鈣調(diào)神經(jīng)磷酸酶-T細胞核因子信號通路在應力誘導成肌細胞凋亡中的作用
2015-12-16 07:24:52丁弦夏晨蕾賀苗孫文娜王芳姜文心張彩霞王爽玉張強姚如永袁曉
華西口腔醫(yī)學雜志 2015年5期
關(guān)鍵詞:檢測
丁弦 夏晨蕾 賀苗 孫文娜 王芳 姜文心張彩霞 王爽玉 張強 姚如永 袁曉
1.青島大學醫(yī)學院附屬青島市立醫(yī)院口腔醫(yī)學中心,青島 266075;2.青島大學醫(yī)學院附屬醫(yī)院中心實驗室,青島 266003
頜面部肌肉組織在錯 畸形的發(fā)生、發(fā)展、矯治和療效維持中發(fā)揮重要作用。臨床上,正畸醫(yī)生通過調(diào)控矯形力來牽張咀嚼肌,影響牙齒、頜骨及顳下頜關(guān)節(jié)的改建,進而使軟硬組織發(fā)生適應性變化,重建新的功能平衡,達到預防錯 畸形及調(diào)控生長的目的[1-2]。在此過程中,成肌細胞作為適應性改建的主要體現(xiàn)者,明確其力學信號轉(zhuǎn)導機制對于闡明功能矯形時面頜部肌肉組織的適應性改建機制有重要意義[3]。本課題前期研究已證實,周期性張應力可誘導成肌細胞凋亡,在凋亡過程中Ca2+作為細胞內(nèi)信號轉(zhuǎn)導分子之一,具有重要的作用[4]。研究發(fā)現(xiàn),鈣調(diào)神經(jīng)磷酸酶(calcineurin,CaN)作為現(xiàn)階段發(fā)現(xiàn)的唯一受Ca2+/鈣調(diào)素(calmodulin,CaM)調(diào)節(jié)的磷蛋白磷酸酶,其介導的信號通路與細胞凋亡關(guān)系密切,在細胞信號傳遞過程中直接受Ca2+的調(diào)節(jié),通過其重要底物活化T細胞核因子(nuclear factor of activated T cells,NFAT)去磷酸化和核內(nèi)轉(zhuǎn)位發(fā)揮功能調(diào)節(jié)作用。CaN-NFAT信號通路是內(nèi)質(zhì)網(wǎng)應激的一條重要通路,本研究擬在周期性張應力作用下明確CaN-NFAT信號通路在成肌細胞凋亡中的作用,進一步探明應力作用下成肌細胞適應性改建的分子機制,為功能矯形提供理論支持。
1 材料和方法
1.1 材料和儀器
大鼠L6成肌細胞株,購自中國科學院上海生科院細胞資源中心。胎牛血清、胰蛋白酶(Hyclone公司,美國),Annexin V-FITC/PI細胞凋亡試劑盒(北京嘉美諾斯生物科技有限公司),RT-PCR試劑盒、Trizol提取試劑盒(Takara公司,日本),半胱天冬氨酸蛋白酶9兔抗大鼠抗體(Abcam公司,英國),BioFlex彈性細胞培養(yǎng)板(Flexcell公司,美國)。
1.2 細胞培養(yǎng)
將成肌細胞按1×107·L-1的濃度接種于培養(yǎng)瓶中,加入含體積分數(shù)10%的胎牛血清、雙抗,配成最終濃度為100 UI·mL-1的高糖DMEM培養(yǎng)液,在CO2孵箱中37 ℃、5%CO2、95%空氣、飽和濕度下培養(yǎng)。48 h換液一次,并通過倒置相差顯微鏡觀察成肌細胞生長狀況及變化。待細胞生長至培養(yǎng)瓶底80%以上時進行傳代。
1.3 成肌細胞體外培養(yǎng)—力學刺激模型的構(gòu)建
選擇對數(shù)生長期傳至第3代左右的成肌細胞胰酶消化后,進行細胞計數(shù),將接種密度調(diào)整到3×105·mL-1后接種至BioFlex培養(yǎng)板,放置于37 ℃、5%CO2培養(yǎng)箱內(nèi)培養(yǎng)1~2 d,當細胞融合達80%以上時,用低血清濃度(含體積分數(shù)為3%胎牛血清的DMEM)的培養(yǎng)液繼續(xù)培養(yǎng)24 h。
