野慈姑對吡嘧磺隆抗性的分子機(jī)理
2016-01-21 07:45:08劉華招高增貴
雜草學(xué)報 2015年3期
劉 延, 劉華招, 高增貴
(1.黑龍江省農(nóng)墾科學(xué)院植物保護(hù)研究所,黑龍江哈爾濱 150038;2.黑龍江省農(nóng)墾科學(xué)院水稻研究所,黑龍江哈爾濱 150038;
3.沈陽農(nóng)業(yè)大學(xué)植物免疫研究所,遼寧沈陽 110866)
?
野慈姑對吡嘧磺隆抗性的分子機(jī)理
劉延1, 劉華招2, 高增貴3
(1.黑龍江省農(nóng)墾科學(xué)院植物保護(hù)研究所,黑龍江哈爾濱 150038;2.黑龍江省農(nóng)墾科學(xué)院水稻研究所,黑龍江哈爾濱 150038;
3.沈陽農(nóng)業(yè)大學(xué)植物免疫研究所,遼寧沈陽 110866)
摘要:野慈姑是我國稻田危害最嚴(yán)重的雜草之一。以黑龍江稻田采集的野慈姑(Sagittaria trifolia Linn.)為研究對象,通過室內(nèi)生物測定和分子克隆技術(shù),對野慈姑的吡嘧磺隆抗性水平進(jìn)行檢測,并從靶標(biāo)位點(diǎn)突變的角度解釋抗性產(chǎn)生原因。結(jié)果表明,采自哈爾濱稻田的2個野慈姑種群N03及N06均為高抗種群,抗性達(dá)到4倍劑量以上。經(jīng)比對2個抗性種群的靶標(biāo)酶ALS基因發(fā)現(xiàn),Pro197的脯氨酸分別被亮氨酸、絲氨酸取代,該位點(diǎn)的突變可能是野慈姑種群對磺酰脲類除草劑產(chǎn)生抗藥性的主要原因。
關(guān)鍵詞:野慈姑;除草劑抗性;ALS基因;吡嘧磺隆

