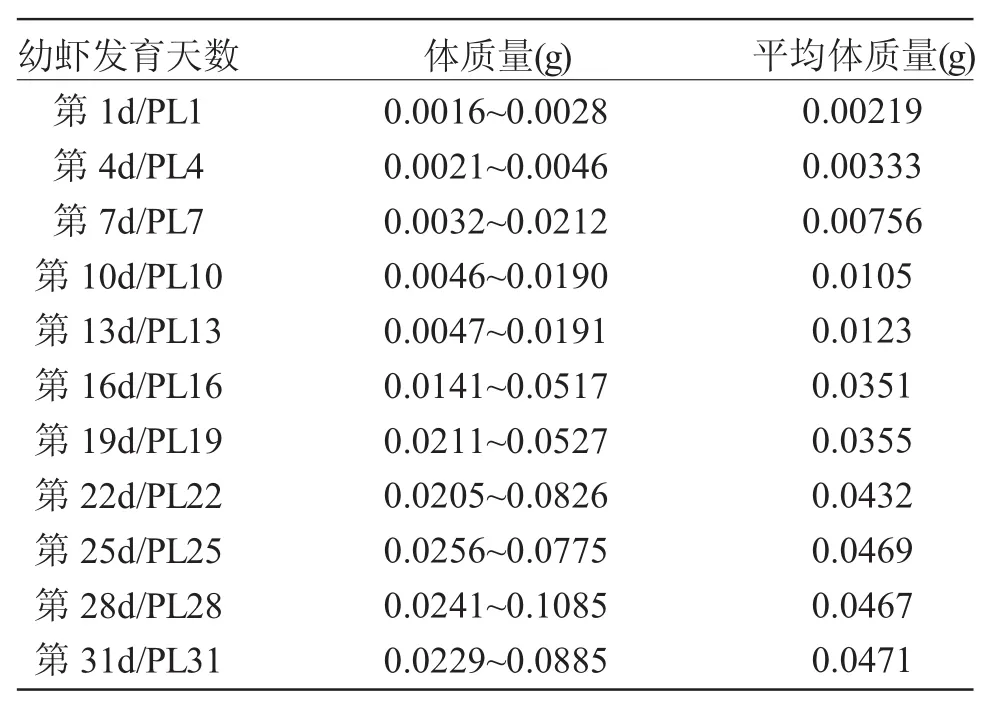
表1 組織學研究中實驗蝦的體質量Tab.1 The body weight of oriental river prawn(Macrobrachium nipponense)in histological observation
1.3 類固醇激素測定
從幼蝦發育第1~31d,每隔3d取樣一次,每次取10只用于測定類固醇激素。實驗蝦的體質量見表2。實驗蝦取樣后立即保存在-20℃環境中。用Beckman Coulter Access 2測定類固醇激素的分泌量。用甲醇提取睪酮及17β-雌二醇,用BECKMAN ACESSⅡT和E2試劑盒分別測定睪酮及17β-雌二醇的分泌量。
2 結果與分析
2.1 促雄腺的組織結構及變化
增殖期:在PL10,促雄腺開始形成而進入增殖期。此期,促雄腺體積較小,成索狀結構,形成少量促雄腺細胞。細胞體積小,直徑約為5.67~8.69μm。細胞核呈較強的嗜堿性(圖1-1)。
合成期:在PL13,促雄腺發育進入合成期。此期,促雄腺腺體及腺細胞均明顯增大,腺細胞呈圓形或橢圓形,直徑為12.98~17.67μm。腺細胞弱嗜堿性。腺體細胞已成典型的腺泡結構,血竇較為清晰(圖1-2)。
分泌期:在PL19,促雄腺發育進入分泌期。此期,促雄腺發育成熟,腺體呈泡狀化,含嗜堿性顆粒。腺細胞已不再有完整的細胞結構,核完全固化,體積很小。促雄腺與輸精管肌層的連接變得疏松,有從其表面脫落的趨勢,輸精管中充滿成熟的精子(圖1-3、圖1-4)。
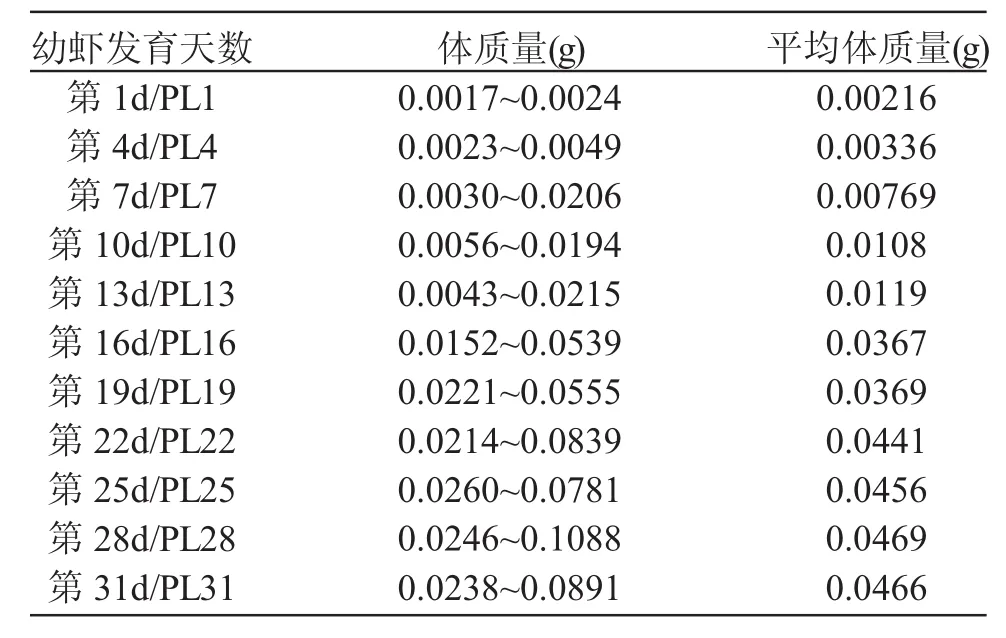
表2 測定類固醇激素含量實驗中青蝦的體質量Tab.2 The body weight of oriental river prawn(Macrobrachium nipponense)for steroid analysis
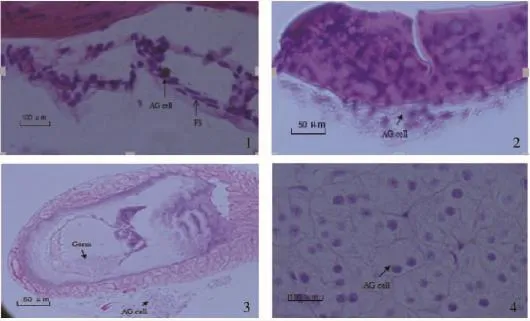
圖1 青蝦幼蝦發育過程中促雄腺的發育Fig.1 The development of androgenic gland in post-larva prawn
2.2 精巢的結構及變化
精原細胞期:在PL13,精巢發育進入精原細胞期。在這一時期,精原細胞開始逐漸形成。精巢體積很小,輸精管充滿了排列不規則的精原細胞(圖2-1)。
