牛主要組織相容性復合物BoLA基因研究進展
2015-12-08 08:57:03向王震王璐瑤巴利民
中國獸醫雜志 2015年11期
關鍵詞:研究
向王震,肖 進,,王璐瑤,李 靜,巴利民,齊 鵬,汪 明,
(1.中國農業大學動物醫學院,北京 海淀 100095;2.農業部獸用生物制品與化藥重點實驗室北京市獸用多肽疫苗設計與制備工程技術中心 中牧實業股份有限公司研究院,北京 海淀 100070)
牛主要組織相容性復合物BoLA基因研究進展
向王震1,肖 進1,2,王璐瑤1,李 靜1,巴利民2,齊 鵬2,汪 明1,2
(1.中國農業大學動物醫學院,北京 海淀 100095;2.農業部獸用生物制品與化藥重點實驗室北京市獸用多肽疫苗設計與制備工程技術中心 中牧實業股份有限公司研究院,北京 海淀 100070)
牛主要組織相容性復合體MHC(BoLA)與移植排斥反應和免疫反應密切相關,是免疫學研究的重要內容。本文簡述了牛BoLA分布、結構及其功能,介紹BoLA與疾病易感性,免疫應答調控之間的關系及表達產物結構方面的研究。
1 簡介
主要組織相容性復合物(major histocompati?bility complex,MHC)是動物體內由緊密連鎖、高度多態的基因座位所組成的一個基因家族。MHC基因產物稱為MHC抗原,是一類存在于細胞表面的糖蛋白,主要負責連接和遞呈抗原性多肽給T淋巴細胞。因此對MHC基因表達的分析是免疫應答與調控相關研究的重要內容。
2 BoLA基因及其表達產物
Amorena B.[1]等(1978)首次報道牛的MHC I類抗原,并將牛MHC系統命名為牛白細胞抗原系統(bovine leucocyteantigen,BoLA)。此后,Letesson等發現BoLAⅡ類抗原。隨著分子生物學的發展,大量分子生物學技術手段被運用來進行BoLA基因的研究。
BOLA基因位于牛的第23號染色體[2],主要分為BoLA-Ⅰ類,BoLA-Ⅱ類,BoLA-Ⅲ類基因。Bo?LA-Ⅰ類區域包含約1 550 kb大小基因片段,且該區域有兩個緊密聯系的基因表達座位BoLA-A和BoLA-B[3]。BoLA-Ⅰ類分子幾乎分布于所有有核細胞表面。大多數組織和細胞都具有較高濃度的BoLA-Ⅰ類分子分布,其中T淋巴細胞、B淋巴細胞以及單核巨噬細胞系統的細胞濃度最高。BoLA-Ⅰ類抗原由一條α鏈和一條β鏈非共價鍵結合而成。α鏈為重鏈,分子量為44 kDa,含346個氨基酸殘基,由α1功能區、α2功能區、α3功能區、跨膜區和胞區質組成。β鏈為輕鏈,分子量為12 kDa,游離于細胞膜外通過非共價鍵與α鏈結合。BoLA-Ⅰ類分子的主要功能是進行內源性抗原的呈遞。BoLA-Ⅰ類分子的溝槽可與細胞內被蛋白酶體加工過的內源性抗原以氫鍵結合,并將其呈遞給T細胞,引發免疫反應。BoLA-Ⅰ類分子結合的多肽一般為8-11個氨基酸。
