γ-谷維素對脂多糖誘導(dǎo)巨噬細(xì)胞RAW264.7炎癥因子表達(dá)的影響
2015-12-20 07:56:46劉遠(yuǎn)錦田媛媛李新華陳婭婭曾琳娜林親錄羅非君
食品科學(xué) 2015年19期
劉遠(yuǎn)錦,田媛媛,劉 博,李新華,陳婭婭,曾琳娜,楊 濤,林親錄,羅非君,*
(1.中南林業(yè)科技大學(xué)食品科學(xué)與工程學(xué)院,稻谷及副產(chǎn)物深加工國家工程實驗室,湖南長沙 410004;2.中南大學(xué)湘雅醫(yī)院消化內(nèi)科,湖南長沙 410008)
γ-谷維素對脂多糖誘導(dǎo)巨噬細(xì)胞RAW264.7炎癥因子表達(dá)的影響
劉遠(yuǎn)錦1,田媛媛1,劉 博1,李新華2,陳婭婭1,曾琳娜1,楊 濤1,林親錄1,羅非君1,*
(1.中南林業(yè)科技大學(xué)食品科學(xué)與工程學(xué)院,稻谷及副產(chǎn)物深加工國家工程實驗室,湖南長沙 410004;2.中南大學(xué)湘雅醫(yī)院消化內(nèi)科,湖南長沙 410008)
利用細(xì)菌脂多糖(lipopolysaccharide,LPS)刺激巨噬細(xì)胞RAW264.7形成炎癥模型,評價γ-谷維素對巨噬細(xì)胞炎癥因子表達(dá)的影響。在LPS刺激的RAW264.7細(xì)胞培養(yǎng)基中,添加不同濃度的γ-谷維素,分析培養(yǎng)基中炎癥因子白細(xì)胞介素-1β(interleukin-1β,IL-1β)、腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)、白細(xì)胞介素-6 (interleukin-6,IL-6)的含量,以及NO2-/NO3-含量,發(fā)現(xiàn)γ-谷維素能抑制炎癥因子的分泌;利用實時熒光定量聚合酶鏈?zhǔn)椒磻?yīng)(real-time quantitative polymerase chain reaction,qPCR)分析mRNA表達(dá)水平,確定γ-谷維素能抑制炎癥因子的基因表達(dá);采用Western blotting分析進一步確定γ-谷維素能抑制炎癥因子的蛋白表達(dá)。綜上所述,γ-谷維素能明顯抑制炎癥因子IL-1β、TNF-α、IL-6、NO等分泌和表達(dá)。
γ-谷維素;炎癥;炎癥因子;米糠
米糠是稻谷加工的主要副產(chǎn)物之一,谷維素是米糠提取物之一,可能具有抗炎的作用。有研究發(fā)現(xiàn),米糠提取物可以降低Ⅱ型糖尿病大鼠體內(nèi)脂肪組織中的炎癥標(biāo)記物表達(dá)量[1];非絕經(jīng)期的肥胖女性經(jīng)常攝入糙米能降低體內(nèi)炎癥標(biāo)記物的表達(dá)水平[2]。在臨床醫(yī)療中,谷維素是調(diào)節(jié)植物神經(jīng)的常用藥物,但徐百勝等[3]采用谷維素與柳氮磺吡啶聯(lián)用治療潰瘍性結(jié)腸炎,發(fā)現(xiàn)谷維素能增強抗炎療效,提示谷維素具有抗炎的功能。在佛波酯誘導(dǎo)的小鼠皮炎模型中,谷維素及其主要活性成分均能顯著抑制小鼠的炎癥反應(yīng)[4]。本課題組最近的研究也發(fā)現(xiàn)γ-谷維素可抑制葡聚糖硫酸鈉(dextran sulfate sodium,DSS)誘導(dǎo)的小鼠急性和慢性潰瘍性結(jié)腸炎[5]。以上研究結(jié)果揭示谷維素可能具有抗炎作用,但迄今為止,其抗炎的分子機理尚不清楚。
脂多糖(lipopolysaccharide,LPS)是由脂質(zhì)和多糖由共價鍵相連組成的,它是革蘭氏陰性細(xì)菌外膜的主要組成部分,當(dāng)機體被細(xì)菌感染或出現(xiàn)敗血癥時,腸道黏膜屏障破壞,導(dǎo)致LPS進入血液,LPS結(jié)合到CD14/Toll樣受體上,促使免疫細(xì)胞大量分泌腫瘤壞死因子-α (tumor necrosis factor-α,TNF-α)、白細(xì)胞介素-1β (interleukin-1β,IL-1β)、白細(xì)胞介素-6(interleukin-6,IL-6)和NO等炎癥因子,導(dǎo)致機體全身性炎癥反應(yīng)[6-7]。本研究采用LPS誘導(dǎo)巨噬細(xì)胞RAW264.7形成炎癥模型,在分子水平探討γ-谷維素對炎癥因子表達(dá)的影響。
