嬰兒音樂基本認知能力的發展及大腦偏側化機制研究
2015-12-06 01:30:21侯建成
音樂探索 2015年4期
宋 蓓 侯建成
(1.哈爾濱音樂學院 黑龍江哈爾濱 150028;2.俄亥俄州立大學心理學系 美國俄亥俄州 43210)
嬰兒音樂基本認知能力的發展及大腦偏側化機制研究
宋 蓓1侯建成2
(1.哈爾濱音樂學院 黑龍江哈爾濱 150028;2.俄亥俄州立大學心理學系 美國俄亥俄州 43210)
嬰兒對音高、節奏等音樂基本要素認知發展過程的研究表明,嬰兒音樂基本認知能力的發展是由音樂知覺表征發展到意義表征,在音高和節奏認知的意義表征階段出現大腦認知功能偏側化,即整體和局部信息加工分別由大腦右半側和左半側負責(具體任務分屬腦區不同)。嬰兒的音樂情緒認知具有先天性,從7個月起出現認知功能的偏側與分化。對嬰兒的音高、節奏和情緒認知發展及大腦認知功能進行研究有助于發現和揭示嬰兒音樂能力發展的規律。
嬰兒音樂基本認知;音高;節奏;情緒;大腦偏側化
一、問題的提出
音樂基本認知能力是最普遍、最基本的音樂能力。目前,對嬰兒音樂基本認知能力的研究主要關注嬰兒出生時所具有的先天性音樂認知能力和隨后天經驗增加所表現出的音樂認知能力的變化,這是探索嬰兒如何習得音樂知識與技能的重要途徑。就先天條件來說,音樂認知能力存在于所有物種之中,盡管人與動物之間的音樂認知能力大不相同,但這種能力有可能是從共同的祖先那里演化而來的,如人類某些音樂能力在一些靈長類動物中也存在①[1]。
同時,后天環境與經驗對嬰兒音樂認知能力的發展也具有重要影響。嬰兒生活經驗的增加可以促進認知能力和大腦功能的變化。表征是認知發展的重要內容,表征能力的強弱直接體現了認知發展的水平。發展心理學表明,嬰兒的表征能力由感知表征向意義表征逐步發展,最終形成概念和意義;同時,大腦功能的不斷發展分化了左右半球和各腦區之間的認知功能,對刺激任務的理解更加具體。此外,嬰兒語言能力的發展也可以促進音樂能力發展,因為語言中的音高、節奏、語調和句法等結構要素與音樂的音高、節奏、音調和曲式等要素具有一致性,并同樣對意義表達及情緒表現具有很大影響[2]。
音樂有很多構成要素,但最基本、最重要的是音高和節奏。大量文獻表明,音樂最根本作用的就是表達、傳遞和誘發情緒活動。而音高和節奏作為音樂的基本要素,是構成音樂情緒體驗的基礎,對嬰兒的音樂音高、節奏和情緒認知發展及大腦認知功能進行研究有助于發現和揭示嬰兒音樂能力發展的規律。
二、音高認知研究
(一)針對“習慣化—去習慣化”的研究
“習慣化”是指由于某一刺激重復出現而導致反應逐漸減弱,注意程度及某些生理反應(如心率、呼吸頻率等)等降低的現象。若此時一個新異刺激出現,則會導致較強刺激反應,這種現象被稱為“去習慣化”。習慣化和去習慣化的指標之一是嬰兒注意刺激時間的長短,用以研究嬰兒對不同刺激進行知覺區分的能力[3]。嬰兒對新異刺激會注意較長時間,這種注意不需要概念支持,而是對知覺原形習慣化做出的自動反應[3]。習慣化和去習慣化使嬰兒不斷關注不熟悉的刺激,這是他們進行有效學習的一條重要途徑。
Rose等研究者分別向兩組兩個月大的嬰兒呈現由協和音程與不協和音程所構成的音高輪廓,然后再呈現由不協和音程與協和音程所構成的音高輪廓,結果發現兩組嬰兒均對后者的注意時間增加[4]。Trehub向24名5個月的嬰兒依次播放6段不同的音調所組成的音高輪廓,直到嬰兒對這些音高輪廓感到熟悉(即習慣化,其心率恢復正常值),然后主試利用新的音調代替原有音調②,結果顯示嬰兒面對新的音高刺激時心率減慢[5]。此外,Dalla等研究者對1組8個半月大的嬰兒播放音樂,待其熟悉該音高刺激后,將某1個音高升高五度,然后與原有音調刺激先后呈現,他們發現嬰兒將頭轉向了音高變化的聲源方向且注意時間較長[5]。這說明,嬰兒對音高刺激熟悉之后能夠識別變化了的音高,能夠在原有音樂感知的基礎上對新的音樂刺激進行歸類,即具有音樂知覺分類的能力。
以上針對嬰兒對熟悉與新異音高刺激的比較研究表明,無論音程結構是否協和,嬰兒均能敏感地感知兩者間的變化,對新異音高刺激注意時間較長,即對原有音高刺激去習慣化的自動反應。
(二) 音高認知能力的發展
研究表明,嬰兒天生具有對協和音程與不協和音程區分能力。例如, Soken等向兩個月大的嬰兒分別呈現協和音程與不協和音程,結果發現嬰兒不僅將頭偏向了協和音程一側,并且對協和音程有著較長時間的注意,而對不協和音程表現出注意力分散的現象[6]。Robertson等發現1~2個月的嬰兒在聆聽《小步舞曲》時,對其中的協和音程予以更長時間的關注[7]。此外,Nawrot向兩組兩個月大的嬰兒分別呈現由協和音程與不協和音程所構成的曲調后,再分別呈現由不協和音程和協和音程所組成的曲調,發現兩組嬰兒對協和音程所組成的曲調的注意時間均長于不協和音程所構成的曲調[7]。