精母細胞期:在PL16,精巢發育進入精母細胞期。在這一時期,精巢及輸精管體積顯著增大。處于這一時期的精巢包含初級精母細胞、次級精母細胞及少量的精原細胞。大多數剩余的精原細胞正處于分化狀態(圖2-2)。
精細胞期:在PL19,精巢發育進入精細胞期。在這一時期,精巢體積進一步增大,仍可發現一些精母細胞及精原細胞,及少量成熟的精子(圖2-3)。生精小管繼續變大,營養物質散布在精細胞中。
精子期:在PL22,精巢發育進入精子期。在這一時期,精巢發育成熟,生精小管內充滿著成熟的精子,但腔內仍有一定量的精母細胞(圖2-4)。
退化期:在PL22,精巢發育進入退化期。此時,精巢發育停止,成熟的精子被排出。精巢進入新的發育周期,殘存的精原細胞開始恢復增殖(圖2-5)。
2.3 卵巢的組織結構及變化
卵原細胞期:在PL13,卵巢開始發育進入卵原細胞期。此時,生殖上皮分化出的卵原細胞呈橢圓或多邊形,細胞最大直徑為40.7~101.5μm。細胞核大,約占細胞體積的4/5。核中央有1個大而明顯的核仁。細胞質僅在核外一薄層(圖3-1)。
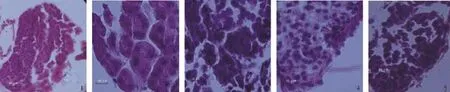
圖2 青蝦幼蝦發育過程中精巢的發育Fig.2 The development of testis in post-larva prawn
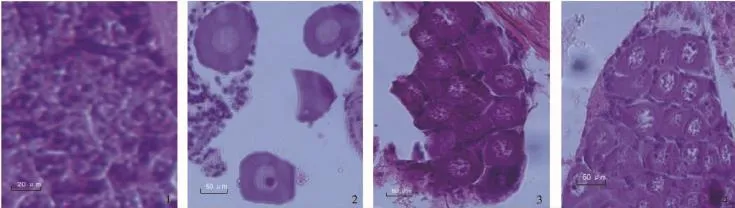
圖3 青蝦幼蝦發育過程中卵巢的發育Fig.3 The development of ovary in post-larva prawn
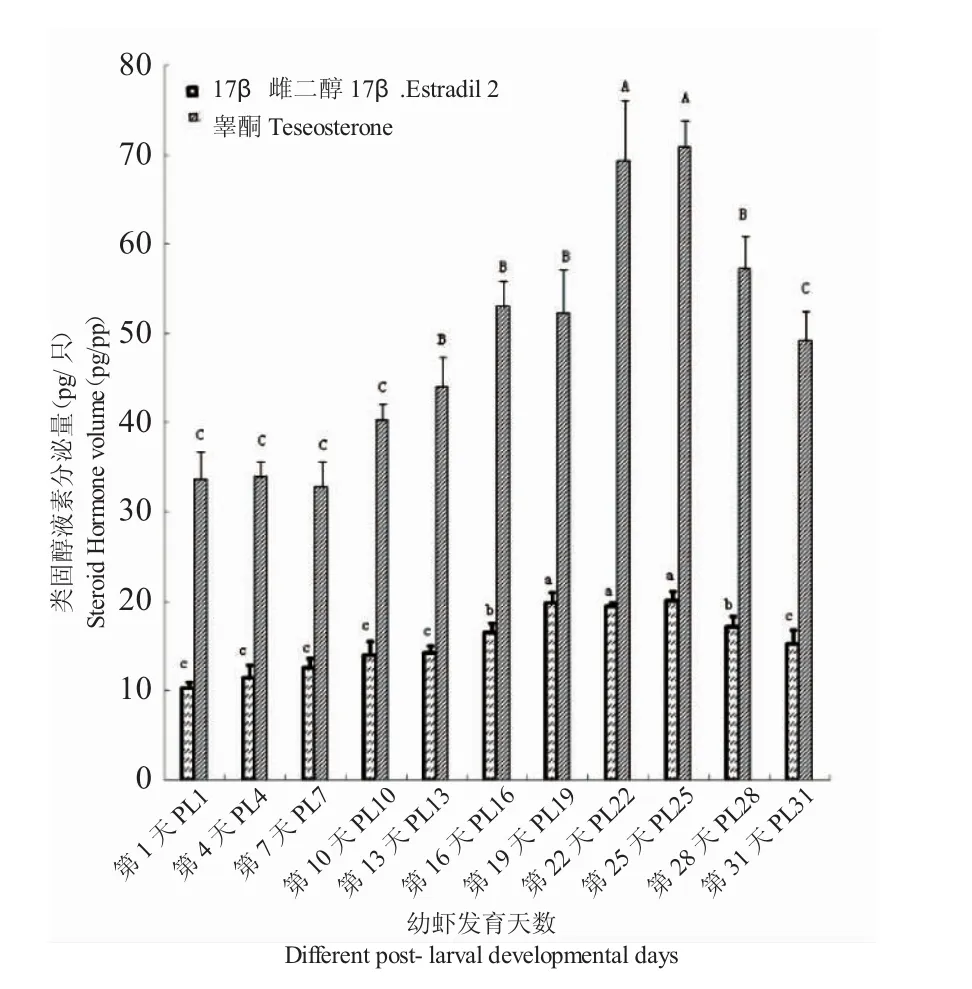
圖4 青蝦幼蝦發育期睪酮及17β-雌二醇的分泌量Fig.4 The contents of testosterone and 17β-estradiol in the post-larva prawn
初級卵母細胞期:在PL16,卵巢發育進入初級卵母細胞期。此時,卵原細胞體積逐漸變大,變為圓形或橢圓形,成初級卵母細胞,最大細胞直徑為55.7~129.1μm。核質比例相對減小,無卵黃積累,細胞質區增厚是營養物質開始增長的預兆(圖3-2)。
次級卵母細胞期:在PL16,卵巢發育進入次級卵母細胞期。此時,營養物質迅速積累,空泡內外已有卵黃粒積累。卵黃的堆積使得卵細胞體積大幅增加,最大細胞直徑可達69.5~164.7μm。細胞核位于細胞中央,圓形,有數個小核仁(圖3-3)。