BoLA-Ⅱ類基因靠近牛23號染色體的著絲點。Ⅱ類基因分為兩個亞區-Ⅱa和Ⅱb。Ⅱa包括DR基因和DQ基因。DR基因含DRA、DRB1、DRB2和DRB3四個基因座位,DQ基因至少含有DQA1、DQA2、DQB1和DQB2等四個基因座位;Ⅱb區域包括DMA,DMB,LMP2,LMP7和TAP等基因,它們參與抗原遞呈和傳輸的過程。其他基因DNA,DOB,DIB,DYA和DYB,目前具體功能還不明確[4]。BoLA-Ⅱ類基因的編碼產物稱為BoLA-Ⅱ類分子。BoLA-Ⅱ類分子是由一條重鏈(α鏈)和一條輕鏈(β鏈)以非共價鍵結合而成的復合物。這兩條鏈都是跨膜結構,α鏈和β鏈的化學結構極為相似,只是α鏈稍大于β鏈,其分子量分別為33 kDa和28 kDa。X-衍射晶體結構分析表明α鏈和β鏈在細胞外分別含有兩個功能區α1、α2,和β1、β2,每個功能區分別由約90個氨基酸殘基組成。BoLA-Ⅱ類分子的主要作用是遞呈外源性抗原多肽,其遞呈抗原肽長度大約為12~18個氨基酸殘基,最長可達28個[5-6]。
BoLA-Ⅲ類基因由一組不同種類的基因構成,這些基因與免疫功能相關,負責編碼補體成分B因子和C4,類固醇21-羥化酶,熱休克蛋白70以及腫瘤壞死因子α和β。目前對于BoLA-Ⅲ類基因及其表達產物的研究較少[4]。
3 BOLAⅡ類a亞區DR基因
DR基因含DRA、DRB1、DRB2和DRB3四個基因座位。DRA基因編碼DR分子的α鏈,BoLA-DRA由于只有一個等位基因被報道,所以一直以來被認為是單形態。然而,近期的一些研究表明,牛
的DRA座位出現了等位變異。相比之下,編碼DR分子β鏈的DRB基因具有高度多態性。多態性主要集中在編碼多肽結合位點可變部分的第二外顯子。牛有至少3個DRB基因座位,但僅僅BoLADRB3基因是有功能的。BoLA-DRB1是假基因,在β1和橫跨膜區域攜帶終止密碼子,其基本不具多態性。BoLA-DRB2在所有基因表達時表達水平很低,不具備多態性[7]。
3.1 牛DRB3基因座位的多態性 目前研究多集中在BoLA-DRB3基因多態性方面。大量研究報道了在DRB3的第二外顯子位點存在高度多態性,并且不同品種牛該位點等位基因的頻率各不相同(表1)[6]。
DRB3基因座位的多態性研究具有重要意義,因為這個區域存在著抗原遞呈位點并且該區域的多態性可能與不同個體面對特殊病原時激發不同水平免疫應答有關。很多研究報道了某一個或多個BoLA-DRB3.2等位基因與機體對某些傳染病的抗病性的關系(表2)[6]。

表1 不同牛品種DRB3.2基因座位上頻率高于0.05的等位基因
3.2 DRB3.2基因座位的多樣性和疫苗接種 觀察BoLA-Ⅱ類DRB3基因座位等位基因的多樣性是否影響不同個體在疫苗接種后保護力的相關研究已經開展。Ballingal K T[8]等報道,在與子孢子P67抗原發生免疫反應之后,BoLA-Ⅱ型DRB3*2703和DRB3* 1501等位基因和面對小泰勒蟲威脅時的保護力水平相關。Baxter[9]等報道,BoLA-DR分子肽結合口袋對于牛口蹄疫合成肽疫苗設計意義重大。研究表明,BoLA-DRB3等位基因的結合口袋與針對口蹄疫病毒VP1基因表達的四十肽的免疫應答相關。位于DR分子多肽結合區域不同位點的幾個氨基酸和抗體應答水平重要相關。進一步的分析表明,結合口袋的特殊殘基對于疫苗設計至關重要。
