絲光綠蠅不同蟲態附著蠟蚧輪枝菌分生孢子的能力及與其體表結構的關系
2015-12-03 05:55:34龐仁乙章一鳴李正躍吳國星
環境昆蟲學報 2015年1期
龐仁乙,高 熹,章一鳴,譚 清,李正躍,吳國星
(云南農業大學植物保護學院,昆明 650201)
蠟蚧輪枝菌Verticillium lecanii 屬于半知菌亞綱絲孢菌類輪枝菌屬匍匐組,是一種寄主范圍廣的蟲生真菌。通過對罹病棕尾別麻蠅Boettcherisca peregrina 蟲體上的病原微生物進行分離純化,找到了適合蠅類害蟲生物防治的蠟蚧輪枝菌KMZW-1菌株(王凌超等,2010)。為了安全有效地防治分布廣泛且常見的一種半住區性蠅種——絲光綠蠅Lucilia sericata,蠅蛆病病原菌攜帶者,筆者用高濃度蠟蚧輪枝菌分生孢子懸浮液分別處理其成蟲、蛹和幼蟲,結果對成蟲有較高的致病性而對幼蟲和蛹較差(未發表資料)。另外,Sharififard 等(2011)也有報道,球孢白僵菌Beauveria bassiana和金龜子綠僵菌Metarhizium anisopliate 兩種蟲生真菌對家蠅Musca domestica 成蟲、幼蟲和蛹同樣呈現致病差異性。然而,目前關于蟲生真菌對同一昆蟲不同蟲態間的致病性差異性的機理少見報道。
蟲生真菌致病昆蟲的過程一般包括分生孢子附著、萌發和穿透寄主表皮以及菌絲段在血腔內增殖等步驟(周燚等,2006)。蟲生真菌侵染成功的首要條件是分生孢子牢穩地附著在寄主體壁上。附著的分生孢子萌發后入侵蟲體,當侵入的基數超過一定量時才能有效地克服寄主的免疫抵抗,且侵入的孢子數愈多蟲體愈易發病致死(周燚等,2006)。在某特定生態條件下,附著的分生孢子萌發率相對穩定時,入侵寄主的分生孢子數量主要與蟲體附著的孢子數量有關,故蟲生真菌致病力的強弱和相應寄主體壁上附著分生孢子的數量有關(Altre et al.,1999)。有關研究還證明寄主的體表結構影響著其體壁附著分生孢子的能力(王海川和尤民生,1999;李國霞和杜家緯,2000;王宏民等,2009)。因此,本實驗采用血球計數板計數法測量絲光綠蠅成蟲、幼蟲和蛹供試蟲體體表上附著的蠟蚧輪枝菌分生孢子量,并利用掃描電子顯微鏡等顯微技術來觀察供試蟲體體壁外長物分布情況以及分生孢子附著部位的結構形態,來討論絲光綠蠅不同蟲態的體表結構差異和各自附著蠟蚧輪枝菌分生孢子能力的關系,進而為闡述蟲生真菌對同一昆蟲不同蟲態間的致病差異性的機理提供一定參考。
1 材料與方法
1.1 供試材料