1 材料與方法
1.1 材料與試劑
小鼠單核巨噬細(xì)胞RAW264.7 無錫博慧斯生物醫(yī)藥科技有限公司;γ-谷維素(純度99%) 日本OSHA Haz有限公司;DSS(純度99%)、LPS(純度99%)美國Sigma公司。
高糖DMEM培養(yǎng)基 美國Hyclone公司;胎牛血清(fetal bovine serum,F(xiàn)BS) 浙江天杭生物科技有限公司;反轉(zhuǎn)錄試劑盒、熒光SYBR 北京全式金生物技術(shù)有限公司;鼠IL-1β酶聯(lián)免疫吸附(enzyme-linked immunosorbent assay,ELISA)試劑盒、鼠TNF-α ELISA試劑盒、鼠IL-6 ELISA試劑盒 美國R&D公司;β-actin抗體、IL-1β抗體、TNF-α抗體、IL-6抗體、誘導(dǎo)型一氧化氮合酶(inducible nitric oxide synthase,iNOS)抗體 美國Cell Signaling公司;二抗Anti-rabbit IgG-HRP (SC-2004)、Anti-mouse IgG-HRP(SC-2005) 美國Santa Cruz公司;化學(xué)發(fā)光底物 美國Pierce公司;其他試劑均為國產(chǎn)分析純。
1.2 方法
1.2.1 RAW264.7細(xì)胞培養(yǎng)與分組
RAW246.7巨噬細(xì)胞用含10%胎牛血清、5%青鏈霉素混合液的DMEM高糖培養(yǎng)基培養(yǎng)。隔天傳代,生長至80%~90%時,采用磷酸鹽緩沖液(phosphate buffered saline,PBS)清洗兩次,加入0.25%胰酶在37 ℃消化6~8 min,用移液器打散細(xì)胞后1∶2傳代,37 ℃、5% CO2培養(yǎng)箱中培養(yǎng)。LPS采用純水溶解,γ-谷維素采用質(zhì)量分?jǐn)?shù)1%的四氫呋喃溶液溶解,再用純水稀釋,使四氫呋喃在培基中最終質(zhì)量分?jǐn)?shù)為0.1%。培養(yǎng)的RAW246.7細(xì)胞經(jīng)胰酶消化后,接種于6 孔板,分為4 組進行處理:對照組,單純LPS處理組,高、低濃度γ-谷維素組。高、低濃度γ-谷維素組中分別加入12.5、25 μmol/L γ-谷維素預(yù)處理30 min后加入終質(zhì)量濃度為1 μg/mL的LPS,單純LPS處理組僅加入LPS溶液和四氫呋喃,對照組加入蒸餾水和四氫呋喃,使4 組細(xì)胞培養(yǎng)基中四氫呋喃最終質(zhì)量分?jǐn)?shù)均為0.1%,細(xì)胞培養(yǎng)24 h后,分別收集培養(yǎng)基和細(xì)胞做進一步分析。
1.2.2 γ-谷維素對RAW264.7細(xì)胞的毒性
采用四甲基偶氮唑藍(lán)(methyl thiazolyl tetrazolium,MTT)比色法測定γ-谷維素和LPS對細(xì)胞是否有毒性作用[8-9]。取對數(shù)生長期的RAW246.7細(xì)胞,消化后吹打均勻,用培養(yǎng)基稀釋至細(xì)胞密度為7×104個/mL,以每孔100 μL接種于96 孔板中,置于37 ℃、5% CO2培養(yǎng)箱中培養(yǎng)。待細(xì)胞貼壁后,每孔加入2 μL的γ-谷維素溶液,使得四氫呋喃最終質(zhì)量分?jǐn)?shù)為0.1%,γ-谷維素的終濃度分別為0、3.125、6.25、12.5、25、50、100 μmol/L。每組設(shè)置5 個重復(fù)孔。輕輕振蕩,放入培養(yǎng)箱培養(yǎng)20 h后,每孔加入20 μL MTS(3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium, inner salt)溶液。再次培養(yǎng)4 h后,采用酶標(biāo)儀在490 nm波長處測定每個孔的光密度值。