卵子期:在PL19,卵巢發育進入卵子期。卵巢發育成熟并生成卵子。此時營養物進入成熟期。卵黃顆粒在空泡內外先后沉積,充塞了幾乎全部細胞,使得卵細胞相互擠壓呈多邊形,最大直徑為409.5~517.3μm。已不見細胞核(圖3-4)。
退化期:在PL19,卵巢進入退化期。此時,卵巢中成熟的卵子排出,剩余的卵原細胞回到初級卵母細胞期,并開始恢復生殖功能,卵巢進入新的發育周期。
2.4 睪酮與17β-雌二醇的分泌量
青蝦幼蝦發育過程中睪酮的分泌量從PL1的33.69pg/尾逐漸增加到PL22的69.27pg/尾,然后維持在穩定水平到PL25,之后開始逐漸下降;17β-雌二醇的分泌量從PL1的10.26pg/尾逐漸增加到PL19的19.77pg/尾,然后維持在穩定水平到PL25,之后開始逐漸下降(圖4)。統計學分析結果顯示:在性腺發育成熟期,睪酮與17β-雌二醇的分泌量顯著高于其他發育階段(P<0.05)。
3 討論
3.1 促雄腺發育研究
促雄性腺與性別分化、雄性發育及維持有關,在甲殼類性別調控中發揮著關鍵作用[2]。根據促雄腺發育的形態變化,可將甲殼動物的促雄腺分為不同的發育階段,如鯨螯蝦Orconectes nais的促雄腺分為4個發育時期[11],長額蝦Pandalus platyceros分為6個時期[12],中華絨螯蟹劃分為3個時期[13]。根據組織學觀察,青蝦促雄腺在PL10開始分化,隨后經歷增殖期(PL10)、合成期(PL13)和分泌期(PL19),在PL19發育成熟,形成完整的促雄腺結構。而在PL10時,性腺原基已發育成熟,可分化為精巢或卵巢。
3.2 精巢和卵巢發育的研究
青蝦精巢在PL13開始分化,隨后經歷精原細胞期(PL13)、精母細胞期(PL16)、精細胞期(PL19)和精子期(PL22)4個發育時期,在PL22發育成熟。促雄腺與精巢發育過程均持續10d,但促雄腺早于精巢3d開始發育和成熟,表明促雄腺的發育可調控精巢發育[5]。溪蟹Potamon koolooense的促雄腺和精巢發育與本文中所述的結果相似,當促雄腺最大時,精巢剛好啟動精子發生;當精子發育時,促雄腺逐漸縮小[14]。卵巢也從PL13開始分化,隨后經歷卵原細胞期(PL13)、初級卵母細胞期(PL16)、次級卵母細胞期(PL16)、卵子期(PL19),在PL19發育成熟。與其他的水產物種相似,青蝦精巢和卵巢經歷4個發育時期,直至成熟并退化[15-18]。測定睪酮及17β-雌二醇的分泌量已為多種水產動物的性腺發育提供了依據。