上皮鈣黏素/連環素復合體對大鼠神經干細胞增殖作用的影響
2015-12-31 00:53:06顏建輝,黃麗娟,楊柳等
中國老年學雜志 2015年5期
上皮鈣黏素/連環素復合體對大鼠神經干細胞增殖作用的影響
顏建輝1黃麗娟1楊柳1陳雁斌1華力明1
(湘南學院心腦血管研究所,湖南長沙410000)
摘要〔〕目的探討E-鈣黏素/連環素(E-cadherin/catenin)復合體對神經干細胞(NSCs)增殖作用的影響。方法從胎鼠腦組織中分離、培養NSCs,制備細胞懸液,隨機分為正常組、實驗1組、實驗2組。克隆大鼠E-cadherin全長于慢病毒(Lentivirus),轉染至實驗1組NSCs;采用siRNA法將E-cadherin對應片段克隆進入Lentivirus載體,轉染至實驗2組。RT-PCR及Western印跡方法檢測各組細胞E-cadherin、β-catenin、p120-ctn的mRNA及蛋白表達水平。以MTT法繪制細胞生長曲線,免疫熒光染色標記計數Nestin、BrdU 陽性細胞數目。結果實驗1組E-cadherin、β-catenin、p120-ctn的mRNA及蛋白表達水平均顯著高于正常組,但NSCs增長率及BrdU 陽性細胞數目顯著低于正常組(P<0.05);實驗2組E-cadherin、β-catenin、p120-ctn的mRNA及蛋白表達水平顯著低于正常組,但NSCs增長率及Nestin、BrdU 陽性細胞數目均顯著高于正常組(P<0.05)。結論下調E-cadherin/catenin復合體的表達可能促進體外培養的NSCs增殖。
關鍵詞〔〕E-鈣黏素/連環素;神經干細胞;慢病毒
中圖分類號〔〕R-36〔文獻標識碼〕A〔
1湘南學院附屬醫院
第一作者:顏建輝(1974-),男,副教授,副主任醫師,主要從事腦血管病變研究。
神經干細胞(NSCs)是一類具有分化為神經元細胞,并能夠提供大量腦組織細胞能力的干細胞,因NSCs的自我更新、多向分化潛能的性質,被譽為“萬能細胞”,在中樞神經系統受損的多種疾病中都有重要的治療價值。上皮鈣黏蛋白(E-cadherin)是一種重要的細胞間黏附分子,參與形成和維護正常細胞間的連接〔1〕。E-cadherin功能的發揮離不開連環素(catenin)的作用,它可以與α-catenin、β-catenin、γ-catenin及p120-catenin一起組成E-cadherin/catenin 復合體,在維持上皮細胞的組織結構完整性和新陳代謝過程中起到重要作用。p120ctn是穩定E-cadherin/catenin復合體的主要成分,最新研究表明,E-cadherin與p120-catenin結合,可以調控人胚胎干細胞的微環境以及重編程功能〔2〕。在小鼠的胚胎干細胞中研究發現,E-cadherin/β-catenin介導細胞與細胞間的黏附,同時可以抑制GSK-3β活性,調控干細胞的自我更新活性〔3〕。但是,E-cadherin/catenin復合體對NSCs的增殖作用鮮有報道。本課題觀察E-cadherin/catenin復合體內黏附分子變化對移植NSCs的增殖的影響,為提高NSCs移植治療相關疾病的臨床療效及新藥的開發提供實驗依據。
1材料與方法
1.1動物孕14 d Wistar大鼠1只,由武漢大學動物實驗中心提供。
1.2主要試劑多聚賴氨酸、巢蛋白(nestin)、5-溴脫氧尿嘧啶核苷(Brdu)、Lipofectamine 2000均由Sigma公司提供; B27復合物、胎牛血清、堿性成纖維細胞生長因子(bFGF)、表皮細胞生長因子(EGF)由北京中杉金橋試劑有限公司提供; DMEM/F12基礎培養基、Trizol試劑盒、RNA 抽提試劑盒、熒光標記FITC試劑、DAB顯色劑、SABC免疫組化試劑盒均由武漢子心生物科技有限公司提供;兔抗山羊IgG-Cy3,山羊抗鼠IgG-FITC均由武漢博士德生物有限公司提供;E-cadherin單克隆抗體及p120-catenin抗體均由Abcam公司提供;Lentivirus病毒由上海吉滿生物科技公司提供。
1.3方法
1.3.1NSCs的分離與培養取孕14 d Wistar大鼠,脫臼處死后于無菌條件下取出胎鼠,取出大腦組織D-Hank漂洗后,剪碎,吹打分散制成細胞懸液,離心,棄上清,置于含有2%B27,20 μg/L bFGF的DMEM/F12培養基中,在CO2培養箱中懸浮培養,3 d后待克隆球形成后,換液,用同樣的方法傳代培養3~4次,收集懸浮神經球制備單細胞懸液,調整濃度至1×105/ml。
1.3.2實驗分組將上述NSCs單細胞懸液等分為3份,分別為正常組、實驗1組及實驗2組。正常組不作任何處理,繼續培養。
1.3.3E-cadherin克隆及過表達 克隆大鼠E-cadherin編碼序列全長,表達于Lentivirus病毒載體,經293T細胞包裝后,收集病毒,轉染至實驗2組的NSCs,具體轉染步驟按Lipofectamine 2000試劑說明書操作。
1.3.4siRNA-Lentivirus病毒載體構建及轉染根據siRNA的設計原則,使用Invitrogen公司siRNA設計軟件,載體pGenesil-的酶切位點以及GenBank中大鼠E-cadherin(GenBank accession AB017696.1)的編碼序列,設計并合成片段如下:siRNA:5'-AAGATAGGAGTTCTCTGATGC-3' 陰性序列:5'-GGCGUGUCUCUCUUACGAC-3'。序列采用慢病毒(Lentivirus)載體,將對應片段克隆進入載體,經293T細胞包裝后,收集病毒,轉染至實驗1組的NSCs,操作同上。
1.4MTT法測定NSCs的生長曲線取上述各組NSCs懸液,以無血清培養基接種于96孔培養板(細胞濃度1×104個),150 μl/孔,觀察記錄細胞的生長狀況,在每孔加入MTT(5 g/L)20 μl,培養4h,手機孔內細胞并離心,每孔加入150 μlDMSO,震蕩10 min,并分別于1、2、3、4、5、6和7 d以酶標儀自動測定490 nm波長時OD值。
1.5RT-PCR檢測E-cadherin、β-catenin 及p120-ctn 的mRNA的表達Trizol提取總RNA,反轉錄為cDNA,再以cDNA為模板行PCR 擴增,各基因的引物序列,見表1。PCR 產物以2%瓊脂糖凝膠電泳測定擴增帶灰度值,得到mRNA的相對表達量,以β-actin為參照。
表1各基因引物序列及擴增片段長度