2012年6月,將盛有新鮮豬肝的罐頭瓶子掛置在室外樹梢上,讓絲光綠蠅產卵于其中。為避免在產卵時間段11∶00-21∶00(余小輝,2010)驚擾蟲體,故于每天上午11∶00 以前觀察產卵情況。數天后將產有蟲卵的罐頭瓶置于人工養蟲室中,生長環境溫度25°C,RH 為70%-80%,光周期12L∶12D。待蟲體種群數量擴大后,分別挑取鮮活的絲光綠蠅三齡初期幼蟲、發育階段Ⅶ期的蛹(王江峰等,2000)和羽化后4 d 的成蟲作為實驗供試蟲體。2012年8月,取云南農業大學植保學院農藥系實驗室保藏的蠟蚧輪枝菌KMZW-1菌株作為供試菌源。該菌種用絲光綠蠅成蟲連續復壯2-3 次后接種至PDA 培養基中純化、擴繁。7 d 后采收蠟蚧輪枝菌(于蓉蓉等,2004),parafilm 封口膜密封培養皿后再置于冰箱中4°C 儲藏備用。
1.2 絲光綠蠅三種蟲態附著蠟蚧輪枝菌分生孢子量的測定
參照彭國良等(2009)的方法配制好1×107個/mL的蠟蚧輪枝菌分生孢子懸浮液,在隨機條件下選取絲光綠蠅幼蟲、蛹、成蟲供試蟲體,用0.65%昆蟲生理鹽水清洗供試蟲體體壁雜質,待體壁干燥后用小型噴霧器將10 mL 分生孢子懸浮液均勻地噴向置于中等規格蠅籠(30 cm×30 cm×25 cm)中的供試蟲體表面。供試蟲體表面水漬全干后,用柴春煊(1984)介紹的乙醚冷凍麻醉蠅的實驗方法處理供試蟲體。稍后,參考方羽生等(2004)的方法,將被麻醉的10 頭供試蟲體置于盛有5 mL 70%乙醇的燒杯中,再利用超聲波震蕩清洗3-4 min。稍后,用0.1%Tween-80 溶液沖洗蟲體表面并收集洗脫液于燒杯中。待燒杯中的乙醇揮發一定程度后測量剩余液體積V,再移液400 μL 剩液至血球計數板中,顯微觀察并記錄孢子個數N。重復試驗5 次,測量供試蟲體的Vi和Ni,按公式ni=Ni* Vi* 106/800(1≤i≤5)計算供試蟲體所附孢子數。
1.3 絲光綠蠅供試蟲體體表的顯微觀察
1.3.1 制樣:取得被分生孢子懸浮液噴霧處理過的供試蟲體樣品后,用3.5% 戊二醛4℃下固定2 d,再用PBS 液洗滌樣品3 遍(每次約30 min),1%鋨酸后固定2 h,接著繼續用PBS 液洗滌。之后,按30%、50%、70%、80%、90%、95%、100%的酒精濃度梯度逐級脫水,每次時間約20 min。再后將樣品放入注滿叔丁醇(AR)的干燥籃內進行約2 h 滲透,再將干燥籃放入-20℃冷凍。最后,將脫水過的樣品置于ES-2030 型冷凍干燥儀中進行干燥處理;干燥后的樣品在粘臺后用E-1010 型離子濺射儀進行噴鍍。
1.3.2 絲光綠蠅三蟲態體表結構顯微觀察:先使用PXS-E2040 型體式顯微鏡、XPC-3 型雙目生物顯微鏡等光學顯微觀察技術了解絲光綠蠅各蟲態體表附著分生孢子的主要結構位點;使用Hitachi S-3400N 型掃描電子顯微鏡拍攝絲光綠蠅不同蟲態體壁結構的顯微形態,并在系統軟件SEM Data Manager(SDM)界面中觀察并測量蟲體不同部位體壁外長物分布和密度。
1.4 統計學分析
參照唐啟義和馮明光(2002)數據統計分析方法,將實驗原始數據輸入DPS v6.55 版軟件數據表,執行數據特征描述處理和單因素方差分析(One-Way ANOVA)和鄧肯氏(Duncan's)新復極差多重比較測驗,并用Microsoft Excel 2003 作統計圖表(Interactive Graph and Table);掃描電鏡圖片用Photoshop cs5 軟件進行標注、測距等處理。