3.3 MHCⅡ類分子的多肽結合位點 Srinivasan M[10]等證明,每個抗原遞呈細胞結合加工抗原的MHCⅡ類受體分子數目很少,并且這個量足以激活T淋巴細胞。另外,Chicz RM[11]等報道,利用高效液相色譜,質譜法和微測序分析法對酸洗脫的肽段與HLA-DR1結合的特點進行研究。多肽相對分子質量在1 602到2 996之間(13~25個殘基)。Mouritsen S[12]等利用等電子聚焦分離MHCⅡ類分子和其結合的自身多肽來研究MHCⅡ類分子與肽段結合的特點。他們觀察到從某一種等位基因形態的MHCⅡ類分子上洗脫下來的肽段能夠與相同等位基因形態的MHCⅡ類分子結合,而不能與不同等位基因形態的MHCⅡ類分子結合。
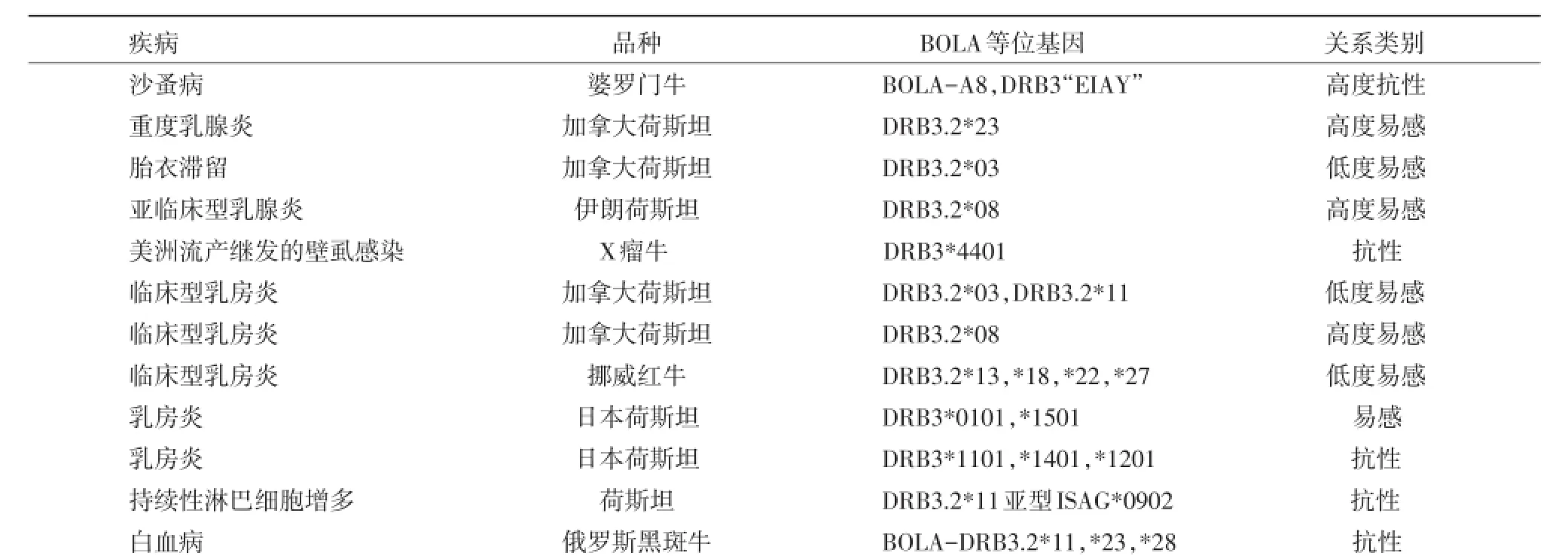
表2 在不同品種牛中不同疾病和BOLA-DRB3.2等位基因的關系
有關肽段與MHCⅡ類分子結合特點的研究具有重要意義,這將有助于我們更好的明白這種結合的方式,對于構想設計疫苗來提高動物機體面對傳染病威脅時的保護力至關重要。
4 BoLA表達蛋白產物結構分析
2010年,第一個BOLA I類分子N*01301與泰勒原蟲免疫優勢表位的結構被解析出來。對其遞呈的優勢表位構象的分析揭示了該MHC I類表型易引發強有效免疫反應的結構基礎。值得注意的是,BoLA等位基因表達產物結構分析目前在國內研究得較少。2011年,夏春[13]等利用X射線衍射技術,解析了BoLAⅠ類N*01801基因表達蛋白與牛瘟病毒來源的抗原肽復合物的晶體結構,并且分析了遞呈的抗原肽兩種不同的構象。他們發現第73位異亮氨酸是靈活可變的,允許MHC分子凹槽適應不同的構象來容納牛瘟病毒來源的抗原肽。這種宿主遞呈抗原的方式有利于創造出一個更為豐富的T細胞受體庫,這對于產生更為有效的細胞毒性T細胞免疫應答意義重大。2014年,夏春[14]等又解析了牛CD8αα分子同遞呈口蹄疫病毒來源肽段FMDV-VP1YY9的BoLA-2*02201 I類分子復合物的結構。國外有學者認為,與BoLA分子結合的抗原肽氨基酸殘基替換后能夠導致等位基因蛋白表達產物獨特空間構象發生變化,進而影響免疫應答過程中保護性抗體的表達量[15]。這對設計合成肽疫苗激發更為有效免疫應答有著重要啟示意義。目前,已報道有3個BoLAⅠ類相關分子結構被解析,而還沒有BoLA II類分子結構被解析出來的相關報道。
5 展望
MHC分子在機體免疫系統中發揮著重要作用。Ⅱ類分子廣泛的結構多態性是造成同一種群中不同個體針對傳染性病原體產生不同免疫應答的原因。而DRB3基因的第二外顯子的高度多態性可能有助于鑒別抗病性強的單倍型。對主要組織相容性復合體(MHC)的研究也有助于開發和設計含有病原體的一種或多種T細胞表位的合成肽疫苗。這將給動物機體提供更好的保護力,給未來傳染病防控提供更為有效的措施。
[1]Amorena B,Stone W H.Serologically defined(SD)locus in cattle [J].Science,1978,201(4351):159-160.