將成肌細胞分為加力組和對照組,利用多通道應力加載系統(tǒng)對加力組細胞分別施加1、2、6、12、24 h頻率為10 cycles·min-1、15%細胞形變的張應力,每一循環(huán)包括3 s拉伸和3 s松弛;對照組放在同一培養(yǎng)箱內(nèi)不加力(即加力0 h)。應力結(jié)束后,于倒置相差顯微鏡下觀察成肌細胞生長狀況以及形態(tài)變化。
1.4 Hoechst 33258染色法檢測成肌細胞凋亡
2組細胞加入固定液固定10 min,沖洗完后向培養(yǎng)板中加入Hoechst 33258染色液,染色5 min,于熒光顯微鏡(激發(fā)波長350 nm,發(fā)射波長460 nm)下觀察細胞凋亡情況。將CaN的特異性抑制劑環(huán)孢素(cyclosporin A,CsA)加入24 h(加抑制劑組)后,觀察細胞凋亡情況。每組重復3次。
1.5 流式細胞術(shù)檢測成肌細胞凋亡
2組細胞棄舊培養(yǎng)液,PBS洗兩遍,用不含EDTA的胰酶消化后收集細胞至離心管中,再按照Annexin V-FITC/PI試劑盒說明書進行標記,在室溫下避光染色15 min后用流式細胞儀檢測細胞凋亡。在加力24 h的成肌細胞中加入CsA(加抑制劑組)后,再次檢測細胞凋亡。每組重復3次。
1.6 實時聚合酶鏈式反應(real-time polymerase chain reaction,real-time PCR)檢測CaN和NFAT mRNA的表達情況
收集2組細胞,按照Gibco BRL公司的說明書提取細胞總RNA,分光光度計測量RNA的濃度和純度,利用反轉(zhuǎn)錄試劑盒將RNA反轉(zhuǎn)錄為cDNA,將得到的反應液加入到real-time PCR的反應體系中,再對上海生工生物技術(shù)有限公司合成的引物進行PCR擴增反應,檢測CaN和NFAT mRNA的表達情況。2組細胞中加入CsA后,再次檢測CaN和NFAT mRNA的表達情況。各因子的引物見表1。

表1 real-time PCR各因子引物Tab 1 Primers of genes used in real-time PCR
1.7 蛋白質(zhì)印跡法(western blot)檢測NFAT3的蛋白含量
收集2組細胞,使用裂解液提取總蛋白,采用BCA蛋白濃度測定試劑盒測定蛋白濃度,各組取等量蛋白上樣,電泳分離后轉(zhuǎn)移至PVDF膜上,封閉后將一抗與一抗稀釋液按比例混勻后與PVDF膜4 ℃過夜,PBST沖洗3遍;再將配好的二抗稀釋液和膜在搖床上搖動孵育1 h,用PBST沖洗3遍。加顯影液后放入計算機顯影系統(tǒng)內(nèi),將蛋白的表達量拍照,并保存記錄。2組細胞中加入CsA后,再重復上述步驟。對圖像進行分析并計算灰度值。
1.8 統(tǒng)計學分析
采用SPSS 17.0軟件進行分析,對數(shù)據(jù)進行方差分析和配對t檢驗,P<0.05為差異具有統(tǒng)計學意義。
2 結(jié)果
2.1 成肌細胞體外培養(yǎng)力學刺激模型的構(gòu)建
倒置相差顯微鏡下觀察可見,對照組成肌細胞貼壁良好,但排列方向雜亂無章;加力組細胞貼壁良好且生長狀態(tài)佳,無明顯變性、脫落;隨著加力時間的延長,成肌細胞逐漸順應力場方向排列,且呈時間依賴性(圖1)。成肌細胞體外培養(yǎng)—力學刺激模型構(gòu)建成功。

圖1 各組成肌細胞的形態(tài) 倒置相差顯微鏡 × 400Fig 1 Myoblasts morphology of every group inverted phase contrast microscope × 400