水稻是黑龍江省的主要糧食作物。2014年黑龍江省水稻種植面積突破340萬hm2,除北部高寒地區(qū)外,全省80個市(縣)中72個市(縣)種植水稻[1]。雜草是影響水稻產(chǎn)量的主要因素之一[2],隨著除草劑的大量使用,稻田雜草的種群變化和群落演替加速,原來處于次要防治地位的莎草科及澤瀉科雜草成為主要雜草。野慈姑(SagittariatrifoliaL.)別稱狹葉慈姑、長瓣慈姑,俗稱驢耳菜,為澤瀉科多年沼生草本植物。野慈姑喜溫暖濕潤環(huán)境,生長適宜溫度為20~25 ℃,既能種子繁殖也可宿生根繁殖[3-4]。通過田間調(diào)查和與農(nóng)民交流獲知,野慈姑近年來發(fā)生日益嚴(yán)重,己成為黑龍江省寒地稻區(qū)主要惡性雜草。
磺酰脲類除草劑芐嘧磺隆和吡嘧磺隆,自20世紀(jì)80年代末在我國登記以來,由于其具有活性高、用量低、殺草譜廣、對作物安全及毒性低等特點(diǎn),一直是水田防除闊葉雜草的主推品種[5]。我國東北地區(qū)水稻田使用磺酰脲類除草劑已有20多年歷史[6]。長期單一品種的除草劑施用極易誘導(dǎo)雜草產(chǎn)生抗藥性,造成野慈姑對磺酰脲類除草劑敏感性降低。日本于2014年報道了由于ALS基因突變而對芐嘧磺隆和吡嘧磺隆高抗的野慈姑種群[7]。國內(nèi)也有相關(guān)報道:吳明根等在吉林省延邊已發(fā)現(xiàn)抗磺酰脲類除草劑芐嘧磺隆生態(tài)型慈姑[8];陳麗麗等在黑龍江省慶安縣發(fā)現(xiàn)ED50比值4.5倍于敏感型的野慈姑[9]。筆者在試驗(yàn)區(qū)中也發(fā)現(xiàn)對芐嘧磺隆、吡嘧磺隆不敏感的野慈姑生態(tài)型。
世界范圍內(nèi)已有大量與ALS抑制劑抗性相關(guān)的報道。對于抗性機(jī)制的解釋主要有2個方面——靶標(biāo)位點(diǎn)抗性(target-site-resistance,TSR)和非靶標(biāo)位點(diǎn)抗性(non-target-site-resistance,NTSR)。目前已發(fā)現(xiàn)的151種ALS抑制劑抗藥性雜草中,經(jīng)證實(shí)與TSR直接相關(guān)的ALS基因突變位點(diǎn)共有8個[10-11]。根據(jù)擬南芥中ALS氨基酸的位置,8個位點(diǎn)分別為Ala122、Pro197、Ala205、Asp376、Arg377、Trp574、Ser653、Gly654。對于NTSR的研究主要集中在解毒和代謝2個方面[12]。