2 結果與分析
2.1 絲光綠蠅三種蟲態附著蠟蚧輪枝菌分生孢子數的比較結果
絲光綠蠅成蟲附著的蠟蚧輪枝菌分生孢子初始數量最多,均值為3.38×104個孢子/頭,蛹為1.32×104個孢子/頭,幼蟲所附分生孢子均值數最小,0.89×104個孢子/頭。結果表明,絲光綠蠅初始附著的分生孢子數與其所處蟲態相關,三種蟲態附著的分生孢子數存在顯著差異性(F=63.0750,df=2,P=10-4)。由圖1 可以看出,在0.01 水平上蛹和幼蟲所附孢子數量無極顯著差異,而兩者同與成蟲存在極顯著差異。
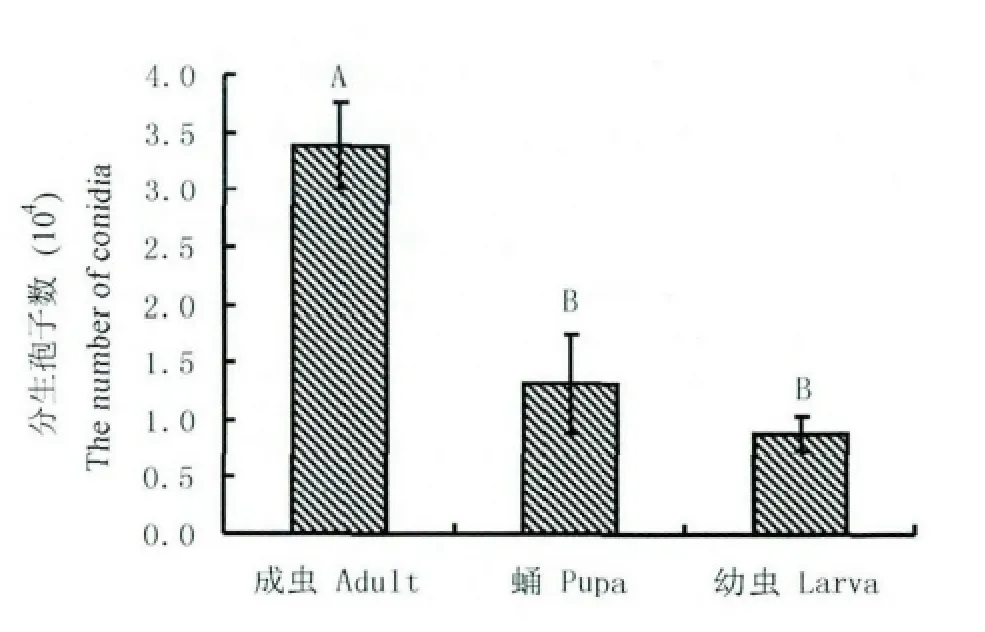
圖1 絲光綠蠅三種蟲態所附分生孢子數量Fig.1 The number of Verticilliu lecanii conidia adhered to surface of Lucilia sericate at different developmental stages
2.2 絲光綠蠅三蟲態體表結構顯微觀察
2.2.1 成蟲胸、腹部體壁外長物的形態及分布情況
背向觀察成蟲胸部的中胸盾片以及小盾片等處體壁外長物形態與分布:第2 對前中鬃長度達到第1 對后中鬃的基部,背中鬃區分布有短促且稀疏的小毛,小盾片處著生的小毛卻較為密集,腋瓣表面上的纖毛分布更為密集(圖版Ⅰ:1)。胸部的微毛以微毛a 為主,其間還夾雜有少量其它微毛,整體上其分布呈星羅狀(圖版Ⅰ:3),形體較為短粗,端部彎曲呈鉤狀,基部著生的表皮呈“山丘”隆起,且分生孢子粘液層多能牢固地粘附在這類微毛的基部與其著生的表皮位點上(圖版Ⅰ:5,7)。
尾向觀察成蟲腹部的生殖節以及生殖后節等處體壁外長物形態及分布情況:第二、三腹節的兩側著生有稀少側鬃,第三、四腹節近節間處著生有單行緣鬃,此外成蟲末腹節上著生有較前兩者密集的心鬃;腹部剛毛和鬃整體分布較為均勻,剛毛的端部、基部以及毛窩與鬃在形態上相似,且兩者在形態大小上也很接近(圖版Ⅰ:2)。腹部的微毛為微毛b,整體分布較為茂密卻幾乎無重疊(圖版Ⅰ:4),其形體較為細直,微毛端部上著生有數個凸起的“小泡”,能與其上的少量孢子殘液穩固粘附(圖版Ⅰ:6,8)。
總的看來,成蟲胸、腹部的剛毛、鬃和小毛等多是一些具有縱切向均勻“溝紋”的單細胞體壁外長物(圖版Ⅰ:3,4),溝紋以及毛窩等處均能附著孢子,而成蟲附著分生孢子主要位點在胸、腹部密集著生的微毛周圍。這些體壁外長物在成蟲胸、腹部的具體分布情況可見下表1。

表1 絲光綠蠅成蟲不同部位所著生的體壁外長物種類及密度Table 1 The diversities and densities of outgrowth in different parts of Lucilia sericate adult