本實驗中,睪酮的分泌量從PL1到PL22逐漸上升,并維持在穩定的分泌水平直到PL25,然后逐漸下降;然而,17β-雌二醇的分泌量從PL1~PL19逐漸上升,并維持在穩定的分泌水平直到PL25,然后逐漸下降。該結果與組織學觀察結果一致,證明精巢在PL22發育成熟,而卵巢在PL19發育成熟。類固醇激素在幼蝦發育后期分泌量下降表明,精巢和卵巢進入了新的發育周期。該結果說明,近親繁殖不但發生在新生的雌蝦與親本雄蝦之間,還可以發生在新生雄蝦與親本雌蝦之間,這嚴重制約著青蝦產業的可持續發展。Zhang等[19]報道,在28℃水溫條件下,青蝦在PL11可觀察到性腺原基,卵巢在PL18開始分化,而精巢在PL20開始分化。該結果顯示,溫度可顯著促進青蝦性腺的發育。
組織學觀察發現,同一天發育的幼蝦的性腺發育狀況差異極顯著。例如,在PL19實驗蝦中,體質量為0.0289g的幼蝦卵巢僅發育到初級卵母細胞期,而體質量為0.0466g及0.0517g的幼蝦卵巢則分別發育到次級卵母細胞期及卵子期。另外在PL22,體質量為0.0254g的幼蝦精巢僅發育到精母細胞期,而體質量為0.0756g及0.1117g的幼蝦精巢則分別發育到精細胞期及精子期。該結果表明,青蝦精巢和卵巢的發育狀況與其體長及體質量顯著相關。
[1]金舒博,王寧,喬慧,等.青蝦高血糖激素基因全長cDNA序列的克隆及表達分析[J].中國水產科學,2013,20(1): 1-11.
[2]Ventura T,Manor R,Aflalo E D,et al.Temporal silencing of an androgenic gland-specific insulin-like gene affecting phenotypical gender differences and spermatogenesis[J]. Endocrinology,2009,150(3):1278-1286.
[3]Ventura T and Sagi A.The insulin-like androgenic gland hormone in crustaceans:from a single gene silencing to a wide array of sexual manipulation-based biotechnologies[J].BiotechnologyAdvances,2012,6:1543-1550.