1.2.3 ELISA法檢測炎癥因子TNF-α、IL-6和IL-1β的含量
TNF-α、IL-6和IL-1β是重要的炎癥因子。按照1.2.1節(jié)方法將RAW246.7細(xì)胞分為4 組并做相應(yīng)處理,每組3 個重復(fù),培養(yǎng)24 h后取細(xì)胞培養(yǎng)液,室溫條件下1 000×g離心10 min,收集細(xì)胞培養(yǎng)液。嚴(yán)格按照試劑盒說明書步驟操作。繪制出標(biāo)準(zhǔn)曲線,測定細(xì)胞培養(yǎng)基中TNF-α、IL-6和IL-1β的含量。
1.2.4 NO2-/NO3-含量檢測
巨噬細(xì)胞受到LPS信號的刺激會釋放NO,但NO非常不穩(wěn)定,會形成亞硝酸鹽(NO2-)和硝酸鹽(NO3-)。取對數(shù)生長期的RAW246.7細(xì)胞消化,稀釋成5×105個/mL細(xì)胞懸浮液,接種于24 孔板中,每孔1 mL。24 h后,棄掉舊培養(yǎng)基,加入新培養(yǎng)基,每孔加入LPS使其終質(zhì)量濃度為1 μg/mL,加入γ-谷維素使其終濃度分別為12.5、25 μmol/L,四氫呋喃終質(zhì)量分?jǐn)?shù)為0.1%。培養(yǎng)24 h后,取細(xì)胞培養(yǎng)液按照NO試劑盒說明書方法進行操作,測定NO含量(以NO2-/NO3-表示,為NO2-和NO3-的總量)。按照下式計算NO2-/NO3-含量。

式中:20 μmol/L為標(biāo)準(zhǔn)品濃度;n為樣品測定前的稀釋倍數(shù);空白組為未加細(xì)胞只有培養(yǎng)基的處理。
1.2.5 實時熒光定量聚合酶鏈?zhǔn)椒磻?yīng)(real-time quantitative polymerase chain reaction,qPCR)分析基因mRNA表達(dá)水平
6 孔板培養(yǎng)RAW246.7細(xì)胞,用不同濃度γ-谷維素處理,LPS刺激12 h后,用PBS清洗2 次,以Trizol提取細(xì)胞總RNA,0.8%瓊脂糖膠電泳檢測RNA的完整性并測定其濃度,用反轉(zhuǎn)錄試劑盒合成cDNA后進行PCR擴增反應(yīng)。iNOS:上游引物5’-CAG CTG GGC TGT ACA AAC CTT-3’,下游引物5’-CAT TGG AAG TGA AGC GTT TCG-3’;TNF-α:上游引物5’-CAA AAT TCG AGT GAC AAG CCT G-3’,下游引物5’-GAG ATC CAT GCC GTT GGC-3’;IL-1β:上游引物5’-GAG CAC CTT CTT TTC CTT CAT CTT-3’,下游引物5’-TCA CAC ACC AGC AGG TTA TCA TC-3’;IL-6:上游引物5’-ATG GAT GCT ACC AAA CTG GAT-3’,下游引物5’-TGA AGG ACT CTG GCT TTG TCT-3’;β-actin:上游引物5’-CCA TAA ACG ATG CCG GA-3’,下游引物5’-CAC CAC CCA TAG AAT CAA GA-3’。qPCR反應(yīng)條件:94 ℃,3 min;94 ℃,30 s;60 ℃,40 s;72 ℃,1 min;40 個循環(huán)。
1.2.6 Western blotting分析
將RAW264.7細(xì)胞用γ-谷維素和LPS處理24 h后,PBS清洗2 次,用含苯甲基磺酰氟(phenylmethylsulfonyl fluoride,PMSF)的RIPA裂解液(50 mmol/L Tris-HCl (pH 7.4)、150 mmol/L NaCl、1% NP-40、0.1%十二烷基硫酸鈉(sodium dodecyl sulfate,SDS))裂解后,5 000×g離心10 min,取上清液。取10 μL蛋白質(zhì)裂解液,直接采用Nanodrop儀測定蛋白質(zhì)濃度,取20 μg的蛋白質(zhì)樣品進行SDS-聚丙烯酰胺凝膠電泳(SDS-polyacrylamide gelelectrophoresis,SDS-PAGE),再轉(zhuǎn)移到硝酸纖維素膜上。將硝酸纖維膜在含有5%脫脂奶粉的PBST(PBS和Tween-20混合)中封閉1 h。一抗孵育過夜(稀釋比例為1∶1 000),孵育后用PBST清洗4 次,室溫條件下二抗孵育2 h(稀釋比例為1∶1 000)。繼續(xù)用PBST清洗4 次。加入化學(xué)發(fā)光底物,X光片曝光、顯影、定影顯示陽性條帶,樣本結(jié)果用看家蛋白β-actin條帶作為內(nèi)參校正。