目前野慈姑抗性機(jī)制的解釋主要為編碼靶標(biāo)酶ALS基因發(fā)生突變。本研究以吡嘧磺隆為例,主要目的為探明黑龍江省野慈姑對磺酰脲類除草劑產(chǎn)生抗性的靶標(biāo)位點(diǎn)的抗性機(jī)制。
1材料與方法
1.1供試材料
采自黑龍江省哈爾濱市不同田塊的野慈姑種群(SagittariatrifoliaL.)見表1。各種群分別取植株幼嫩部分,凍存于-80 ℃。

表1 慈姑種群采集地點(diǎn)及用藥史
1.2試驗(yàn)方法
1.2.1整株生測法選取4~6葉期長勢均勻的野慈姑,進(jìn)行噴霧處理。試驗(yàn)按照表2設(shè)計用量處理,每處理重復(fù)4次。噴藥后放在暗處吸收6 h,然后光暗交替12 h繼續(xù)培養(yǎng)。
1.2.2野慈姑ALS基因擴(kuò)增引物的設(shè)計與合成根據(jù)已發(fā)表的野慈姑ALS基因序列(AB301496.1),設(shè)計并于生工生物工程(上海)股份有限公司合成正反向2條引物(表3)。
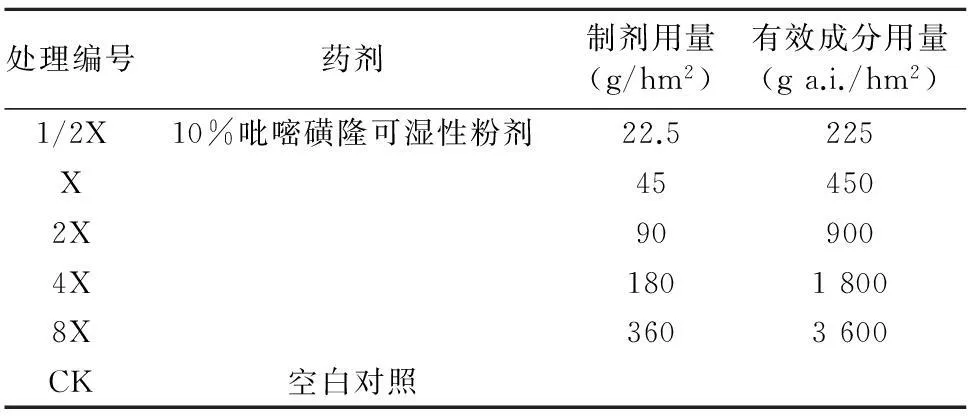
表2 吡嘧磺隆試驗(yàn)用量設(shè)計

表3 擴(kuò)增野慈姑ALS基因的引物
1.2.3野慈姑ALS基因片段的克隆及測序?qū)δ康臈l帶切膠回收、純化后連接到pMD18-T載體,然后轉(zhuǎn)到大腸桿菌感受態(tài)細(xì)胞中。將菌液均勻涂布于含有Ampicilin和X-gal/IPTG的LB固體培養(yǎng)基上,37 ℃過夜培養(yǎng)。從培養(yǎng)基上挑取15個白色單菌落到含Ampicilin的LB培養(yǎng)液中,以200 r/min的速度在37 ℃搖床內(nèi)振蕩培養(yǎng)5 h,然后以菌液為模板進(jìn)行轉(zhuǎn)化鑒定。