基因引物序列(5'→3')擴增片段長度(bp)E-cadherin正義鏈:TGCCCCAGTATCGTCCCCGT186反義鏈:CGGTTGCCCCATTCGT-TCAGATAAp120-ctn正義鏈:TGCCCTGCTGGATTTGTCTT250反義鏈:CGCTGGGTGTCCTGATGTGCβ-catenin正義鏈:GACTCTGAGAAACTTGTC-CGATGC375反義鏈:CGCTGGGTGTCCTGATGTGCβ-actin正義鏈:GAAATCGTGCGTGACATTAAG665反義鏈:CTAGAAGCATTTGCGGTGGA
1.6Western印跡法檢測E-cadherin、β-catenin 及p120-ctn的蛋白表達各組NSCs懸液,裂解、變性后離心,取上清液,以Bradford測定總蛋白量。取10 μg蛋白上樣,采用SDS-PAGE凝膠電泳后,轉印至PVDF膜;然后將膜室溫下封閉2 h,PBS漂洗,分別加入一抗E-cadherin(1∶400)、β-catenin(1∶200)及p120-ctn(1∶400) ,4℃過夜;辣根過氧化物酶標記二抗37℃孵育1 h,ECL發光顯色,以圖像分析系統觀察表達情況,以β-actin作為對照。
1.7Nestin、BrdU免疫熒光染色及計數各組NSCs接種于無菌蓋玻片的24孔培養板中(預先包被0.01%多聚賴氨酸),于37℃,5%CO2培養箱中培養,收集貼壁細胞制備細胞爬片,4%多聚甲醛固定,PBS沖洗,加一抗Nestin(1∶200)、BrdU(1∶200)進行免疫熒光染色,隨機觀察100個視野,熒光顯微鏡下計數每個視野的陽性細胞數,并取其平均值。
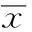
2結果
2.1各組E-cadherin、β-catenin 及p120-ctn的表達Western印跡和RT-PCR法顯示,實驗1組E-cadherin蛋白及mRNA表達較正常組則顯著增強(P<0.05),呈過表達狀態,β-catenin 及p120-ctn蛋白及mRNA表達也顯著增強(P<0.05);而經過siRNA干擾后,實驗2組的E-cadherin的蛋白及mRNA表達均較正常組顯著減弱,差異均有統計學意義(P<0.05),β-catenin 及p120-ctn的蛋白及mRNA表達較正常組也顯著減弱(P<0.05),見圖1,圖2。

A、B、C:正常組、實驗2組及實驗1組,下圖同 圖1 E-cadherin、β-catenin及p120-ctn的蛋白表達

圖2 E-cadherin、β-catenin 及p120-ctn的mRNA表達
2.2各組NSCs的生長曲線實驗1組的OD值與正常組相比顯著降低(P<0.05),而實驗2組的OD值則顯著高于正常組及實驗1組(P<0.05),見圖3。

圖3 各組NSCs的生長情況
2.3免疫熒光測定各組Nestin及BrdU陽性細胞數目BrdU 熒光染色陽性細胞胞質著紅色,Nestin染色陽性細胞則呈綠色;與正常組相比,實驗1組的Nestin及BrdU陽性細胞數目顯著減少(P<0.05),而實驗2組的Nestin及BrdU陽性細胞數目則顯著高于正常組及實驗1組,差異有統計學意義(P<0.05),見表2及圖4,圖5。