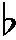
Balaban和Anderson選取40名8~10個月的嬰兒作為被試,采用的實驗材料包括由6個音高構成的標準輪廓、異輪廓和同輪廓。異輪廓和同輪廓都在標準輪廓的基礎上改變了第3個音符的音高(變化幅度為5個半音),區別是異輪廓改變了音高輪廓運動方向,屬于整體性變化,而同輪廓沒有改變音高運動方向,是局部性變化(見圖1)。主試給嬰兒呈現標準刺激后再分別呈現兩個變化刺激,結果表明嬰兒能夠辨別輪廓的變化,且對異輪廓的分辨正確率遠高于同輪廓,只有個別嬰兒將同輪廓和標準輪廓混淆[9]。而后,Ferland和Mendelson在9~10個月的嬰兒身上獲得了類似的研究結果[10]。

圖1
相對于音高同輪廓,嬰兒能更好地感知音高異輪廓。Trehub認為這是由于嬰兒對整體性的音高變化更容易作出判斷。音高異輪廓屬于整體性變化,涉及到整體性信息加工,嬰兒需按照模式對之進行提取和辨別;而同輪廓涉及局部信息加工,音高變化更多出現在局部水平上,所以異輪廓和同輪廓的認知比較體現了嬰兒對整體和局部音樂信息加工的差異[9]。8~9個月的嬰兒不僅可以感知整體信息,還可以從知覺分析和整體信息中抽取某些特征來建立局部信息。
Balaban和Anderson的實驗結果表明,嬰兒對音高的認知加工突破了只能按照音高整體輪廓與音高協和性的限制,感知了音高局部性變化的意義信息,而意義表征是以概念類別為基礎的,也就是說嬰兒具有音高的整體和局部信息概念。Quinn支持嬰兒的音高認知經歷了從簡單的知覺表征到意義表征的發展過程,認為嬰兒一開始關注音高之間的整體與協和性結構,這具有先天性,但隨著生活環境和經驗的增加,嬰兒逐漸注意內部信息特征的變化[11]。Mareschal等認為嬰兒對意義的產生最初來自其直接感知刺激物的表面特征,隨著后天經驗的增加,嬰兒獲得了不同類別特征的經驗,減少了對知覺相似性的依賴而逐漸向意義表征發展[12]。
總地說來,嬰兒能對單一物體表現出習慣化,還能感知典型的變化,初步具有分類能力。這表明,嬰兒在分析、加工、綜合各種信息時力圖理解事物的意義,探求其發展的趨勢和規律。有關原型信息的論據表明,嬰兒能根據刺激不同特征之間的關系來編碼各自獨立的感知結構,而覺察不同刺激之間意義和規律的能力將大大提高這種認知機制的價值,因為對意義和規律的覺察、把握內在知覺表征是十分重要的。
(三)音高認知的腦偏側化機制
嬰兒對音高的整體和局部認知腦機制是什么?起初,人們普遍認為人腦左半球負責抽象思維,右半球負責形象思維和知覺思維(包括音樂知覺),即不管是異輪廓還是同輪廓都分別表現出左側耳—右半球優勢。但Balaban的實驗結果顯示:音高整體性變化時,嬰兒對異輪廓變化具有左側耳—右半球對優勢,對同輪廓具有右側耳—左半球優勢,表現出音高認知的偏側化機制[9]。一些神經心理學家認為左半球在音樂局部線索中很重要,負責“局部”信息處理;而右半球在整體線索中扮演重要角色,負責音樂“整體”信息處理。Brancucci把音樂旋律分成音高輪廓亞系統和音高結構(音程)亞系統,發現右側腦在音高輪廓方面發揮主要作用,而左側腦在音程結構上發揮作用(但需要完整右側半球支持)[13]。研究者相信,這種音樂結構感知的偏側化模式可作為半球聽覺感知功能特異性指標。
同時,研究者認為整體和局部信息加工的半球不對稱功能主要出現于高級知覺加工階段,整體和局部信息表征不僅需要高級知覺腦區(左右側顳葉皮層)參與,還需要頂葉皮層在不同水平之間進行注意監控,這可能是半球不對稱的主要原因[14],但這種解釋還存在爭議。Zatorre認為負責音高加工的腦區位于右側顳葉,Schon也發現音高加工主要位于右側顳—頂葉區[15]。當然,在這些刺激任務中,音高輪廓的音程比較大(至少三度以上),所以被試比較容易識別整體性變化,如果音程關系比較小,則可能會出現相反的實驗結果。筆者認為,對于音高的整體和局部加工是一般性整體和局部信息的一部分,其偏側化機制符合“右半球加工整體信息,左半球加工局部信息”。但是,整體和局部信息加工受到多種復雜因素調節,也隨具體實驗目的和條件不同而結果不一,同時有必要進一步研究個體變量因素的影響。