[4]Suzuki S and Yamasaki K.Sex-reversal of male Armadillidium vulgare(Isopoda Malaeostraea,Crustacea)following andrectomy and partial gonadectomy[J].General and Comparative Endocrinology,1991,83(3):375-378.
[5]吳萍,楊立榮,崇加榮,等.日本沼蝦促雄腺的研究[J].水利漁業,2002,22(5):21-23.
[6]趙光鳳,李廣麗,朱春華.凡納濱對蝦促雄性腺的組織學初步觀察[J].廣東海洋大學學報,2010,30(6):74-77.
[7]管衛兵,王桂忠,李少菁,等.鋸緣青蟹促雄腺和胸腹神經團的直接聯系[J].中國水產科學,2005,12(4):397-401.
[8]蘇青,朱冬發,楊濟芬,等.三疣梭子蟹促雄腺顯微和亞顯微結構的研究[J].水產科學,2010,29(4):193-197.
[9]肖美南,徐佳念,蔡生力,等.中華絨螯蟹促雄性腺素分離、純化的初步研究[J].上海海洋大學學報,2010,19(1):28-33.
[10]Qiao H,Xiong Y,Zhang W,et al.Characterization,expression,and function analysis of gonad-inhibiting hormone in oriental river prawn,Macrobrachium nipponense and its induced expression by temperature[J].Comparative Biochemistryand Physiology,Part A,2015,185:1-8.
[11]Carpenter M B and deRoos R.Seasonal morphology and histologyoftheandrogenicglandofthecrayfish,Orconectes nais[J].General Comparative Endocrinology,1970,15(1):143-157.
[12]Hoffman DL.The development ofthe androgenic glands of aprotandricshrimp[J].BiolBull,1969,137(2):286-296.
[13]邱高峰,吳萍,樓允東.中華絨螯蟹促雄腺的結構與功能[J].水產學報,2000,24(2):108-112.