組別Nestin陽性細胞BrdU陽性細胞正常組43.28±12.4532.15±10.64實驗1組20.07±9.131)14.22±8.931)實驗2組72.63±13.011)2)61.37±11.061)2)
與正常組比較:1)P<0.01;與實驗1組比較:2)P<0.01

正常組 實驗1組 實驗2組 圖4 BrdU陽性細胞數目(免疫熒光×400)

正常組 實驗1組 實驗2組 圖5 Nestin陽性細胞數目(免疫熒光,×400)
3討論
NSCs能夠生成神經組織,在體外能維持未分化狀態,因其具有多向分化潛能和自我更新能力〔4,5〕,故而NSCs移植為腦損傷修復以及神經性疾病的治療帶來了希望。近年來,隨著人們對影響NSCs增殖的不斷深入研究,發現微環境對于干細胞的自我更新,增殖,分化調控以及干細胞遷移和歸巢至關重要,而微環境中E-cadherin是其中重要組成部分。研究發現E-cadherin介導的細胞與細胞間連接,可以促進小鼠胚胎干細胞自我更新,且不依賴LIF因子,這表明E-cadherin在干細胞的自我更新中扮演關鍵性角色〔6〕。catenin是一類位于細胞質內結構相似的糖蛋白家族,也是E-cadherin最為密切的細胞質內黏附相關蛋白配體,E-cadherin黏附功能的正常發揮,主要受catenin的調節。該家族中主要包括α-catenin、β-catenin、γ-catenin及p120-catenin。當胞質內α-catenin將β-catenin和(或)γ-catenin與細胞骨架的肌動蛋白微絲相連,而β-catenin和(或)γ-catenin
直接與E-cadherin 的胞質區結合,即形成了E-cadherin/ catenin 復合體。該復合體能將E-cadherin定位于細胞與細胞間的接觸部位,在細胞間黏附、建立細胞極性、維持組織形態中起重要作用。P120ctn 是穩定E-cadherin/catenin復合體的主要成分〔7〕。大量研究表明,腫瘤組織中如果E-cadherin/catenin復合體出現破壞或丟失的現象,細胞間黏附將會丟失,有利于惡性細胞的無限制增生〔8〕。而上皮細胞內E-cadherin過度表達能抑制細胞的游走與增殖,誘導細胞凋亡〔9〕。因此,推測如果下調E-cadherin/catenin復合體的表達,可能會促進體外培養NSCs的增殖。
本實驗結果提示隨著E-cadherin、β-catenin 及p120-ctn表達的下降,導致E-cadherin與catenin連接受阻,E-cadherin/catenin復合體與細胞骨架相互作用減弱,使E-cadherin不能定位于膜上,被胞質內蛋白酶降解,細胞的黏附功能障礙,細胞間連接松散,黏附力下降,有利于傳遞增殖信號導,細胞分裂增殖能力增強,從而促進了NSCs的增殖。
綜上所述,下調E-cadherin/catenin復合體的表達可能促進體外培養的NSCs增殖,為NSCs治療中樞神經系統疾病提供了新的視點,但是其具體作用機制尚需要更深入的研究。
4參考文獻
1Zhang L,Ju X,Cheng Y,etal.Identifying Tmem59 related gene regulatory network of mouse neural stem cell from a compendium of expression profiles〔J〕.BMC Syst Biol,2011;29(5):152.
2Li D,Zhou J,Wang L,etal.Integrated biochemical and mechanical signals regulate multifaceted human embryonic stem cell functions〔J〕.J Cell Biol,2010;191:631-44.
3Sineva GS,Pospelov VA.Inhibition of GSK3beta enhances both adhesive and signalling activities of beta-catenin in mouse embryonic stem cells〔J〕.Biol Cell,2010;102:549-60.
4Hong BH,Ha S,Joo Y,etal.Amyloid precursor protein binding protein-1 knockdown reduces neuronal differentiation in fetal neural stem cells〔J〕.Neuroreport,2012;23(2):61-6.
5Crockett S,Clarke M,Reeves S,etal.Cystine glutamate exchanger upregulation by retinoic acid induces neuroprotection in neural stem cells〔J〕.Neuroreport,2011;22(12):598-602.
6Soncin F,Mohamet L,Eckardt D,etal.Abrogation of E-cadherin-mediated cell-cell contact in mouse embryonic stem cells results in reversible LIF-independent self-renewal〔J〕.Stem Cell,2009;27(9):2069-80.
7Berx G,van Roy F.Involvement of members of the cadherin super-family in cancer〔J〕.Cold Spring Harb Perspect Biol,2009;1(6):a003129.
8Brooks SA,Lomax Browne HJ,Carter TM ,etal.Molecular interact ions in cancer cell metastasis〔J〕.Acta Histochemica,2010;112(1):3-25.
9趙遠,王永忠.上皮鈣粘蛋白和β-連環蛋白的研究進展〔J〕.江蘇醫藥,2011;37(13):1581-4.
〔2013-12-07修回〕
(編輯趙慧玲/曹夢園)