除整體和局部因素外,筆者認為音高的語義性也是一個重要因素。音樂是語言的一種特殊表現形式,包括音高在內的音樂結構對音樂傳達的意義及情緒有著很大影響,許多ERP③研究發現,改變旋律中的某些音高會在右側顳上回和頂葉出現N400④,且變化越明顯則波幅也越大。在前后情境下音高能啟動相關詞匯意義,音高變化也改變了樂段表達的語義信息,使當前的相關啟動詞與原先意義不匹配而誘發N400[2]。音高變化與音樂結構也有關聯,結構不穩定時在中央頂區出現P600⑤,變化越明顯則波幅越大且潛伏期越短[2]。音高變化改變了樂段前后意義聯系和結構性,類似語言加工的語義不合適或類別錯誤(N400)和句法結構違反(P600)。另外,從語言能力發展上看,Colombo和Bundy發現,大約8~10月的嬰兒不再把聲音看作單獨的個體組成,而是開始表征語音對應關系理解語言的意義[16]。可以認為在Balaban等的研究中,8~10個月的嬰兒不僅能夠感知音高輪廓的整體和局部信息,還感受了內在音樂結構及音樂語義信息。
總之,上述研究證實,兩個月之前的嬰兒偏向協和、標準化結構的音程,這具有一定的先天性;而3~4個月的嬰兒開始能夠辨別協和和不協和音高輪廓;5個月的嬰兒敏感于對熟悉(或適應)音樂的音高變化,不管此時音高的音程及結構是否協和,這是對知覺原形的習慣化自動反應,而不需要概念支持;6、7個月的嬰兒能夠分辨前后音高距離(音程) 的大小,即使對音高轉換后仍感受到差異;從第8個月起,嬰兒對同輪廓和異輪廓旋律都能做出敏銳判斷,這表明嬰兒開始感知整體信息并從中抽取某些特征建立局部信息,左右半球的顳—頂葉區分別負責局部和整體信息處理,表現出音高認知功能的大腦偏側化。也可能與音高變化改變音樂意義及結構有關,反映了嬰兒感知音樂表達的內在結構及語義信息。
三、節奏認知
(一)節奏認知能力的發展
節奏被譽為音樂的靈魂,是構成音樂最根本的要素之一。關于對節奏的認知,Pickens等讓1組兩個月大的嬰兒適應一段2/4拍子的節奏刺激后,再呈現3/4拍子節奏刺激,嬰兒對后者予以注意并增加了注意時間,他們認為嬰兒在兩個月之前就能感知音樂節奏及其變化[17]。這表明嬰兒熟悉一段節奏刺激后分辨了新異節奏刺激,并能在原型感知基礎上進行分類,表現出知覺分類的能力是習慣化后的一種自動反應。此后,Trehub等觀察了3~5個月嬰兒的節奏認知能力,選擇兩段三拍子的節奏刺激,時長分別為600-200-400和200-600-400毫秒,嬰兒對此表現出辨別兩種不同節奏刺激的傾向[18]。Hannon認為這是因為嬰兒能根據節拍和時值分辨不同節奏,根據節拍類型和時值長短做出不同的反應[18]。此年齡段的嬰兒是非概念有機體,對音樂節奏的知覺反應建立在某些模塊匹配基礎上,根據節奏輪廓識別不同節奏及變化。
在此基礎上,Hannon和Johnson進一步測查了6~9個月嬰兒的節奏識別能力,他們選取3~5和6~9個月兩組嬰兒被試,向被試依次呈現一小節四拍子和三拍子的節奏模式,然后分別對兩種節奏中的某些拍子改成實拍和空拍(圖2),結果表明,嬰兒不但能辨別兩種不同的節奏,而且還敏感于每種節奏的節拍變化,對實拍和空拍的轉變表現出強烈的新異傾向[18]。這反映出嬰兒不僅能夠感知整體節奏輪廓變化,還能感知節奏局部信息變化。此外,節奏輪廓屬于整體性結構,涉及到整體信息加工,而節拍屬于局部性信息加工,所以嬰兒需按照整體模式和某些局部特征來分辨節奏的差異。這也表明嬰兒對節奏的認知加工也突破了節奏整體輪廓的限制,感知了節奏局部信息變化,形成了節奏的整體和局部信息概念。
除了節奏的整體輪廓與局部信息外,筆者認為,嬰兒通過生活經驗建立了音樂與表現對象之間的關系,即聯覺反應。例如,時值較長(節奏較慢)

圖2
表現了大、慢、沉重的對象,時值較短(節奏較快)則表現小、快、靈巧的對象,介乎二者之間則會產生從容、適意、自然的體驗,所以嬰兒通過日常經驗來理解音樂意義。節奏的時值長短差異造成音樂運動張力不同,時值較長的張力較小,時值較短的張力較大,兩者具有反比關系,因而時值對節奏性質產生影響,也就是說某1個或幾個音符時值之間的跨距被改變時,節奏性質以及音樂表現意義常常隨之變化。另一方面,音樂的節奏是語言節奏的變形,能直接反映出類似語言的語義信息[19]。因此,同音高一樣,節奏也能傳達類似語言的意義,嬰兒從具體音響形態的分析中獲得節奏這個要素,通過時值和節拍提取音樂表達意義。