1.3 統(tǒng)計學(xué)分析
實驗數(shù)據(jù)用SPSS 17.0統(tǒng)計軟件進行處理,數(shù)據(jù)結(jié)果以±s表示,組間比較采用方差分析和t檢驗,P<0.05表示差異顯著,P<0.01表示差異極顯著。
2 結(jié)果與分析
2.1 γ-谷維素對RAW246.7細(xì)胞的毒性作用分析
如圖1所示,不同濃度γ-谷維素作用于RAW246.7細(xì)胞24 h后,對照組和實驗組細(xì)胞存活率分別為:(100±2.03)%、(103.99±4.98)%、(107.65±9.05)%、(103.58±3.90)%、(111.24±12.15)%、(106.38±6.37)%、(97.96±7.20)%。與對照組相比,不同濃度γ-谷維素處理后,細(xì)胞活力均無明顯改變(P>0.05),提示0~100 μmol/L的γ-谷維素對RAW264.7細(xì)胞無明顯的毒性作用,該模型可用于后續(xù)實驗。后續(xù)研究將選用1 μg/mL LPS(經(jīng)典處理劑量)刺激RAW246.7細(xì)胞建立細(xì)胞炎癥模型,采用12.5、25 μmol/L γ-谷維素來評估其抗炎功效及分子機理。

圖1 1 γ--谷維素對RAW264.7細(xì)胞的毒性作用Fig.1 Effect of γ-oryzanol on the viability of RAW264.7 cells
2.2 γ-谷維素對LPS誘導(dǎo)RAW264.7細(xì)胞炎癥因子分泌量的影響
LPS和γ-谷維素處理RAW246.7細(xì)胞24 h后,收集細(xì)胞培養(yǎng)液,離心取上清液,測定細(xì)胞炎癥因子的分泌量,結(jié)果見圖2。
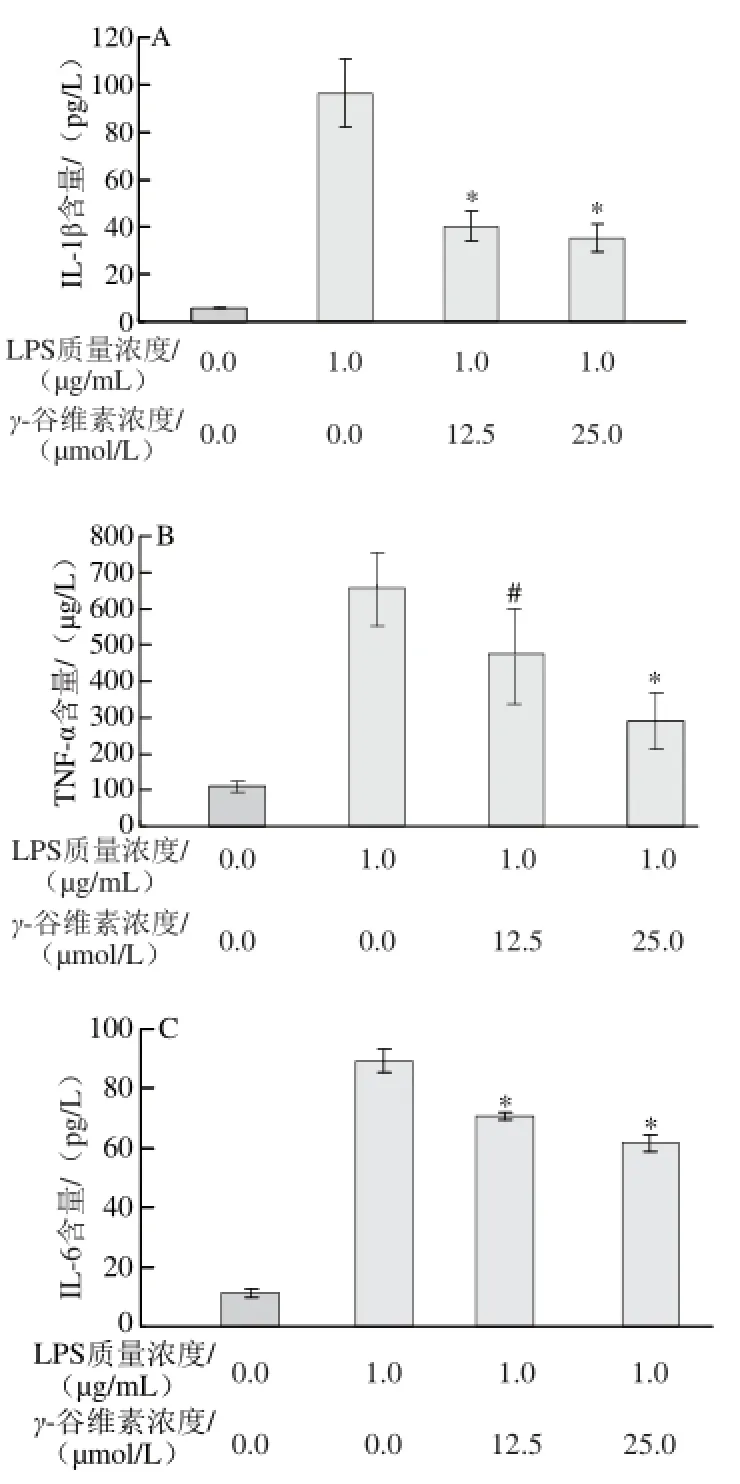
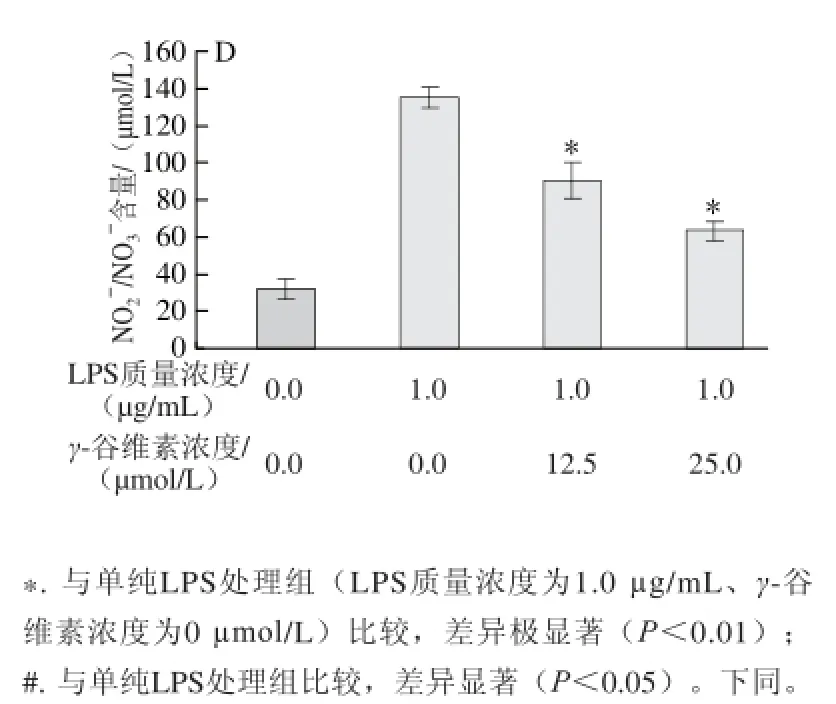
圖2 2 γ-谷維素對LPS誘導(dǎo)RAW264.7細(xì)胞炎癥因子分泌量的影響Fig.2 Effect of γ-oryzanol on the production of infl ammatory factors in LPS-stimulated RAW264.7 cells
如圖2所示,ELISA分析結(jié)果顯示,加入LPS刺激后,培養(yǎng)基中IL-1β含量從(15.31±3.12)pg/L增加至(96.32±14.53)pg/L,12.5、25 μmol/L γ-谷維素處理后,培養(yǎng)基中IL-1β含量分別為(40.15±6.56)、(35.31±5.87) pg/L,相對于單純LPS處理組,γ-谷維素處理可極顯著抑制RAW246.7細(xì)胞分泌IL-1β (P<0.01)(圖2A)。γ-谷維素也能抑制TNF-α的分泌,12.5、25 μmol/L γ-谷維素處理后,培養(yǎng)基中TNF-α含量從(656.32±98.37) μg/L分別降至(475.86±130.78)、(290.62±75.46) μg/L,其中25 μmol/L γ-谷維素處理組與單純LPS處理組相比具有極顯著差異(P<0.01)(圖2B)。LPS刺激RAW264.7細(xì)胞炎癥模型中,培養(yǎng)基中IL-6含量從(11.13±0.19) pg/L上升至(89.28±3.75) pg/L,12.5、25 μmol/L γ-谷維素處理后,培養(yǎng)基中IL-6含量分別降低至(70.84±1.24)、(61.54±2.78)pg/L(P<0.01)(圖2C)。在炎癥反應(yīng)中,巨噬細(xì)胞釋放的NO非常不穩(wěn)定,在培養(yǎng)基中形成會形成亞硝酸鹽(NO2-)和硝酸鹽(NO3-),NO測定試劑盒將硝酸鹽還原為亞硝酸鹽,確定培基中NO的釋放量,結(jié)果發(fā)現(xiàn)γ-谷維素亦能抑制NO的釋放,12.5、25 μmol/L γ-谷維素處理后,培養(yǎng)基中NO2-/NO3-濃度從(135.47±4.85)μmol/L分別降至(90.48±9.48)、(64.06±4.53)μmol/L(P<0.01)(圖2D)。