[14]Joshi P C and Khanna S S.Studies on the androgenic gland ofthe freshwater crab,Potamon koolooense(Rathbum)[J].Zmikrosk-anat Forsch Leipzig,1987,101(4): 699-713.
[15]鄧道貴,胡艷玲.粗糙沼蝦精巢發育的組織學[J].動物學雜志,2002,37(3):62-65.
[16]羅宇良,吳志新,陳孝煊,等.紅螯螯蝦精巢發育的組織學研究[J].華中農業大學學報,1999,18(1):78-80.
[17]何緒剛,張訓蒲,龔世園,等.武湖日本沼蝦卵巢發育研究[J].華中農業大學學報,2002,21(2):148-151.
[18]馬丹丹,康現江,董麗君,等.中華絨螯蟹精巢發育組織學[J].水產科學,2006,25(6):291-296.
[19]ZhangY P,Sun S M,Fu H T,et al.Characterization ofthe male-specific lethal 3 gene in the oriental river prawn, Macrobrachium nipponense[J].Genetic Molecular Research,2015,14(2):3106-3120.
Histological Observation of Gonadal Development During Post-larva in Oriental River Prawn,Macrobrachium nipponense
JIN Shu-bo1,2,ZHANG Ying3,GUAN Hai-hong3,FU Hong-tuo1,2,JIANG Su-fei2,XIONG Yi-wei2, QIAO Hui2,ZHANG Wen-yi2,GONG Yong-sheng2,WU Yan2
(1.Wuxi Fisheries College,Nanjing Agricultural University,Wuxi 214081,China; 2.Key Laboratory of Freshwater Fisheries and Germplasm Resources Utilization,Ministry of Agriculture,Freshwater Fisheries Research Center,Chinese Academy of Fishery Sciences,Wuxi 214081,China; 3.Heilongjiang River Fisheries Research Institute,Chinese Academy of Fishery Sciences,Harbin 150070,China)
In this study,histological method and steroid hormone detection were used to analyze the beginning period,developmental process and mature period of gonad including testes,ovary,and androgenic gland during post-larvae of oriental river prawn(Macrobrachium nipponense).The histological analysis revealed that the initial development of funicular androgenic gland with a few androgenic gland cells was observed at post-larval stage of 10 days(PL10).The androgenic gland development was divided into 3 developmental stages:proliferating phase at PL10,synthesis phase at PL13 and secretary phase PL19,full maturity at PL19.Both small testes and ovary were found to begin to develop at PL13 when the spermatogonia with irregular arrangement were the first time observed. Oogonia differentiated from germinal epithelia were oval or polygon in shape.The testis development was divided into 4 stages including spermatogonium stage,spermatocyte stage,sperm cell stage and sperm stage and matured at PL22 when the seminiferous tubule of matured testis was filled with mature spermatozoa.The ovary development was divided into 4 stages including oogonia stage,primary oocyte stage,secondary oocyte stage and ovum stage,matured at PL19 when the matured ovary was full of ovum.The 17β-E2 levelwas shown to be gradually increased from 10.26 pg/prawn at PL1 to 19.77 pg/prawn at PL19,constant until PL25,and then the 17β-E2 level was gradually decreased.The level of 17α-T,however,was gradually increased from 33.69 pg/prawn at PL1 to 69.27 pg/prawn at PL22,constant until PL25,and then gradually decreased.Both androgenic gland and gonad were developed in 10 days,3 days earlier in development and maturity of androgenic gland than those in gonad.The findings provide information for the screening of the genes related to sexual differentiation and reproduction and for the mechanism of sex control during the developmental process.
Macrobrachium nipponense;histological observation;steroid hormone detection;gonadal development;sexual precocity
S954.48
A
1005-3832(2016)04-0011-06
2016-05-16
中央級科研院所基本科研業務費(2015JBFM11);國家自然科學基金(31502154).
金舒博(1986-),男,助理研究員,從事水產動物遺傳育種研究.E-mail:jinsb@ffrc.cn
傅洪拓(1964-),研究員,博士生導師,從事水產動物遺傳育種研究.E-mail:fuht@ffrc.cn