除節拍和時值以外,Hannon和Trehub認為嬰兒也能夠從節拍重音判斷出節奏差異[20]。重音是節奏及節拍的中心點,具有位置固定和靜態特征[19]。這種特征使得音樂具有律動感并能吸引嬰兒的注意。Palmer等觀察到10~12個月的嬰兒在同樣的節奏中對節拍重音予以更多和更長的注意時間[21]。這表明,嬰兒通過不同形態和類型的重音來確定節奏的運動方式并把握不同節奏運動形態,認知了具有節拍重音的節奏刺激的不同分組和意義。所以嬰兒從10個月起不但能分辨節奏型輪廓和識別節奏意義,而且能夠通過節拍重音感知節奏律動加工并識別和分類不同節奏。
筆者認為,嬰兒除了通過節拍重音確定節奏運動方式并把握節奏運動特點外,還與重音本身特征有關。第一,從言語學習看,相關研究表明言語加工的重音是言語學習的節奏或拍子,在語言環境中表示強調。同時,ERP研究發現,語言重音后面的CPS⑥成分往往反映出個體對語調短語邊界的加工,同時反映出一種與記憶和注意有關的過程,即引導人們將注意焦點從一個短語轉移到下一個短語,從而繼續對后面短語進行加工[22];第二,Knosche和Neuhaus等使用ERP在音樂的重音、休止等處都發現與語言一致的CPS,這反映了音樂節奏更高級的認知加工,而不只是感知到重音和停頓等因素[22];第三,重音在音樂中表示突出和強調,通過響度、音高等因素與前后音符建立意義聯系,反映了音樂邊界加工和聲學邊界征兆的自下而上加工[23]。因此,嬰兒通過節拍重音來解釋和組織輸入的音樂信息,并表現出對新異刺激注意和時間增加的外顯行為。
(二)節奏認知的腦偏側化機制
通過左側腦損傷患者出現的節奏識別混亂現象,Zatorre認為節奏(包括節拍和時值)屬于左側半球加工[24],Midorikawa等通過神經成像認為節奏屬于左側半球加工[24]。Wilson等經過腦損傷和神經成像研究后認為,節拍和節奏分屬不同腦半球加工,節拍屬于右側半球加工,節奏屬于左側半球加工[25]。這可能是由于腦損傷患者需要分別調動右半球和左半球的相應功能來補償節拍和節奏感知的缺失。Sakai等通過對正常人的研究發現,節拍加工依賴于左側半球的初級運動皮層和頂葉皮層,而右側半球的前額葉和頂葉皮層負責時值和節奏加工[24]。筆者認為,節奏的整體和局部加工機制與音高以及其它認知任務的整體和局部加工機制具有一致性,音高輪廓的整體信息加工需要右側半球支持,局部信息加工需要左半球支持;在其它認知任務中,Han等在視覺任務加工中也發現,整體和局部性質加工也需要相應的右半球和左半球加工[14]。盡管具體任務誘發的腦區活動不盡相同(音高涉及顳葉和頂葉皮層、節奏涉及初級運動和頂葉皮層、視覺涉及頂葉和枕葉皮層等),但都表現了局部—左半球和整體—右半球偏側化傾向。此外,研究者還觀察到以上認知任務都誘發了頂葉神經活動,這可能是頂葉不同子區域選擇性地加工了認知信息。Sack認為頂上區與縮小注意范圍使之聚焦有關,頂下區與擴大注意范圍有關;注意范圍變化時分別引起左側和右側頂葉活動,同時涉及任務信息的短時儲存、維持以及信息注意控制[26]。此外,嬰兒的生活和音樂經驗非常少,因此相關ERP研究發現音樂學習經驗貧乏者加工音樂節拍時,左側頂區的N100比音樂經驗豐富者要小[27],據此可以推測出嬰兒加工節拍時左側頂區有著較小的活動強度。
總之,嬰兒的節奏認知能力與音高認知能力有著相似的發展特征,兩個月及之前對某一節奏適應后能區分不同的節奏輪廓,這具有先天性;3~5個月的嬰兒能通過節拍和時值辨別不同節奏,對節奏進行簡單辨別和歸類;6~9個月的嬰兒不僅能夠感知整體節奏輪廓變化,還能感知節奏局部信息的變化,這表明其從簡單知覺分類過渡到注意刺激內部意義,識別音樂節奏意義。10~12個月的嬰兒能通過節拍重音關注節奏特征,感知節奏律動并確定節奏運動方式及運動形態。在大腦加工機制方面,節拍依賴于左半球的初級運動皮層和頂葉皮層加工,時值和節奏依賴右半球的前額葉和頂葉皮層加工,這與其它認知任務的整體和局部加工偏側機制相同。
四、音樂情緒認知的發展
旋律由音高和節奏所組成,被比喻為音樂的靈魂,是表達情緒之藝術語言的主要手段,因此人們往往通過旋律來探討音樂誘發情緒的心理表現。Gentile比較了3~8個月嬰兒的音樂情緒體驗,他在被試的左右兩側分別呈現面部表情愉快與不愉快的毛絨動物玩具,同時呈現音樂旋律訓練嬰兒在正性音樂情緒(高興、歡快等)時注意愉快面孔玩具,負性音樂情緒(悲痛、憂傷等)時注意不愉快面孔玩具;在正式實驗時向被試依次呈現新的正性和負性情緒的音樂,同時觀察嬰兒的面部運動(注意)情況。結果表明,5~8個月嬰兒出現明顯的外顯行為反應且正確率高,但3~4個月的嬰兒沒有外顯行為,還有個別嬰兒混淆了愉快和不愉快面孔[28]。因此,Gentile認為嬰兒從5個月起開始學會識別音樂的情緒意義,5個月之前的嬰兒還不具有這種能力。