2.3 γ-谷維素對細(xì)胞炎癥因子和iNOS mRNA表達(dá)水平的影響
如圖3所示,單純LPS處理RAW246.7細(xì)胞12 h后,IL-1β mRNA相對表達(dá)量從1.00±0.31增加至15.31±3.12,12.5、25 μmol/L γ-谷維素處理后,RAW246.7細(xì)胞的IL-1β mRNA相對表達(dá)量分別為10.02±1.82和5.12±1.09,相對于單純LPS處理組,γ-谷維素處理可極顯著抑制IL-1β mRNA轉(zhuǎn)錄(P<0.01)。γ-谷維素也能抑制TNF-α mRNA轉(zhuǎn)錄,12.5、25 μmol/L γ-谷維素處理后,RAW246.7細(xì)胞的TNF-α mRNA相對表達(dá)量從4.08±0.41(單純LPS處理組)分別降至2.61±0.54 和1.81±0.31,與單純LPS處理組相比具有極顯著差異(P<0.01)。LPS刺激RAW264.7細(xì)胞炎癥模型中,IL-6 mRNA相對表達(dá)量為4.57±1.38,12.5、25 μmol/L γ-谷維素處理后,RAW246.7細(xì)胞的IL-6 mRNA相對表達(dá)量分別降至2.25±0.47和1.96±0.45。在炎癥反應(yīng)中,iNOS高表達(dá)是促使炎癥因子NO釋放的重要條件之一,本研究發(fā)現(xiàn),γ-谷維素亦能抑制iNOS基因的轉(zhuǎn)錄,12.5 μmol/L γ-谷維素處理后,RAW246.7細(xì)胞的iNOS mRNA相對表達(dá)量從8.07±1.54下降到6.01±1.82 (P<0.05);25 μmol/L γ-谷維素處理后,iNOS mRNA相對表達(dá)量則降至3.17±1.25(P<0.01)。
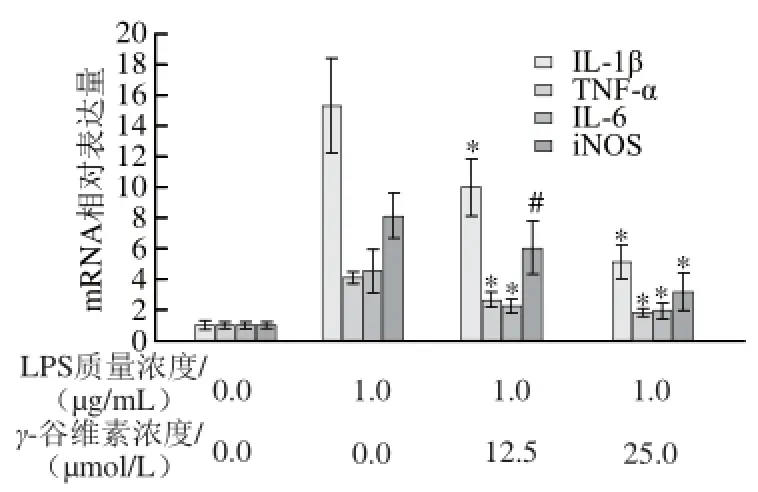
圖 33 γ-谷維素對LPS刺激RAW264.7細(xì)胞炎癥因子和iiNNOOSS mRNA表達(dá)水平的影響Fig.3 Effect of γ-oryzanol on mRNA expression of infl ammatory factors and iNOS in LPS-stimulated RAW264.7 cells
2.4 γ-谷維素對炎癥因子和iNOS蛋白表達(dá)水平的影響


圖4 Western blotting分析γ-谷維素細(xì)胞炎癥因子和iNOS蛋白表達(dá)的影響Fig.4 Effect of γ-oryzanol on protein expression of infl ammatory factors and iNOS in LPS-stimulated RAW264.7 cells
如圖4所示,Western blotting分析結(jié)果表明,LPS處理RAW246.7細(xì)胞24 h后,IL-1β、TNF-α、IL-6和iNOS蛋白表達(dá)水平明顯上升,而在對照組(0、0)中其表達(dá)水平非常低。與單純LPS處理組相比,γ-谷維素處理后,RAW246.7細(xì)胞中各種炎癥因子水平均有所下降。
3 討 論
炎癥是機體針對外界病毒、化學(xué)物質(zhì)入侵而發(fā)起的維持機體穩(wěn)態(tài)的自發(fā)反應(yīng)。大量研究表明,炎癥因子的異常激活與眾多疾病有密切的關(guān)系[10-11]。如有研究表明慢性炎癥與癌變密切相關(guān),參與機體的癌變演進[12-13]。Sun 等[14]研究發(fā)現(xiàn)炎癥因子的高表達(dá)與癌細(xì)胞的藥物抗性密切相關(guān),IL-6的高表達(dá)可以提高乳腺癌患者對他莫昔芬(一種抗雌激素)的耐藥性。潰瘍性結(jié)腸炎是一種常見的慢性炎癥,其病因復(fù)雜,常反復(fù)發(fā)作并逐漸加重,在潰瘍性結(jié)腸炎組織中存在大量炎癥因子的表達(dá)。因此,對炎癥因子的調(diào)控具有非常重要的意義,特別是利用食品中的營養(yǎng)成分來控制炎癥因子的表達(dá)具有副作用少等突出優(yōu)點。