25 μL的菌液PCR組分包括9.5 μL ddH2O、12.5 μL 2×Pfu PCR MasterMix(TIANGEN公司)、1 μL菌液、上下游引物各1 μL(10 μmol/L)。挑選其中至少5個陽性克隆送公司雙向測序,引物為通用引物M13。
1.2.4生物學(xué)信息分析用DNAMAN V6軟件對試驗(yàn)反枝莧種群ALS基因片段的測序結(jié)果進(jìn)行比對,尋找6個突變位點(diǎn)的差異性。
2結(jié)果與分析
2.1生物測定結(jié)果及分析
施藥7 d后,調(diào)查吡嘧磺隆對各野慈姑樣本地上鮮質(zhì)量的抑制差異。隨吡嘧磺隆劑量的升高,對8個樣本的抑制率增強(qiáng)。其中樣本N03和N06抗性水平較高,4倍劑量時的生長抑制率低于50%,即對吡嘧磺隆抗性達(dá)到4倍劑量以上(表4)。對這2個野慈姑抗性種群和敏感種群進(jìn)行ALS基因片段的生物信息學(xué)分析。
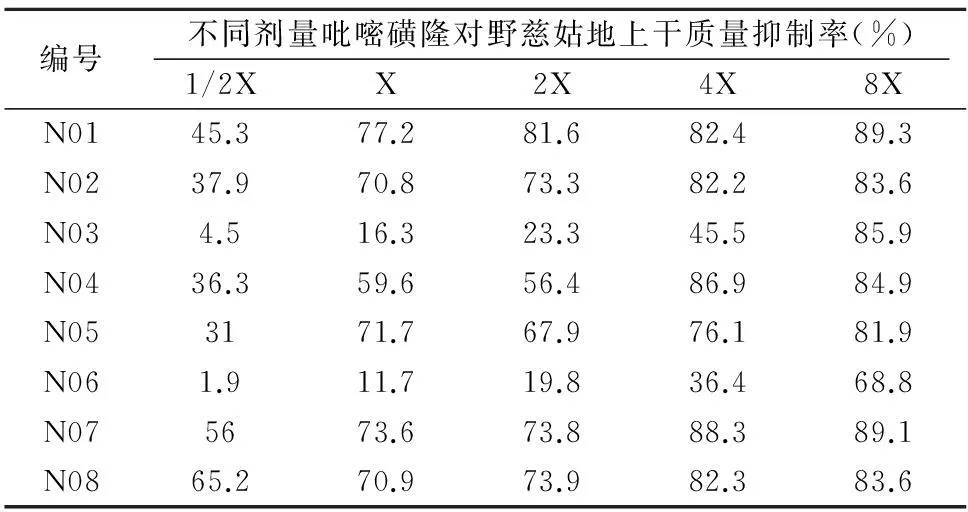
表4 吡嘧磺隆對野慈姑地上干質(zhì)量的影響
2.2野慈姑基因組DNA的提取及目的片段擴(kuò)增
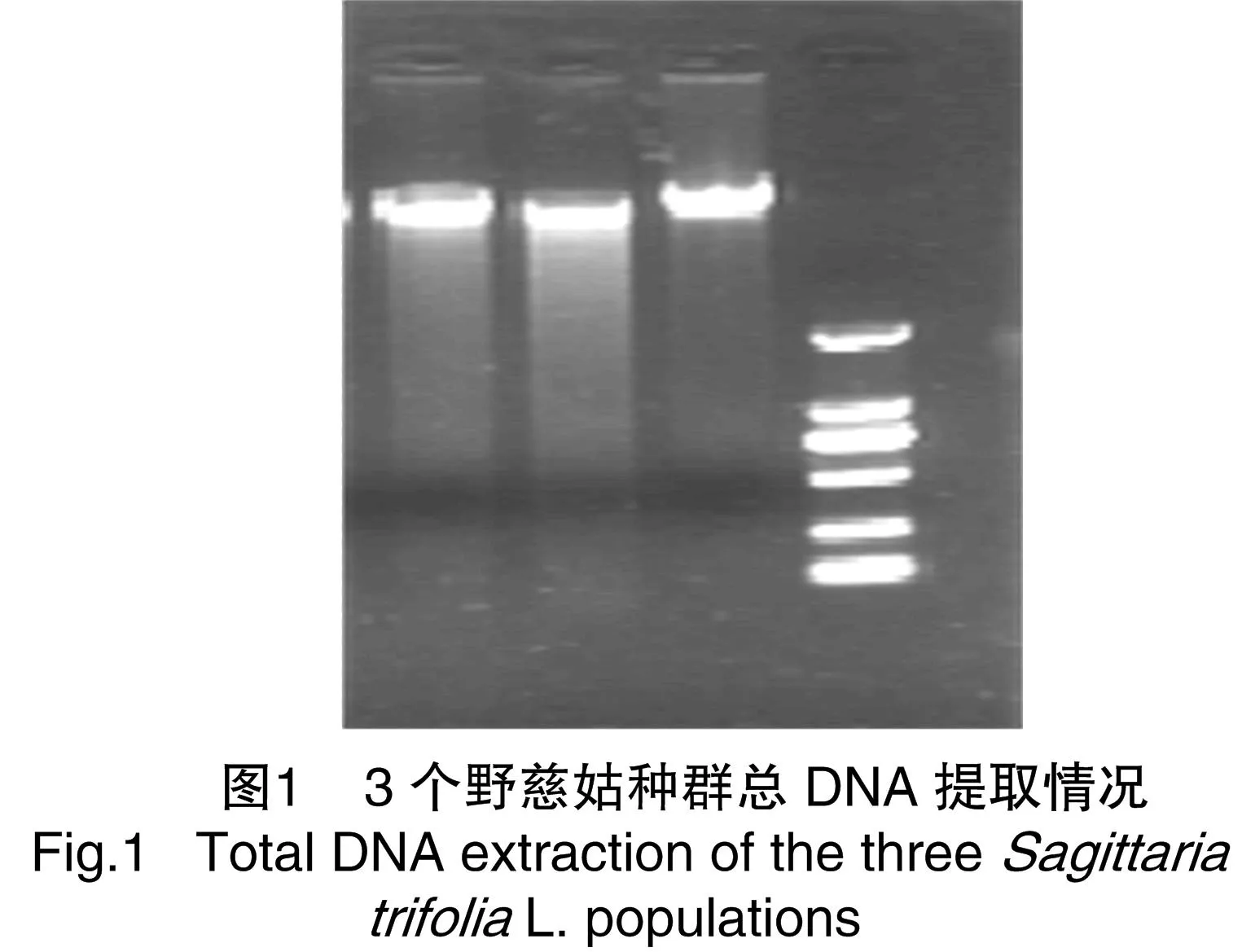
3種野慈姑取幼嫩組織在液氮中研磨成細(xì)粉,使用快捷型植物基因組DNA提取系統(tǒng),對基因組DNA進(jìn)行提取,電泳檢測提取成功(圖1)。
PCR反應(yīng)條件為94 ℃預(yù)變性5 min;94 ℃變性30 s、59 ℃退火30 s、72 ℃延伸2 min,共進(jìn)行33 次循環(huán);72 ℃延伸10 min。擴(kuò)增產(chǎn)物點(diǎn)樣于1%瓊脂糖凝膠中進(jìn)行電泳檢測,擴(kuò)增成功(圖2)。
2.3野慈姑ALS基因片段的生物信息學(xué)分析
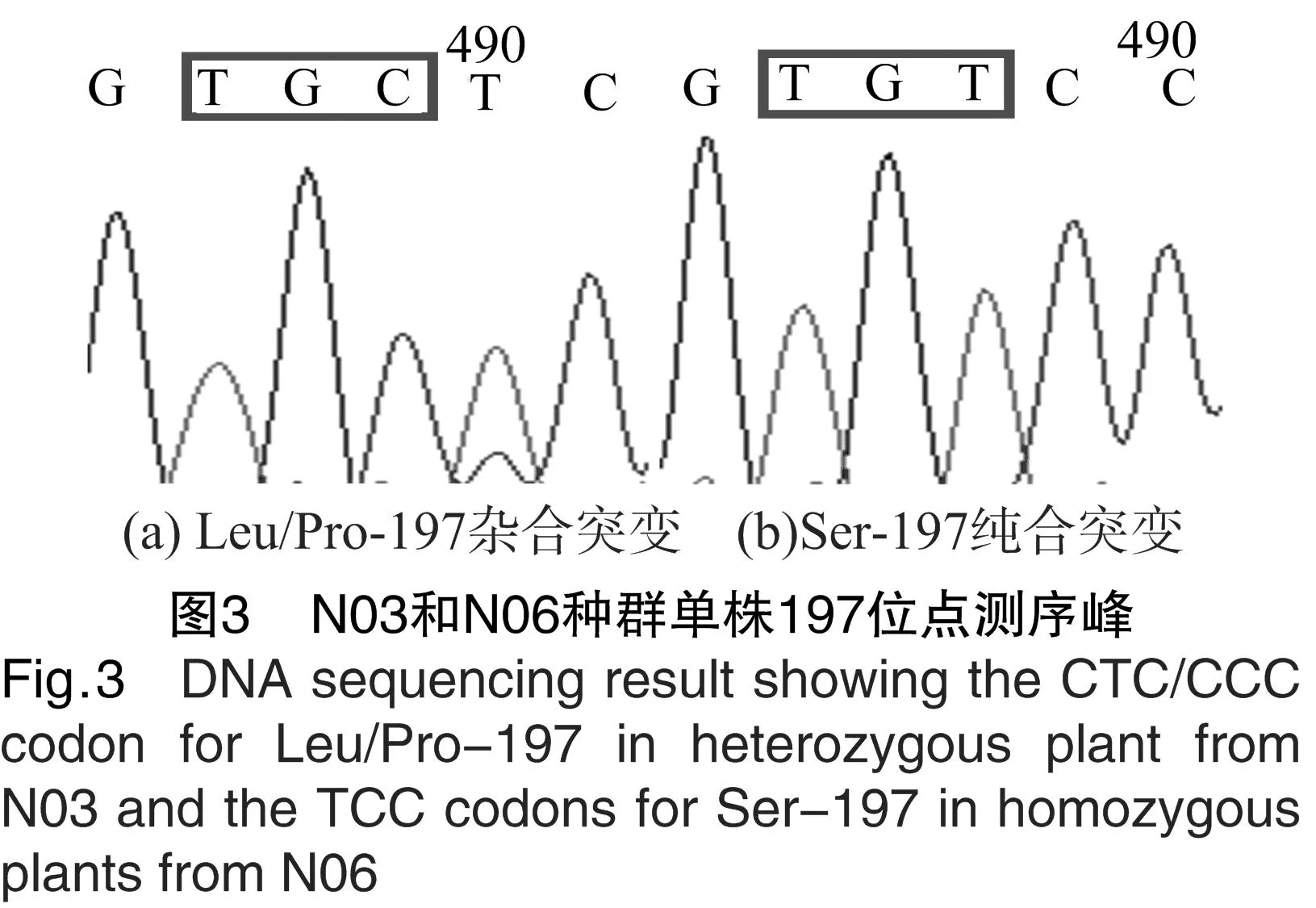