2.2.2 蛹和幼蟲的體表結構及形態觀察
蛹與幼蟲體壁上著生的外長物主要為多細胞棘刺,這類體壁外長物多分布于蛹、幼蟲體節上,呈“環帶狀”間隔分布(圖版Ⅱ:1,2),且每條棘刺帶所占同體節的表面積不超過20%。觀察比較蛹和幼蟲體節上棘刺的形態結構發現同序數體節上的蛹棘刺帶的寬度和列數相比幼蟲,更寬和更多。蛹體節上的棘刺尖銳突起形似“槐刺”,其基部周圍凹陷多坑,棘刺列著生處出現骨板斷層并留下明顯的斜縫,并命名其為棘刺A(圖版Ⅱ:6);幼蟲體節上著生的棘刺形體呈“圓錐”狀,其基部越顯膨脹,棘刺叢周圍表皮雖有凹陷,但較為淺顯,命名其為棘刺B(圖版Ⅱ:7)。同樣觀察比較兩者體節上非棘刺帶部位:蛹在體節上非棘刺帶區密布有“簸箕狀”的花紋縫隙,縫寬變化范圍大,為1-14 μm,而幼蟲體節上非棘刺帶區除布有少量刻紋外整體上比較光滑(圖版Ⅱ:5,8)。
除體節外,蛹與幼蟲體表上著生體壁外長物的地方還包括蛹尾端部以及幼蟲氣門板等部位。觀察蛹和幼蟲上述部位的體壁結構形態:蛹尾端著生有11個圍氣門瘤,周圍分布嵴狀突起結構,呈輪狀發散,脊間起伏大,基部相連(圖版Ⅱ:4);幼蟲氣門板上著生的兩對大、小錐狀突間有數列呈隊形排列的小棘突—棘刺C,其基部約顯膨脹,端部尖銳形似“針芒”,和其它類型棘刺相比該棘刺排列更為緊密(圖版Ⅱ:3)。

圖版Ⅰ 絲光綠蠅成蟲胸、腹部體表結構電鏡掃描PlateⅠ Surface structure of thorax and abdomen of Lucilia sericata adult observed by scanning electron microscope
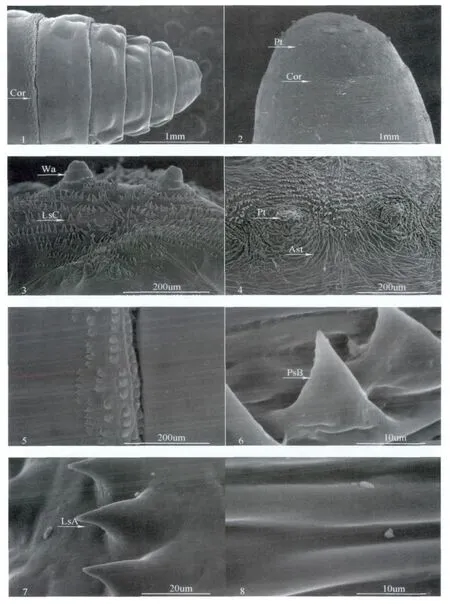
圖版Ⅱ 絲光綠蠅幼蟲、蛹體表結構電鏡掃描Plate Ⅱ Surface structure of larvae and pupa of Lucilia sericata observed by scanning electron microscope
3 結論與討論
蟲生真菌侵染的主要途徑是通過孢子萌發穿透體壁侵染蟲體,附著在昆蟲體壁上是成功侵染的第一步(周燚等,2006)。蟲生真菌對不同昆蟲的致病能力與蟲體表面附著分生孢子數量有關(Jeffs et al.,1999;李文華等,2001)。根據實驗結果,認為影響絲光綠蠅各蟲態附著蠟蚧輪枝菌分生孢子能力的因素包括:(1)蟲體附著霧滴的能力差異;(2)附著孢子的體壁結構的拓撲構型及其密度;(3)蠟蚧輪枝菌分生孢子粘附基質的類型。
噴霧處理后親水性的蠟蚧輪枝菌分生孢子隨霧流傳送至蟲體表面,附著小霧滴的能力直接影響蟲體附著孢子量(Manocha and Chen,1990;李文華等,2001)。成蟲周身布滿縱切向生有均勻“溝紋”的體壁外長物—剛毛、鬃、纖毛,而蛹與幼蟲體壁外長物主要為呈帶狀間隔分布的棘刺,但同數體節上蛹棘刺列數比縮減期前的幼蟲更多(趙博等,2009)。相比光滑的棘刺,剛毛等體壁外長物在單位體積上表面積更大,附著其上的霧滴表面張力更小,因此更容易成功附著。由此可見,絲光綠蠅各蟲態附著霧滴的能力依次為成蟲>蛹>幼蟲。