從嬰兒的音高認知發展來看,3~5個月的嬰兒能識別音高輪廓變化,能進行簡單的歸類,6~9個月的能感知整體和局部的音高變化且識別局部信息意義,即表現出從簡單知覺分類過渡到整體和局部信息特征的建立。也就是說,在Gentile的實驗中,5個月的嬰兒不但能感知音高輪廓,還能感知音高之間的局部結構信息,這以調式結構體現。從正性和負性情緒的調式結構上看,正性情緒音樂基本由大調式構成,包含較多低緊張度音程,而負性情緒音樂大多由小調式構成,包含較多高緊張度音程[29],因而音程形成的大、小調不同會產生不同的情態特征體驗。在Gentile的實驗中,嬰兒面對玩具的面孔表情時,正性音樂情緒與愉快面孔相對應,負性音樂情緒與不愉快面孔相對應,這是因為音樂情緒體驗與其表現對象之間具有某種對應關系,兩者通過“聯覺”這種心理活動建立聯系,這種聯覺活動并非是個別的個體經驗而是一種普遍性的人類心理活動規律,也是人類心理活動的自然規律[30]。
Gentile在實驗中僅觀察了嬰兒的外顯行為反應,認為嬰兒只有發展到一定年齡階段才產生情緒反應。而Thompson把情緒視作先天和后天作用共同形成的,從先天性來說正性和負性情緒在人類進化過程中體現不同的適應意義[31]。Denham則認為剛出生嬰兒的情緒容易受到外界刺激影響,盡管不會有明顯的顯現[32]。所以,研究嬰兒的情緒反應機制需要突破單純的外顯行為研究來觀察。Schmidt等通過EEG⑦的α波(8~13Hz)觀察了音樂誘發嬰兒情緒的內在生理機制[33]。他們選擇3、6、9和12月大的嬰兒被試各一組,向每組被試依次呈現快樂、悲傷和恐怖的音樂,觀察其大腦左右兩側的額葉(雙側額中回F3、F4)和頂葉(雙側頂葉中部P3、P4)的活動情況。結果發現,出現音樂刺激時,3和6個月大嬰兒的雙側額葉和頂葉α波頻低于基線(無任何刺激)的α波頻率,即雙側額葉和頂葉活動強度高于基準線條件下的活動水平(α波與大腦活動呈負相關,即α波越高則大腦活動程度越低,α波越低則大腦活動程度越高),這表明3個月的嬰兒已能夠體驗音樂情緒,但左右兩側的額葉和頂葉α波頻率均沒有顯著差異;而9個月大嬰兒的雙側額葉和頂葉α波頻率不但低于基線的頻率,且雙側額葉α波頻率低于雙側頂葉,這表明其活動強度也高于頂葉;12個月大嬰兒的雙側額葉活動強度高于雙側頂葉,同時左側額葉α波頻率低于右側額葉,表明左側活動強度高于右側。
嬰兒的大腦從一出生就不斷發展,各個腦區的功能不斷分化并相互協調。大量研究表明,大腦額葉是情緒感知的重要部位。嬰兒從出生后1個月起,大腦額葉錐體神經元和中間神經元就開始發育,同時伴隨著外側和背外側額葉的發育,但同時頂葉也不斷發展,額、頂葉分化活動相差不大[34]。Huttenlocher等發現,嬰兒從出生到6個月期間,大部分腦區域神經連接分化程度不高,但7~12個月間神經連接密度增長較快且出現腦區功能分化[35];Chugani等通過PET⑧測量嬰兒大腦的神經連接作用后得出相似結論[36]。所以,在Schmidt的實驗中,盡管3個月和6個月大嬰兒的雙側額葉和頂葉在音樂情緒中發揮了作用,但由于大腦功能分化度不高,故沒有出現差異。9~12個月時,由于大腦神經連接的迅速發展,大腦功能分化程度明顯增加,負責情緒認知加工的額葉的活動強度顯著高于頂葉的活動強度。
伴隨著左右額葉功能的進一步分化,左側額葉活動與右側額葉活動也有所不同。Heilman認為大腦雙側額葉對情緒種類的辨別具有重要作用,左側調節正性情緒,右側調節負性情緒[37]。Zatorre等通過PET也發現正性和負性音樂情緒分別激活了額葉部位的左側和右側,而且激活的程度越強烈,相對應的大腦兩側活動強度也越高[38]。在Schmidt的實驗中,嬰兒大腦左側額葉的活動強度高于右側強度,這表明嬰兒更多體驗了快樂音樂的情緒。Cummings等在1995年的研究發現,嬰兒在體驗到憤怒和悲痛時消極情緒較少,但有著更加積極的情緒調控,同時研究者認為除情緒特征外,嬰兒本身就存在一定調節能力,積極情緒的調控和形成與嬰兒和外界交際行為環境,特別是與母親的關系緊密相關[39]。Roberts和Strayer研究認為,具有先天適應環境和模仿學習本能的嬰兒通過母親與嬰兒之間的交往行為(安撫、語言對話等)來感知、發展和控制情緒表現,這種“安全型親子關系”的建立促進了嬰兒積極情緒的發展和形成[39]。這不僅滿足了嬰兒的生理需要,還提高了與嬰兒情感交流的積極性與主動性。所以Schmidt發現呈現音樂旋律刺激時,嬰兒受到正性情緒的影響而激活左側額葉,而通過某些情緒調節策略抑制了由悲傷和恐怖音樂誘發的負性情緒,這也有可能是嬰兒將日常習得的情緒加工和調控策略遷移到音樂情緒認知活動中,表現出積極的音樂情緒。