對3個野慈姑種群的ALS測序片段進(jìn)行比對后發(fā)現(xiàn),抗性種群N03和N06在Pro197位發(fā)生了導(dǎo)致不同氨基酸發(fā)生取代的突變。其中,N03由CCC突變?yōu)镃TC,亮氨酸取代了脯氨酸;N06由CCC突變?yōu)門CC,絲氨酸取代了脯氨酸(表5)。從測序峰圖(圖3)上來看,N03種群的單株在編碼Ser的第2個堿基處出現(xiàn)了套峰,N06種群的單株無套峰出現(xiàn)。由于野慈姑為二倍體[13],因此峰圖表明,N03的197位點(diǎn)突變僅發(fā)生在野慈姑的1條染色體上,即N03種群野慈姑單株P(guān)ro197Pro/Leu為雜合突變,而N06種群野慈姑單株P(guān)ro197Ser為純合突變。
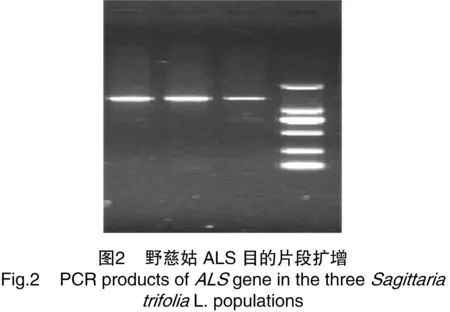
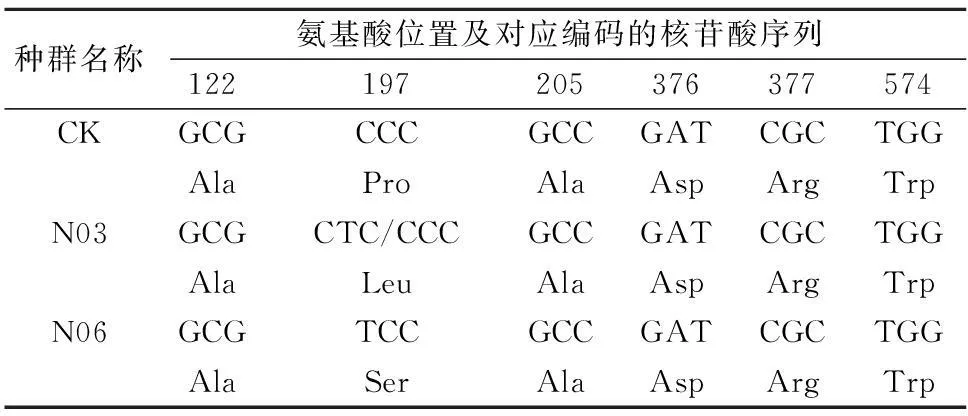
表5 3個野慈姑種群ALS基因片段及編碼氨基酸的比較
3討論與結(jié)論
本試驗(yàn)通過整株生物測定法,對采自8個地塊野慈姑的抗藥性水平進(jìn)行了檢測,發(fā)現(xiàn)有2個地塊的野慈姑對吡嘧磺隆的敏感性降低,抗性達(dá)到4倍劑量以上。對靶標(biāo)酶ALS的部分基因片段進(jìn)行擴(kuò)增、克隆、測序后發(fā)現(xiàn),2個抗性種群——N03、N06靶標(biāo)酶ALS的高度保守區(qū)域Domain A發(fā)生了基因突變,197位由CCC分別突變?yōu)镃TC/CCC、TCC,分別導(dǎo)致脯氨酸被亮氨酸、絲氨酸取代。突變的發(fā)生可能是導(dǎo)致野慈姑對磺酰脲類除草劑產(chǎn)生抗性的重要原因之一。