[2]Fries R,Hediger R,Stranzinger G.Tentative chromosomal local?ization of the bovinemajor histocompatibility complex by in situ hybridization[J].Animal genetics,1986,17(2):287-294.
[3]Bensaid A,Kaushal A,Baldwin C L,et al.Identification of ex?pressed bovine class IMHC genes at two loci and demonstration of physical linkage[J].Immunogenetics,1991,33(4):247-254.
[4]Lewin H A.Genetic organization,polymorphism,and function of the bovinemajor histocom-patibility complex[J].The Major Histo?compatibility Complex Region of Domestic Animal Species,CRC Series in Comparative Immunology CRC Press Boca Raton Flori?da Chapter 4,1996,65-98.
[5]Amills M,Ramiya V,Norimine J,et al.Themajor histocompati?bility complex of ruminants[J].Revue scientifique et technique(International Office of Epizootics),1998,17(1):108-120.
[6]Joshi B K.The Major Histocompatibility Complex in Bovines:A Review[J].ISRN veterinary science,2012,2012.
[7] Takeshima SN,Aida Y.Structure,function and disease suscepti?bility of the bovine major histocompatibility complex[J].Animal Science Journal,2006,77(2):138-150.
[8]Ballingall K T,Ellis SA,MacHugh N D,etal.The DY genes of the cattle MHC:expression and comparative analysis of an unusu?al class I IMHC gene pair[J].Immunogenetics,2004,55(11):748-755.
[9]Baxter R,Craigmile SC,Haley C,et al.BoLA-DR peptide bind?ing pocketsare fundamental for foot-and-mouth disease virus vac?cine design in cattle[J].Vaccine,2009,28(1):28-37.
[10]Srinivasan M,Marsh EW,Pierce SK.Characterization of natural?ly processed antigen bound to major histocompatibility complex class IImolecules[J].Proceedingsof the National Academy of Sci?ences of the United States of America,1991,88(18):7928-7932.
[11]Chicz R M,Urban R G,Lane W S,et al.Predominant naturally processed peptides bound to HLA-DR1 are derived from MHC-related molecules and are heterogeneous in size[J].Nature,1992,358(6389):764-768.
[12]Mouritsen S,Dalum I,Engel A M,etal.MHC class IIbound selfpeptides can be effectively separated by isoelectric focusing and bind optimally to their MHC class II restriction elements around pH 5.0[J].Immunology,1994,82(4):529-534.
[13]Li X,Liu J,Qi J,etal.Two distinct conformations of a rinderpest virus epitope presented by bovine major histocompatibility com?plex class IN*01801:a host strategy to present featured peptides [J].Journal ofvirology,2011,85(12):6038-6048.
[14]Wang Z,Chen R,Tariq M,et al.Complex assembly,crystalliza?tion and preliminary X-ray crystallographic analysis of the bovine CD8aa-BoLA-2*02201 complex[J].Acta Crystallographica Sec?tion F,2014,F70:742-746.
[15]Gowane G R,Sharma A K,Sankar M,et al.Association of<i>BoLA DRB3</i>allelesw ith variability in immune response among the crossbred cattle vaccinated for foot-and-mouth disease(FMD)[J]. Research in veterinary science,2013,95(1):156-163.
S823
A
0529-6005(2015)11-0072-03
2014-10-20
向王震(1990-),男,碩士,從事預防獸醫學研究,E-mail:13141319644@163.com
汪明,E-mail:vetdean@cau.edu.cn
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