負責情緒加工的主要腦區除了額葉中回以外,前額皮層也是重要區域,邊緣系統的前扣帶回、海馬、杏仁核等也發揮一定作用。前額皮層左側參與正性情緒的加工,右側更多參與負性情緒加工,左半球會對右半球作用的負性情緒給予一定抑制[40],所以額葉對情緒加工的不對稱性在人類情緒調節方面起著關鍵作用,使情緒保持平衡狀態。前扣帶回用來評價、調節或有意識地調整情緒反應,是情緒執行系統的一部分。邊緣系統的海馬和杏仁核共同處理情緒信息,兩者分別對認知任務進行評價并通過兩者之間神經通路調節情緒[41]。前扣帶回、海馬等部位的情緒反應也會帶動杏仁核反應,使杏仁核得到激活[42]。這些都是基于成人的音樂情緒認知機制的研究,那么嬰兒的音樂情緒認知的發育機制、各機制之間如何相互作用及規律等問題有待進一步探討。
五、總結與展望
綜合以上研究結果,我們得出如下結論:第一,嬰兒天生具有音樂感知能力,并且隨年齡和生活經驗的增加,音樂認知能力不斷發展。第二,音高和節奏是音樂的兩個基本要素,嬰兒對其有著相似的認知發展過程:兩個月及之前的嬰兒能區分不同的音高和節奏輪廓,3~5個月的能對音高輪廓和節奏進行簡單的歸類,這是一種習慣化自動反應而不需概念支持;6~9個月的可以感知整體和局部音高輪廓和節奏的信息變化,能識別局部信息意義;10~12個月嬰兒可以通過音高和節奏的各種特征確定其運動的形態及意義,特別是音高變化與改變音樂語義及結構有關,這反映了嬰兒感知音樂內在結構及語義信息。第三,由音高和節奏組成的音樂旋律誘發情緒的腦機制研究中發現,6個月之前的嬰兒已具有情緒反應,腦區功能之間的分化度不高且外顯行為不明顯;9~12月個嬰兒的大腦額葉迅速發育,活動強度高于頂葉活動強度,左右額葉功能出現分化,左側的積極情緒活動強度高于右側的消極活動強度,這種積極情緒的調控和形成與嬰兒和外界交際環境特別是母嬰關系緊密相關,提高了嬰兒情感交流的積極性與主動性。第四,在大腦認知機制方面,音高和節奏都表現出一定偏側化機制,嬰兒對音高整體輪廓和節奏整體性加工表現出左側耳—右半球優勢,對音高局部性和節奏中節拍成分的加工表現出右側耳—左半球優勢,這些與其它認知任務的整體和局部加工的偏側機制相同。具體任務誘發的腦區活動不盡相同,音高加工涉及顳葉和頂葉皮層,節奏加工涉及初級運動和頂葉皮層。這種音樂結構感知的偏側化模式可作為半球聽覺感知功能特異性指標。
總之,音樂認知涵蓋的內容很多,僅研究嬰兒對音樂的音高、節奏以及情緒感知能力發展遠不能滿足要求。嬰兒的音樂認知能力與兒童、青少年和成人的音樂認知能力發展是延續性的,嬰兒可能運用不同的神經通路和加工策略來獲得并保存音樂信息,借用認知神經科學范式來研究嬰兒的音樂認知神經基礎有助于這一問題的解決,但面臨的問題是某些方法和范式是否適用于嬰兒。因此,未來借鑒嬰兒認知心理學發展的研究,在現有理論基礎上采取更好的研究設計,改進和采用算法更先進、精確并適合嬰幼兒的功能神經影像將有助于對嬰兒音樂認知能力的深刻和全面的理解。
(兩位作者對本文均同等貢獻,并列第一作者)
注釋:
① 蝙蝠、鯨類等脊椎動物和鳥類都能夠感知單音,并且這種能力隨季節變化而變化,以達到求偶、結伴以及傳遞信息的目的;與人類親緣最為接近的黑猩猩和大猩猩等動物具有辨別八度音階以內音高的能力。
② 主試重新排列這6段音調并改變其呈現順序,第1個音調與原有第1個音調相同,因為過去實驗中以具有明顯差別的音調作為開頭,很難確定使嬰兒做出反應的原因是整體的音高輪廓還是第1個音高。
③ Event-related Potentials的縮寫,即事件相關電位,是一種在神經科學研究領域廣泛應用的腦電技術,其通過外加特定的刺激于感覺系統或腦的某一部位來觀察腦區的電位變化。
④ N400是經典的腦電信號之一, 是1個大振幅的負向波,最大值達到刺激出現后約400毫秒的時間范圍內。
⑤ P600是常用的ERP指標之一,是1個大振幅的正向波,通常出現在刺激后的500-600毫秒之間。
⑥ Closure positive shift的縮寫,指終止正漂移,是特異性腦電成分之一。
⑦ Electroencephalograph的縮寫,即腦電圖。
⑧ Positron emission tomography的縮寫,即正電子發射計算機斷層掃描。
[1] Benitez B L. The Biology of Music[J]. Science, 2001, 29 (292): 2432-2433.