基于靶標(biāo)位點(diǎn)突變的ALS抑制劑抗性是雜草產(chǎn)生抗性的重要機(jī)制,通常是由于ALS的編碼基因保守區(qū)域發(fā)生突變,導(dǎo)致單個氨基酸被取代[14-15],這種取代也可在多個位點(diǎn)同時發(fā)生。在過去20年里,全球范圍內(nèi)在50個雜草種群中,共發(fā)現(xiàn)8個能夠?qū)е驴剐援a(chǎn)生的突變位點(diǎn)以及26種氨基酸替代方式[16]。目前野慈姑的靶標(biāo)位點(diǎn)抗性情況已有報道,2014年日本Iwakami等發(fā)現(xiàn)了發(fā)生CCC/TCC雜合突變的野慈姑種群,本研究發(fā)現(xiàn)的2種突變方式尚未報道。
非靶標(biāo)位點(diǎn)導(dǎo)致的除草劑抗性原理較復(fù)雜,可能由除草劑代謝機(jī)制的增強(qiáng)或解毒酶(P450解毒酶,GSTs等)活性提高引起[12,17]。本次試驗(yàn)未對非靶標(biāo)方面機(jī)理進(jìn)行研究,因此不能排除其對抗性產(chǎn)生的貢獻(xiàn),這有待進(jìn)一步證實(shí)。
參考文獻(xiàn):
[1]關(guān)昕,安龍哲. 黑龍江省水稻生產(chǎn)概況[J]. 農(nóng)機(jī)使用與維修,2013(1):10-12.
[2]Florez J A,F(xiàn)ischer A J,Ramirez H,et al. Predicting rice yield losses caused by multispecies weed competition[J]. Agronomy Journal,1999,91(1):87-92.
[3]王枝榮. 中國農(nóng)田雜草原色圖譜[M]. 北京:農(nóng)業(yè)出版社,1990:357.
[4]余柳青,韓逢春,玄松南,等. 東北水稻生產(chǎn)與雜草防除[J]. 雜草科學(xué),2009(4):7-10.
[5]李茹,羅小娟,董立堯,等. 鱧腸種群對除草劑的敏感性[J]. 江蘇農(nóng)業(yè)學(xué)報,2013,29(6):1514-1516.