2.2 Hoechst 33258染色檢測成肌細胞凋亡
對照組成肌細胞的胞核清晰且呈卵圓形,染色質(zhì)分布均勻,發(fā)出淡藍色熒光;加力組出現(xiàn)凋亡細胞,胞核發(fā)生固縮,發(fā)出致密濃染的亮藍色熒光,個別顏色發(fā)白,隨加力時間的延長,凋亡現(xiàn)象越發(fā)明顯;加抑制劑組凋亡細胞明顯比加力24 h組減少,說明CsA能有效抑制細胞凋亡(圖2)。

圖2 各組成肌細胞的形態(tài) Hoechst 33258染色 × 400Fig 2 Myoblasts morphology of every group Hoechst 33258 staining × 400
2.3 流式細胞術(shù)檢測成肌細胞凋亡
流式細胞術(shù)檢測發(fā)現(xiàn),對照組細胞的凋亡率為(4.47±0.93)%,加力1、2、6、12、24 h組的細胞凋亡率分別為(5.83±0.26)%、(6.56±0.34)%、(6.93±0.52)%、(7.75±0.15)%、(8.17±0.28)%,加抑制劑組的細胞凋亡率為(2.17±0.13)%。統(tǒng)計分析表明,加力組與對照組之間的細胞凋亡率有統(tǒng)計學差異(P<0.05)。隨加力時間的延長,細胞凋亡率及死細胞比率逐漸升高,24 h達到高峰。與單純加力24 h組相比,加抑制劑組的細胞凋亡率和死細胞比率均下降,而活細胞比率有所上升(圖3)。
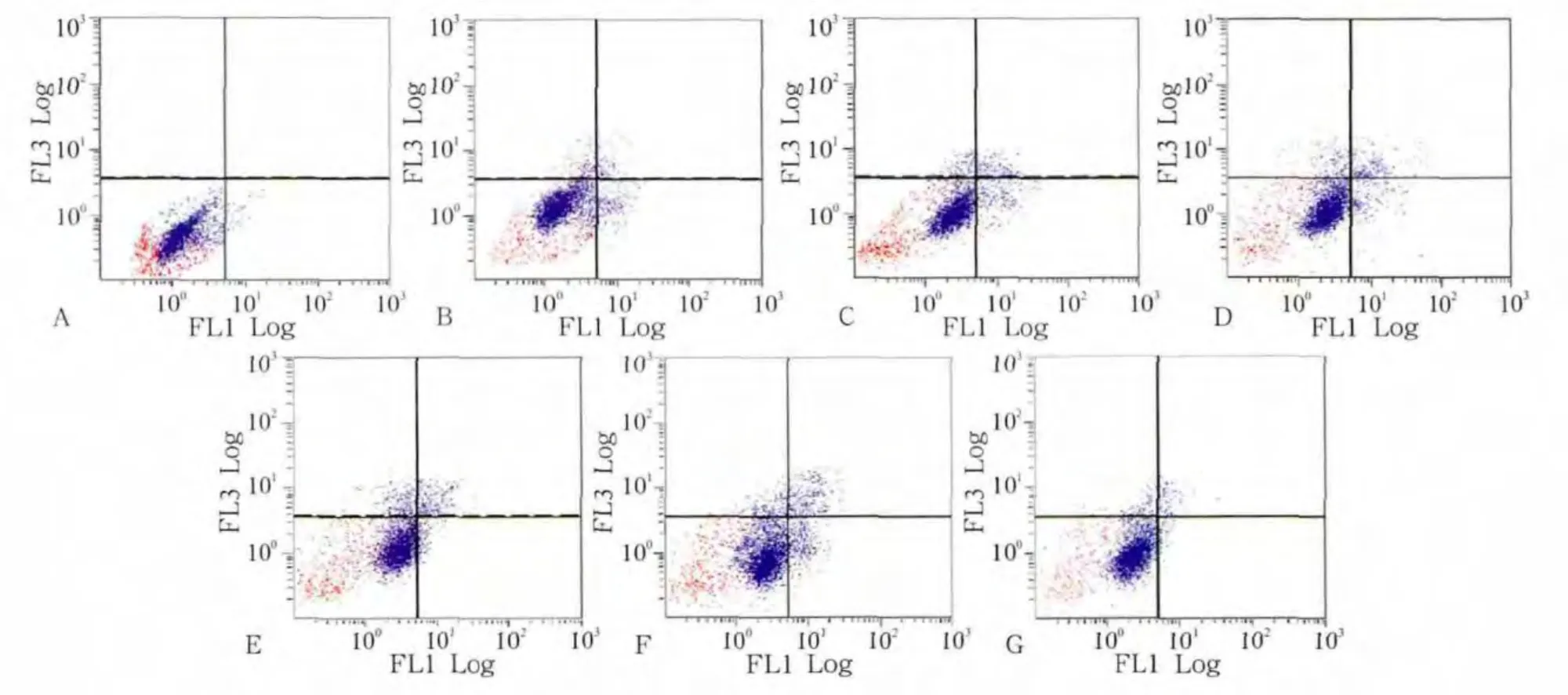
圖3 各組成肌細胞流式細胞儀檢測結(jié)果Fig 3 Flow cytometry analysis in myoblasts of every group
2.4 real-time PCR檢測CaN和NFAT的mRNA表達
real-time PCR檢測結(jié)果表明,隨著加力時間的延長,CaN亞基CnA、CnB及NFAT3的mRNA表達量逐漸升高,24 h達到峰值,與對照組相比有統(tǒng)計學差異(P<0.05);而NFAT4的mRNA表達無統(tǒng)計學差異。在相同加力時間情況下,加入抑制劑后,CnA、NFAT3的mRNA表達比不加抑制劑減少,有統(tǒng)計學差異(P<0.05),CnB的mRNA僅在加力24 h時比不加抑制劑減少,具有統(tǒng)計學差異(P<0.05),NFAT4的mRNA表達2組間無統(tǒng)計學差異(圖4)。

圖4 各組成肌細胞CaN和NFAT的mRNA表達變化的比較Fig 4 Comparison of expression of CaN and NFAT mRNA in myoblasts of every group
2.5 蛋白質(zhì)印跡法檢測NFAT3的蛋白含量
蛋白質(zhì)印跡法檢測結(jié)果表明,隨著加力時間的延長,NFAT3蛋白含量逐漸升高,24 h達到峰值,在12、24 h時與對照組相比具有統(tǒng)計學差異(P<0.05);加入抑制劑后,NFAT3的蛋白含量明顯受到抑制,與相同加力時間的對應組之間均有統(tǒng)計學差異(P<0.05)(圖5)。

圖5 各組成肌細胞NFAT3蛋白量的相對灰度值Fig 5 Relative gray value of NFAT3 protein in myoblasts of every group
3 討論