γ-谷維素主要存在于米糠油及其油腳中,米糠層中γ-谷維素的含量為0.3%~0.5%。在加溫壓榨米糠時,γ-谷維素溶于油中,一般毛糠油中γ-谷維素的含量約為2%~3%。隨著研究的深入,目現(xiàn)已發(fā)現(xiàn)γ-谷維素具有多種藥理活性,如抗氧化、降脂、調(diào)節(jié)機體植物神經(jīng)紊亂等[15-18]。本研究以RAW246.7巨噬細(xì)胞為材料,明確了γ-谷維素對LPS刺激細(xì)胞所產(chǎn)生的炎癥反應(yīng)具有明顯的抑制作用,為γ-谷維素抗炎功效研究提供了新的實驗依據(jù)。當(dāng)前已有醫(yī)生在臨床上將γ-谷維素用于患者的抗炎輔助治療中[3,19],但還亟需提供這方面的實驗依據(jù)支持。
本研究發(fā)現(xiàn)γ-谷維素能抑制主要炎癥因子IL-1β、TNF-α、IL-6和NO等表達(dá),其抗炎作用體現(xiàn)出了多靶點性。也有研究表明,炎癥因子之間可能會存在一些自我加強的作用,如在正常情況下,細(xì)胞產(chǎn)生的IL-6基礎(chǔ)分泌量很少,在有外界刺激時,如在感染、創(chuàng)傷、TNF-α影響下,可觸發(fā)細(xì)胞內(nèi)IL-6釋放或合成[20-21]。NO作為一種重要的細(xì)胞間信息交流調(diào)節(jié)因子,其過量釋放與炎癥的產(chǎn)生有密切關(guān)系,其中iNOS是合成NO重要的限速酶,NO釋放往往與iNOS酶高表達(dá)有關(guān)[20]。關(guān)麗華等[21]研究發(fā)現(xiàn),在潰瘍性結(jié)腸炎模型中,iNOS表達(dá)升高,NO釋放量增加。通過對iNOS基因轉(zhuǎn)錄調(diào)控的研究發(fā)現(xiàn)其基因受核轉(zhuǎn)錄因子-κB(nuclear factor-κB,NF-κB)的調(diào)控[22]。Sakai 等[23]發(fā)現(xiàn)在牛動脈內(nèi)皮細(xì)胞中γ-谷維素能抑制NF-κB的活化,減少細(xì)胞黏附分子VCAM-1和ICAM-1的表達(dá)。同時,已有研究證實IL-1β、TNF-α、IL-6和iNOS這4 個基因的啟動子中均存在NF-κB的結(jié)合位點,其表達(dá)受NF-κB活性的調(diào)控[24-27],提示γ-谷維素也可能通過抑制NF-κB的活化起作用,但需要進一步證實γ-谷維素是否能抑制NF-κB的核轉(zhuǎn)運及其與順式調(diào)控元件的結(jié)合能力。雖然本實驗證實了γ-谷維素能抑制巨噬細(xì)胞炎癥因子的表達(dá),但其調(diào)控炎癥因子表達(dá)的信號轉(zhuǎn)導(dǎo)通路尚需進一步探索。
本研究利用MTT實驗發(fā)現(xiàn)在0~100 μmol/L濃度范圍內(nèi),γ-谷維素對巨噬細(xì)胞均無明顯毒性作用,這將有利于今后γ-谷維素相關(guān)功能性食品和保健品的研發(fā)。
總之,本研究在評估γ-谷維素抗炎功效的基礎(chǔ)上,對γ-谷維素抗炎的分子機理進行了初步的探索,明確了γ-谷維素能明顯抑制炎癥因子IL-1β、TNF-α、IL-6和NO的分泌和表達(dá)。對其分子機理的深入研究,將促進含γ-谷維素的功能食品及保健品的開發(fā),同時,也能為人們的膳食方案提供科學(xué)的理論依據(jù)。
[1] CANDIRACCI M, LUISA JUSTO M, CASTANO A, et al. Rice bran enzymatic extract supplemented diets modulate adipose tissue infl ammation markers in Zucker rats[J]. Nutrition, 2014, 30(4): 466-472.