Manocha and Chen(1990)認為蟲體附著分生孢子的情況和體壁外長物的拓撲構型和密度有關。李茂業等(2011)利用掃描電鏡觀察黃綠綠僵菌Metarhizium flavoviride 侵染褐飛虱Nilaparvata lugens(St?l)時發現橢圓形的綠僵菌分生孢子多分布在褐飛虱節間膜、體表的褶皺凹陷等部位。觀察絲光綠蠅體表,發現其附著蠟蚧輪枝菌分生孢子的能力主要和刺突、微毛等這些體表結構的拓撲構型以及密度有關。成蟲體壁上附著孢子的微毛周身分布且非常密集,而蛹和幼蟲棘刺帶所占體表面積比例小且棘刺著生密度較低。此外,蛹和幼蟲兩者附著孢子的體壁結構與形態也不盡完全相同:幼蟲棘刺基部凹坑相較淺顯,該處容納分生孢子的能力較弱;蛹殼附著孢子的部位除棘刺外還包括端部的嵴狀突起結構和體節非棘刺帶,尤其后者占據了體表大部分的面積。但因其縫寬變幅大,滿足恰好容納下分生孢子以形成足夠的機械阻力來穩固附著的部位就少。
李文華等(2001)曾報道,蠟蚧輪枝菌分生孢子表面包有一層粘液外衣,能以一種“膠”粘附到多種基質上。經3.5%戊二醛等電鏡制樣液處理,觀察發現孢子粘液層在微毛a 基部形成牢固粘附,在微毛b 端部也能殘留部分粘液,而在幾丁質成分含量很高的幼蟲、蛹體壁上粘附效果卻很差,甚至未觀察到分生孢子。可見,蠟蚧輪枝菌分生孢子粘液層更易成功附著在富含蛋白質等成分的細胞外長物上,因此假設認為該型分生孢子體壁外層粘液具有選擇基質類型吸附的特性。Leger(1992)檢測萊氏野村菌Nomuraea rileyi、暗孢耳霉Conidiobolus obscurus 等多種半知菌分生孢子外壁初生粘液層,均檢測出外源凝集素;郭友中(2000)在研究蠟蚧輪枝菌對桃蚜侵染力的主要因素時推測蠟蚧輪枝菌孢子外被粘液中含有某種特異外源凝集素;Jeffs and Khachatourians(1997)檢測發現親水性輪枝菌孢子僅表面外露的多糖成分上結合了ConA(刀豆凝聚素)這一種外源凝集素。這些論據支持了蠟蚧輪枝菌孢子粘液具有選擇附著的觀點,但仍待更多的科研成果支持。
References)
Altre JA,Vandenberg JD,Cantone FA.Pathogenicity of Paecilomyces fumosoroseus isolates to diamondback moth,Plutella xylostella:correlation with spore size,germination speed,and attachment to cuticle[J].Journal of Invertcbrate Pathology,1999,73:332-338.