[2] 侯建成,劉昌. 語言與音樂活動的腦機制研究述評[J]. 心理科學進展,2008,16(1):50-58.
[3] Quinn P C, Eimas P D. The emergence of category representations during infancy: are separate perceptual and conceptual processes required? [J] . Journal of Cognition and Development , 2000(1):55-61.
[4] Rose S A, Feldman J F, Jankowski J J. Processing speed in the first year of life: A longitudinal study of preterm and full-term infants[J]. Developmental Psychology, 2002, 38(6): 895-902.
[5] Trehub S E. Musical predispositions in infancy: An update. In Peretz I. & Zatorre R. J. (Eds.), The Cognitive Neuroscience of Music[M]. Oxford: Oxford University Press,2003:3-20.
[6] Soken N H, Pick A D. Infants’ perception of dynamic affect expressions: Do infants distinguish specific expressions?[J]. Child Development, 1999,70: 1275-1282.
[7] Robertson S S, Bacher L F, Huntington N L. The integration of body movement and attention in young infants[J]. Psychological Science, 2001, 12(6): 523-526.
[8] Plantinga J, Trainor L J. Memory for melody: infants use a relative pitch code[J]. Cognition, 2005, 98(1): 1-11.
[9] Marie T, Balaban M T, Anderson L M, et al. Lateral asymmetries in infant melody perception[J]. Developmental Psychology, 1998, 34(1): 39-48.
[10] Ferland M B, Mendelson M J. Infants' categorization of melodic contour[J]. Infant Behavior and Development, 1989, 12(3): 341-355.
[11] Trainor L J, Tsang C D, Cheung V W. Preference for sensory consonance in 2- and 4- month-old infants[J]. Music Perception, 2002, 20(2): 185-192.
[12] Mareschal D, French R M, Quinn P C. A connectionist account of asymmetric category learning in early infancy[J]. Developmental Psychology, 2000, 6: 635-645.
[13] Brancucci A, SanMartini P. Laterality in the perception of temporal cues of musical timbre[J]. Neuropsychological Chologia, 1999, 37(13): 1445-1451.
[14] 張小聰,蔡厚德. 整體/局部加工中的半球功能不對稱效應[J].心理科學進展, 2008, 16(2):228-233.
[15] 侯建成, 劉昌. 國外有關音樂活動的腦機制的研究概述——兼及“莫扎特效應”[J].中央音樂學院學報, 2008(1): 110-118.
[16] Colombo J, Bundy R S. Infant response to auditory familiarity and novelty[J]. Infant Behavior, 1983, 6(2-3): 305-311.
[17] Pickens J, Bahrick L E. Do infants perceive invariant tempo and rhythm in auditory-visual events? [J] Infant Behavior and Development, 1997, 20(3): 349-357.
[18] Hannon E E, Johnson S P. Infants use meter to categorize rhythms and melodies: Implications for musical structure learning[J]. Cognitive Psychology, 2005, 50(4): 354-377.
[19] 張巍. 論節奏與節拍的關系——對二者概念的評價與討論[J]. 黃鐘, 2005(1): 64-73.
[20] Hannon E E, Trehub S E. Tuning in to musical rhythms: Infants learn more readily than adults[J]. PNAS, 2005, 102 (35): 12639-12643.
[21] Palmer C, Jungers M K, Jusczyk P W. Episodic memory for musical prosody[J]. Journal of Memory and Language, 2001, 45(4): 526-545.
[22] Knosche T R, Neuhaus C,Haueisen J. The perception of phrase structure in music[J]. Human Brain Mapping,2005, 24(4): 259-273.
[23] Nan Y, Knosche T, Friedericia A D. The perception of musical phrase structure: A cross-cultural ERP study[J]. Brain Research, 2006, 1094(1): 179-191.
[24] Bengtsson S L, Ullen F, Ehrsson H H, et al. Listening to rhythms activates motor and premotor cortices[J]. Cortex, 2009, 45(1): 62-71.
[25] Trainor L J, Gao X Q, Lei J J. The primal role of the vestibular system in determining musical rhythm[J]. Cortex, 2009, 45(1): 35-43.