[2] KAZEMADEH M, SAFAVI S M, NEMATOLLAHI S, et al. Effect of brown rice consumption on infl ammatory marker and cardiovascular risk factors among overweight and obese non-menopausal female adults[J]. International Journal of Preventive Medicine, 2014, 5(4): 478-488.
[3] 徐百勝. γ-谷維素、柳氮磺吡啶聯(lián)用治療潰瘍性結(jié)腸炎的療效觀察[J].臨床和實驗醫(yī)學(xué)雜志, 2011, 2(1): 127-128.
[4] YASUKAWA K, AKIHISA T, KIMURA Y, et al. Inhibitory effect of cycloartenol ferulate, a component of rice bran, on tumor promotion in two-stage carcinogenesis in mouse skin[J]. Biological & Pharmaceutical Bulletin, 1998, 21(10): 1072-1076.
[5] 李輝. 米糠γ-谷維素抗急性腸炎的功能評價及初步機理研究[D]. 長沙: 中南林業(yè)科技大學(xué), 2014: 18-30.
[6] LI Meng, ZHANG Lei, CAI Runlan, et al. Lipid-soluble extracts from Salvia miltiorrhiza inhibit production of LPS-induced inflammatory mediators via NF-κB modulation in RAW 264.7 cells and perform antiinflammatory effects in vivo[J]. Phytotherapy Research, 2012, 26(8): 1195-1204.
[7] RYU S, SHIN J S, CHO Y W, et al. Fimasartan, anti-hypertension drug, suppressed inducible nitric oxide synthase expressions via nuclear factor-kappa B and activator protein-1 inactivation[J]. Biological & Pharmaceutical Bulletin, 2013, 36(3): 467-474.
[8] CHOU S T, PENG H Y, HSU J C, et al. Achillea millefolium L. essential oil inhibits LPS-induced oxidative stress and nitric oxideproduction in RAW 264.7 macrophages[J]. International Journal of Molecular Science, 2013, 14(7): 12978-12993.
[9] RAETZ C R H, WHITFIELD C. Lipopolysaccharide endotoxins[J]. Annual Review of Biochemistry, 2002, 71: 635-700.
[10] LI Xiaojun, JIANG Jiaye, SHI Songshan, et al. A RG-II type polysaccharide purified from Aconitum coreanum alleviates lipopolysaccharide-induced inflammation by inhibiting the NF-κB signal pathway[J]. PLoS ONE, 2014, 9(6): e99697-e99705. doi: 10.1371/journal.pone.0099697.
[11] LI Weidong, CHEN Cihui, SAUD S M, et al. Fei-Liu-Ping ointment inhibits lung cancer growth and invasion by suppressing tumor inflammatory microenvironment[J]. BMC Complementary and Alternative Medicine, 2014, 14: 153-159.
[12] LANDSKRON G, de la FUENTE M, THUWAJIT P, et al. Chronic Inflammation and cytokines in the tumor microenvironment[J/OL]. Journal of Immunology Research, 2014. http://dx.doi.org/10.1155/2014/149185.
[13] BALKWILL F,MANTOVANI A. Cancer and inflammation: implications for pharmacology and therapeutics[J]. Clinical Pharmacology & Therapeutics, 2010, 87(4): 401-406.
[14] SUN X, MAO Y, WANG J, et al. IL-6 secreted by cancer-associated fibroblasts induces tamoxifen resistance in luminal breast cancer[J]. Oncogene, 2014, 33(35): 4450-4457.
[15] 田媛媛, 林親錄, 羅非君. 米糠γ-谷維素生理功能研究新進展[J]. 糧食與油脂, 2013, 26(9): 43-46.
[16] 龔院生, 姚惠源. γ-谷維醇清除自由基作用的研究[J]. 中國糧油學(xué)報, 2003, 18(1): 10-12.