Chai CX.Introducing a new method to anesthetize Musca domestica,that is the method of Ether frozen anesthesia[J].Chinese Journal of Applied Entomology,1984,6(1):285-286.[柴春煊.介紹一種麻醉家蠅的新方法——乙醚冷凍麻醉法[J].昆蟲知識,1984,6(1):285-286]
Fang YS,Huang HL,Zhang JG,et al.Researching the resistance of high quality rice's breed to black kernel,and detecting the amount of its bacteria[J].Journal of Yunnan Agricultural University,2004,19(6):748-750.[方羽生,黃華林,張建國,等.水稻優質米品種對黑粒病的抗性及其種子帶菌量檢測[J].云南農業大學學報,2004,19(6):748-750]
Guo YZ.Studies on the effects of the interior and exterior factors on the infection of Verticillium lecannii against Myzus persicae[D].MSc Thesis,the Chinese Academy of Sciences,Shanghai Institute of Entomology,Shanghai,2000.[郭友中.蠟蚧輪枝菌(Verticillium lecannii)對桃蚜(Myzus persicae)侵染力的主要影響因素研究[D].上海:中科院上海昆蟲研究所碩士學位論文,2000]
Jeffs LB,Khachatourians GG.Estimation of spore hydrophobicity for members of the genera Beauveria,Metarhizium and Tolypocladium by salt-mediated aggregation and sedimentation[J].Canadian Journal of Microbiology,1997,43:23-28.
Jeffs LB,Xavier IJ,Matai RE,et al.Relationships between fungal spore morphologies and surface properties for entomopathogenic members of the genera Beauveria,Paecilomyces,Tolypocldium and Verticillium[J].Journal of Microbiology,1999,45:936-948.
Leger RJSt,Cooper RM,Charnley A.Distribution of chymoelastases and trypsin-like enzymes in 5 species of entomopathogenic[J].Archives of Biochemistry and Biophysics,1987,258:123-131.
Li GX,Du JW.Pathogenicity and reproduction effect of four isolates of Verticillium lecanii on the aphid,Myzus persicae[J].Entomological Journal of East China,2000,9(1):38-44.[李國霞,杜家緯.四株蠟蚧輪枝菌對桃蚜的侵染力及對其生殖力的影響[J].華東昆蟲學報,2000,9(1):38-44]
Li MY,Lin HF,Jin L,et al.Observations on infection of Nilaparvata lugens by Metarhizium flavoviride using a scanning electron microscope[J].Chinese Journal of Applied Entomology,2011,48(5):1412-1416.[李茂業,林華峰,金立,等.黃綠綠僵菌對褐飛虱侵染過程的掃描電鏡觀察[J].應用昆蟲學報,2011,48(5):1412-1416]
Li WH,Zhang YJ,Wang ZK.The study about the physical and chemical factors of Inhomogeneous fungus penetrates the insect cuticle.In:Li DM,eds.Insects and the Environment,Proceedings of the 2001 Academic Year of the Entomological Society of China[C].Beijing:China Agriculture Press,2001:473-479.[李文華,張永軍,王中康.蟲生真菌穿透昆蟲表皮相關理化因子的研究.見:李典謨,主編.昆蟲于環境——中國昆蟲學會2001年學術年會議文集[C].北京:中國農業出版社.2001:473-479]
Manocha MS,Chen Y.Specificity of attachment of fungal parasites to their hosts[J].Journal of Microbiology,1990,36:69-76.