臨床上正畸醫(yī)生通過功能矯形治療對生長發(fā)育期兒童施加生物力來引起肌肉形態(tài)和功能的變化,但其具體的作用機制尚不明確。本課題前期研究發(fā)現(xiàn),內(nèi)質(zhì)網(wǎng)應激介導的細胞凋亡參與了面頜部肌肉組織的適應性改建,在此過程中Ca2+起到了十分重要的作用[5]。內(nèi)質(zhì)網(wǎng)是細胞內(nèi)重要的Ca2+處理器,可以調(diào)控細胞內(nèi)Ca2+的攝取、存儲和釋放。研究發(fā)現(xiàn),當細胞受到某些刺激時,胞內(nèi)游離的Ca2+濃度會迅速升高,從而因Ca2+超載引發(fā)內(nèi)質(zhì)網(wǎng)應激,進而發(fā)生核酸內(nèi)切酶的活化、DNA斷裂等,導致細胞凋亡;反之,當細胞外的Ca2+缺乏或者細胞內(nèi)的Ca2+螫合時,可以抑制內(nèi)質(zhì)網(wǎng)應激的發(fā)生,進而減少細胞凋亡。
CaN是一種保守蛋白,由一個相對分子質(zhì)量為61×103的催化亞基CnA和一個相對分子質(zhì)量為19×103的調(diào)節(jié)亞基CnB緊密結(jié)合而成,是目前所發(fā)現(xiàn)的唯一的一種受Ca2+/鈣調(diào)素調(diào)節(jié)的磷蛋白磷酸酶[6]。CaN介導的信號通路與細胞凋亡關(guān)系密切。通過對心肌細胞、神經(jīng)元、癌細胞及淋巴細胞系的研究[7-10]發(fā)現(xiàn),CaN被激活后具有促凋亡和抗凋亡的雙重作用。因此,在細胞類型、刺激種類或強度不同的情況下,CaN所產(chǎn)生的細胞生物學效應也可能不同。
NFAT是介導細胞內(nèi)信號轉(zhuǎn)導的一種能夠快速誘導的核因子,可以調(diào)節(jié)大量基因的表達[11-14]。目前已知的NFAT家族有5個成員,其中NFAT1、NFAT2、NFAT4主要存在于免疫組織中,NFAT3主要存在于其他組織細胞(如心臟)中[15]。NFAT的活化可以調(diào)節(jié)許多細胞因子的信號轉(zhuǎn)導,從而干預某些疾病的發(fā)生、發(fā)展。
本研究應用Hoechst 33258染色及流式細胞術(shù)檢測周期性張應力下成肌細胞凋亡情況,發(fā)現(xiàn)成肌細胞凋亡率隨應力加載時間的延長而升高,說明周期性張應力可誘導細胞凋亡。real-time PCR及蛋白質(zhì)印跡法研究結(jié)果發(fā)現(xiàn),隨著加力時間的延長,CnA、CnB及NFAT3的mRNA表達量及NFAT3的蛋白含量逐漸升高,24 h達峰值(P<0.05),而NFAT4的mRNA表達量無明顯變化,這一結(jié)果可能與NFAT4主要存在于免疫組織而在其他組織中較少表達有關(guān)。加入CaN特異性抑制劑CsA后,CnA、NFAT3的mRNA表達量及NFAT3的蛋白含量明顯受到抑制,與各自對應對照組相比有統(tǒng)計學差異,CnB的mRNA僅在加力24 h組差異具有統(tǒng)計學意義,而NFAT4的mRNA表達量無明顯變化,說明CsA可以抑制CaN催化亞基CnA的表達,但對CaN調(diào)節(jié)亞基CnB的表達僅在其濃度較高時有部分抑制作用。由此可以看出,CaN與NFAT3在應力介導的成肌細胞凋亡過程中發(fā)揮了一定作用, CaN-NFAT3信號通路可能參與了應力介導的成肌細胞凋亡,但其確切機制尚需進一步深入研究。
[1]Lee DH, Yu HS. Masseter muscle changes following orthognathic surgery: a long-term three-dimensional computed tomography follow-up[J]. Angle Orthod, 2012, 82(5):792-798.