[17] ISLAM M S, NAGASAKA R, OHARA K, et al. Biological abilities of rice bran-derived antioxidant phytochemicals for medical therapy[J]. Current Topics in Medicinal Chemistry, 2011, 11(14): 1847-1853.
[18] GOUFO P, TRINDADE H. Rice antioxidants: phenolic acids, fl avonoids, anthocyanins, proanthocyanidins, tocopherols, tocotrienols, γ-oryzanol, and phytic acid[J]. Food Science Nutrition, 2014, 2(2): 75-104.
[19] 雷培森, 趙紅. 甲硝唑、谷維素、維生素B1聯(lián)合治療潰瘍性結(jié)腸炎[J].綜合臨床醫(yī)學(xué), 1995, 4(2): 215-216.
[20] 李昕, 劉佳佳. IL-6對中性粒細(xì)胞在炎癥中作用的影響[J]. 國外醫(yī)學(xué)免疫學(xué)分冊, 2005, 28(5): 277-280.
[21] 關(guān)麗華, 龔玉芳, 張弘, 等. 結(jié)腸康對惡唑酮誘導(dǎo)小鼠潰瘍性結(jié)腸炎MPO、NO、iNOS的影響[J]. 中成藥, 2013, 35(4): 669-673.
[22] ATREYA I, ATREYA R, NEURATH M F. NF-κB in infl ammatory bowel disease[J]. Journal of Internal Medicine, 2008, 263(6): 591-596.
[23] SAKAI S, MURATA T, TSUBOSAKA Y, et al. γ-Oryzanol reduces adhesion molecule expression in vascular endothelial cells via suppression of nuclear factor-κB activation[J]. Journal of Agricultural and Food Chemistry, 2012, 60(13): 3367-3372.
[24] COGSWELL J P, GODLEVSKI M M, WISELY G B, et al. NF-kappa B regulates IL-1 beta transcription through a consensus NF-kappa B binding site and a nonconsensus CRE-like site[J]. Journal of Immunology, 1994, 153(2): 712-723.
[25] MATSUSAKA T, FUJIKAWA K, NISHIO Y, et al. Transcription factors NF-IL6 and NF-kappa B synergistically activate transcription of the inflammatory cytokines, interleukin 6 and interleukin 8[J]. Proceedings of the National Academy of Sciences, 1993, 90(21): 10193-10197.
[26] TREDE N S, TSYTSYKOVA A V, CHATILA T, et al. Transcriptional activation of the human TNF-alpha promoter by superantigen in human monocytic cells: role of NF-kappa B[J]. Journal of Immunology, 1995, 155(2): 902-908.
[27] XIE Q W, KASHIWABARA Y, NATHAN C. Role of transcription factor NF-kappa B/Rel in induction of nitric oxide synthase[J]. Journal of Biological Chemistry, 1994, 269(7): 4705-4708.
Effect of γ-Oryzanol on the Expression of Lipopolysaccharide-Induced Infl ammatory Factors in Macrophages
LIU Yuanjin1, TIAN Yuanyuan1, LIU Bo1, LI Xinhua2, CHEN Yaya1, ZENG Linna1, YANG Tao1, LIN Qinlu1, LUO Feijun1,*
(1. National Engineering Laboratory for Deep Processing of Rice and Byproducts, College of Food Science and Engineering, Central South University of Forestry and Technology, Changsha 410004, China; 2. Department of Gastroenterology, Xiangya Hospital, Central South University, Changsha 410008, China)
In this study, we used macrophage models of lipopolysaccharide (LPS)-stimulated infl ammation to evaluate the effect of γ-oryzanol on the expression of infl ammatory factors. After exposing RAW264.7 cells to different concentrations of γ-oryzanol and LPS, we analyzed the contents of IL-1β, TNF-α, IL-6 and NO2-/NO3-(NO) in the macrophage culturemedium and found that γ-oryzanol could inhibit the secretion of inflammatory factors. Real-time quantitative PCR was employed to analyzed mRNA expression levels of IL-1β, TNF-α, IL-6 and iNOS, and it was found that infl ammatory gene transcriptions were inhibited by γ-oryzanol. Western blotting analysis further confi rmed that γ-oryzanol could decrease the expression of IL-1β, TNF-α, IL-6 and iNOS protein. Our data showed that γ-oryzanol can signifi cantly inhibit the expression and secretion of infl ammatory factors such as IL-1β, TNF-α, IL-6 and NO.
γ-oryzanol; infl ammation; infl ammatory factor; rice bran
R284.1
A
1002-6630(2015)19-0238-06
10.7506/spkx1002-6630-201519043
2014-11-13
湖南省教育廳科學(xué)研究重點項目(13A124);湖南省研究生科研創(chuàng)新項目(122-0035);
中南林業(yè)科技大學(xué)研究生科研創(chuàng)新項目(CX201313357;CX2013B358;CX2013B14)
劉遠(yuǎn)錦(1990-),女,碩士研究生,研究方向為食品分子營養(yǎng)學(xué)。E-mail:312334609@qq.com
*通信作者:羅非君(1968-),男,研究員,博士,研究方向為食品分子營養(yǎng)學(xué)。E-mail:luofeijun@hotmail.com