Peng GL,Xue JL,Liu WM,et al.The role of protease and chitinase,when Verticillium lecanii invade the epidermis of Chrysomphalus aonidum[J].Chinese Journal of Applied and Environmental Biology,2009,2:220-225.[彭國良,薛皎亮,劉衛敏,等.蠟蚧輪枝菌入侵蚧蟲表皮過程中蛋白酶和幾丁質酶的作用[J].應用與環境生物學報,2009,2:220-225]
Sharififard M,Mossadegh MS,Vazirianzadeh B,et al.Laboratory evaluation of pathogenicity of entomopathogenic fungi,Beauveria bassiana(Bals.)vuill.and Metarhizium anisopliae(Metch.)Sorok.to larvae and adults of the house fly,Musca domestica L.(Diptera:Muscidae)[J].Asian Journal of Biological Science,2011,4(2):128-137.
Tang QY,Feng MG.Practical Statistics and DPS Data Statistical Analysis System[M].Beijing:Science Press,2002,33-59.[唐啟義,馮明光.實用統計分析及其DPS 數據統計分析系統[M].北京:科學出版社,2002,33-59]
Wang HC,You MS.Advances in conidia attachment by Metarhizium anisopliae on the host[J].Entomological Knowledge,1999,36(3):189-192.[王海川,尤民生.綠僵菌分生孢子對寄主附著機制的研究進展[J].昆蟲知識,1999,36(3):189-192]
Wang HM,Zhang H,Hao C,et al.Infection and pathogenicity of Paecilomyces fumosoroseus on Plutella xylostella[J].Chinese Journal of Eco-Agriculture,2009,17(4):704-708.[王宏民,張奐,郝赤,等.玫煙色擬青霉對小菜蛾幼蟲的侵染過程及接菌方法對其致病力的影響[J].中國生態農業學報,2009,17(4):704-708]
Wang LC,Gao X,Wang ST,et al.Doing research on the biologicalcharacteristics and pathogenicity of Verticillium lecanii,suffering the diseased Bioettcherisca peregrina[J].Journal of Anhui Agricultural Sciences,2010,38(15):7909-7912.[王凌超,高熹,王松田等.罹病棕尾別麻蠅體上蠟蚧輪枝菌的生物學特性與致病性研究[J].安徽農業科學,2010,38(15):7909-7912]
Wang JF,Cheng YC,Hu C,et al.Utilizing the morphology of Lucilia sericata pupal development to make a judgment for the time of death[J].Journal of Sun Yatsen University,2000,39(6):250-254.[王江峰,陳玉川,胡萃,等.絲光綠蠅蛹發育形態學用于死亡時間判斷的研究[J].中山大學學報,2000,39(6):250-254]
Yu RR,Shi WP,Yan YY,et al.The harvest time of Verticillium lecanii conidial to affect its germination and virulence[J].Acta Phytophylacica Sinica,2004,30(5):41-43.[于蓉蓉,石旺鵬,閆躍英,等.采收時間對蠟蚧輪枝菌分生孢子的萌發及致病力的影響[J].植物保護,2004,30(5):41-43]
Yu XH.The overview of Lucilia sericata research[J].Chinese Journal of Vector Biology and Control,2010,21(4):402-404.[余小輝.絲光綠蠅研究慨況[J].中國媒介生物及控制雜志,2010,21(4):402-404]
Zhao B,Wang L,Wang H,et al.Morphological changes of Lucilia sericata larvae and its significance in forensic medicine[J].Chinese Journal of Vector Biology and Control,2009,20(6):534-537.[趙博,王玲,王賀等.絲光綠蠅幼蟲不同溫度下的形態學變化及法醫學意義[J].中國媒介生物學及控制雜志,2009,20(6):534-537]
Zhou Y,Wang ZK,Yu Z.Research and Application of Microbial Pesticides[M].Beijing:Chemical Industry Press,2006,105-116.[周燚,王中康,喻子.微生物農藥研發與應用[M].北京:化學工業出版社,2006,105-116]