[6]吳明根,鄭承志,李昕珈. 抗芐嘧磺隆慈姑ALS基因突變位點(diǎn)[J]. 江蘇農(nóng)業(yè)學(xué)報,2010,26(1):222-224.
[7]Iwakami S,Watanabe H,Miura T,et al. Occurrence of sulfonylurea resistance inSagittariatrifolia,a basal monocot species,based on target-site and non-target-site resistance[J]. Weed Biology and Management,2014,14(1):43-49.
[8]吳明根,吳松權(quán),樸仁哲,等. 磺酰脲類除草劑對抗、感性慈姑ALS活性的影響[J]. 農(nóng)藥,2007,46(10):701-703.
[9]陳麗麗,何付麗,范丹丹,等. 黑龍江省野慈姑對吡嘧磺隆的敏感性測定[J]. 植物保護(hù),2013,39(6):120-123.
[10]Beckie H J,Tardif F J. Herbicide cross resistance in weeds[J]. Crop Protection,2012,35:15-28.
[11]Powles S B,Yu Q. Evolution in action:plants resistant to herbicides[J]. Annual Review of Plant Biology,2010,61:317-347.
[12]Yu Q,Powles S. Metabolism-based herbicide resistance and cross-resistance in crop weeds:A threat to herbicide sustainability and global crop production[J]. Plant physiology,2014,166(3):1106-1118.
[13]李林初. 澤瀉和慈姑核型的比較研究[J]. 植物科學(xué)學(xué)報,1985,3(4):397-402.
[14]Rajguru S N,Burgos N R,Shivrain V K,et al. Mutations in the red rice ALS gene associated with resistance to imazethapyr[J]. Weed science,2005,53(5):567-577.
[15]Shaner D L. Resistance to acetolactate synthase (ALS) inhibitors in the United States:history,occurrence,detection,and management[J]. Weed Research,1999,44(4):405-411.
[16]Yu Q,Powles S B. Resistance to AHAS inhibitor herbicides:current understanding[J]. Pest Management Science,2014,70(9):1340-1350.
[17]Vila-Aiub M M,Neve P,Powles S B. Resistance cost of a cytochrome P450 herbicide metabolism mechanism but not an ACCase target site mutation in a multiple resistantLoliumrigidumpopulation[J]. New Phytologist,2005,167(3):787-796.
鄒紅梅,李香菊,崔海蘭,等. 菵草對3種ACCase抑制劑抗性的瓊脂快速檢測方法建立[J]. 雜草科學(xué),2015,33(3):24-28.
Molecular Resistance Mechanism ofSagittariatrifoliaL.
to Pyrazosulfuron-methyl
LIU Yan1, LIU Hua-zhao2, GAO Zeng-gui3
(1. Plant Protection Research Institute,Heilongiiang Academy of Land Reclamation Sciences,Harbin 150038,China;
2. Rice Research Institute,Heilongiiang Academy of Land Reclamation Sciences,Harbin 150038,China;
3. Institute of Plant Immunity,Shenyang Agricultural University,Shenyang 110866,China)
Abstract:Sagittaria trifolia L. is one of the most serious weeds in paddy fields in China. In this study,two S. trifolia populations collected from Heilongjiang Province were found highly resistant (resistance indexes above 4.0) to pyrazosulfuron-methyl in dose-response curves. The resistance mechanism was elucidated through amplifycation and cloning sections of the ALS gene. Different mutations were found in Pro197 in the two resistant populations. In one of the populations (N03),CCC was changed to CTC/CCC at 197,leading to the substitution of Pro by Leu/Pro. Population N06 possessed a TCC to CCC mutation,resulting in a Ser-197-Pro substitution. These mutations may be responsible for the resistance mechanism.
Key words:Sagittaria trifolia L.;herbicide resistance;ALS gene;pyrazosulfuron-methyl
作者簡介:劉延(1978—),女,博士,副研究員,主要研究方向?yàn)殡s草抗藥性與農(nóng)田雜草防控。E-mail:liuyanzb1226@163.com。
基金項(xiàng)目:國家公益性行業(yè)(農(nóng)業(yè))科研專項(xiàng)(編號:201203098);黑龍江省農(nóng)墾科學(xué)院2011年院長基金。
收稿日期:2015-03-20
中圖分類號:S451.21
文獻(xiàn)標(biāo)志碼:A
文章編號:1003-935X(2015)03-0020-04