[2]Yagci A, Uysal T, Kara S, et al. The effects of myofunctional appliance treatment on the perioral and masticatory muscles in ClassⅡ, Division 1 patients[J]. World J Orthod,2010, 11(2):117-122.
[3]Sanchez-Esteban J, Wang Y, Cicchiello LA, et al. Cyclic mechanical stretch inhibits cell proliferation and induces apoptosis in fetal rat lung fibroblasts[J]. Am J Physiol Lung Cell Mol Physiol, 2002, 282(3):L448-L456.
[4]溫葭, 陳士林, 徐宏喜, 等. 葛根素對細胞凋亡影響的研究[J]. 現(xiàn)代中藥研究與實踐, 2006, 20(1):61-64.
[5]張強, 王洪玲, 丁弦, 等. 成肌細胞體外培養(yǎng)-力學刺激模型與周期性張應力的影響[J]. 中國組織工程研究, 2014,18(5):669-674.
[6]張榮成, 王萌萌, 王建春. 鈣調(diào)神經(jīng)磷酸酶信號通路在心肌肥厚中的作用[J]. 心血管病學進展, 2007, 28(2):321-324.
[7]Bueno OF, Lips DJ, Kaiser RA, et al. Calcineurin Abeta gene targeting predisposes the myocardium to acute ischemiainduced apoptosis and dysfunction[J]. Circ Res, 2004, 94(1):91-99.
[8]Jayaraman T, Marks AR. Calcineurin is downstream of the inositol 1,4,5-trisphosphate receptor in the apoptotic and cell growth pathways[J]. J Biol Chem, 2000, 275(9):6417-6420.
[9]Tombal B, Weeraratna AT, Denmeade SR, et al. Thapsigargin induces a calmodulin/calcineurin-dependent apoptotic cascade responsible for the death of prostatic cancer cells[J]. Prostate, 2000, 43(4):303-317.
[10]De Windt LJ, Lim HW, Taigen T, et al. Calcineurin-mediated hypertrophy protects cardiomyocytes from apoptosisin vitroandin vivo: an apoptosis-independent model of dilated heart failure[J]. Circ Res, 2000, 86(3):255-263.
[11]Crabtree GR, Olson EN. NFAT signaling: choreographing the social lives of cells[J]. Cell, 2002, 109(Suppl):S67-S79.
[12]Horsley V, Pavlath GK. NFAT: ubiquitous regulator of cell differentiation and adaptation[J]. J Cell Biol, 2002, 156(5):771-774.
[13]GraefiA, Wang F, Charron F, et al. Neurotrophins and netrins require calcineurin/NFAT signaling to stimulate outgrowth of embryonic axons[J]. Cell, 2003, 113(5):657-670.[14]Hill-Eubanks DC, Gomez MF, Stevenson AS, et al. NFAT regulation in smooth muscle[J]. Trends Cardiovasc Med,2003, 13(2):56-62.
[15]Luik RM, Lewis RS. New insights into the molecular mechanisms of store-operated Ca2+signaling in T cells[J]. Trends Mol Med, 2007, 13(3):103-107.
猜你喜歡
中國設(shè)備工程(2022年12期)2022-07-11 04:33:00
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2021年6期)2021-11-22 07:50:58
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2020年12期)2021-01-18 06:57:46
中學生數(shù)理化·七年級數(shù)學人教版(2019年9期)2019-11-25 07:34:36
中學生數(shù)理化·七年級數(shù)學人教版(2019年9期)2019-11-25 07:34:34
中學生數(shù)理化·七年級數(shù)學人教版(2019年12期)2019-05-21 02:53:50
中學生數(shù)理化·七年級數(shù)學人教版(2019年12期)2019-05-21 02:53:48