[26] Sack A, Hubl D, Prvulovic D, et al. The experimental combination of rTMS and fMRI reveals the functional relevance of parietal cortex for visuospatial functions[J]. Cognitive Brain Research, 2002, 13 (1): 85-93.
[27] Vuust P, Roepstorff A, Wallentin M, et al. It doesn’t mean a thing... keeping the rhythm during polyrhythmic tension, activates language areas (BA47) [J]. NeuroImage, 2006, 31(2): 832-841.
[28] Flom R, Gentile D A, Pick A D. Infants’ discrimination of happy and sad music[J]. Infant Behavior & Development, 2008, 31(4): 716-728.
[29] Petetz I, Gagnon L, Bouchard B. Music and emotion: perceptual determinants, immediacy, and isolation after brain damage[J]. Cognition, 1998. 68(2): 111-141.
[30] 周海宏.音樂與其表現的世界[M]. 北京:中央音樂學院出版社, 2004: 130-150.
[31] Thompson R A. Emotion regulation and emotional development [J]. Educational Psychological Review, 1991,3(4):269-307.
[32] Brookes H, Slater A, Quinn P C, et al. Three-month-old infants learn arbitrary auditory-visual pairings between voices and faces[J]. Infant and Child Development, 2001, 10(1-2): 75-82.
[33] Schmidt L A, Trainor L L, Santesso D L Development of frontal electroencephalogram (EEG) and heart rate (ECG) responses to affective musical stimuli during the first 12 months of post-natal life[J]. Brain and Cognition,2003, 52: 27-32.
[34] Picton T W,Taylor M J. Electrophysiological evaluation of human brain development[J]. Developmental Neuropsychology, 2007, 31(3): 249-278.
[35] Diener M L, Mangelsdorf S C, McHale J L, et al. Infants’ behavioral strategies for emotion regulation with fathers and mothers: associations with emotional expressions and attachment quality[J]. Infancy, 2002, 3(2):153-174.
[36] Flom R, Bahrick L E. The development of infant discrimination of affect in multimodal and unimodal stimulation: The role of intersensory redundancy[J]. Developmental Psychology, 2007, 43(1), 238-252.
[37] Heilman K M. The neurobiology of emotional experience[J]. Journal of Neuropsychiatry and Clinical Neuroscience 1997, 9(3) :439-48.
[38] Zatorre R J, Bermudez P. Emotional responses to pleasant and unpleasant music correlate with activity in limbic brain regions[J]. Nature Neuroscience, 1999, 2(4): 382-387.
[39] 陸芳, 陳國鵬. 兒童情緒調節的發展研究[J].心理科學, 2003, 26(5): 928-930.
[40] Heller W. Neuropsychological mechanisms of individual differences in emotion , personality and arousal[J]. Neuropsychology, 1993, 7: 476-489.
[41] Maguire E A, Frith C D. Lateral asymmetry in the hippocampus response to the remoteness of autobiographical memories[J]. Journal of Neuroscience ,2003, 23(12): 302-307.
[42] Gehring W J, Goss B, Coles G H, et al .A neural system for error detection and compensation[J]. Psychological Science, 1993, 4(6): 385-390.
Infants' Development of Basic Cognitive Ability of Music and Its Function of Brain's Lateraluty
SONG Bei HOU Jiancheng
It introduces the developmental process of infants' cognitive development of basic music elements——— pitch,rhythm and emotion,pointing out that infants' basic cognitive development of musical ability is from perceptions- based representation to meanings-based representation. They appear the cerebral cognitive functional laterality,meaning that global and local information process belongs to right and left hemisphere ( special tasks belongs to different brain areas) . Infants' cognition of musical emotion has congenital character,and appears cognitive functional laterality and differentiation form the 7-month. These researches are helpful for discovering the developmental disciplinarian of infants' musical ability.
infant; basic cognition of music; pitch; rhythm; emotion; brain laterality
J603
A
1004-2172(2015)04-0129-09
10.15929/j.cnki.1004-2172.2015.04.020
2015-02-26
2011年國家留學基金委公派留學項目([2011]5025);2014年黑龍江省哲學社會科學研究規劃一般項目“音樂能力與語言材料類型對第二語言記憶的影響研究”(14B087)。
宋 蓓(1977— ), 女,碩士生導師,華東師范大學心理學流動站博士后,哈爾濱音樂學院教授。
侯建成(1980— ), 男,博士,美國俄亥俄州立大學心理學系博士后研究員。
責任編輯:錢芳
猜你喜歡
小天使·一年級語數英綜合(2020年3期)2020-12-16 02:56:12
藝術啟蒙(2018年7期)2018-08-23 09:14:16
風流一代·青春(2018年4期)2018-04-19 16:18:48
風流一代·青春(2018年3期)2018-03-15 17:07:26
風流一代·青春(2018年2期)2018-02-26 15:27:06
風流一代·青春(2017年6期)2018-02-14 19:28:55
風流一代·青春(2017年5期)2018-02-14 09:32:37
兒童繪本(2017年24期)2018-01-07 15:51:37
東方藝術·大家(2016年6期)2016-09-05 07:30:56
商業評論(2014年6期)2015-02-28 04:44:25
