過表達NF-E2相關因子2對白癜風黑素細胞PIG3V線粒體生物合成和功能的影響
2015-12-01 01:09:22朱龍飛田軍堅哲劉邦民肖茜李春英高天文
中華皮膚科雜志 2015年6期
朱龍飛 田軍 堅哲 劉邦民 肖茜 李春英 高天文
過表達NF-E2相關因子2對白癜風黑素細胞PIG3V線粒體生物合成和功能的影響
朱龍飛 田軍 堅哲 劉邦民 肖茜 李春英 高天文
目的 探討過表達NF-E2相關因子2(Nrf2)對黑素細胞線粒體合成和功能的影響。方法 構建含有人Nrf2基因全長的過表達質粒Nrf2-pEX-1,用該質粒瞬時轉染白癜風患者表皮黑素細胞系(PIG3V)。實驗分為空白組(不含質粒)、對照組(轉染pEX-1空質粒)、過表達組(轉染pEX-1-Nrf2過表達質粒)。過表達Nrf2后,RT-PCR和Western印跡法檢測PIG3V線粒體生物合成相關的Nrf2、核呼吸因子1(NRF1)、線粒體轉錄因子A(TFAM)mRNA和蛋白水平的變化;RT-PCR檢測線粒體DNA(mtDNA)拷貝數。流式細胞儀檢測線粒體膜電位(MMP);熒光素酶報告系統檢測細胞ATP含量。采用兩樣本t檢驗進行統計學分析。結果 Nrf2過表達質粒轉染PIG3V細胞后24 h,Nrf2 mRNA、NRF1 mRNA水平較對照組明顯升高,差異有統計學意義(均P<0.001),而TFAM mRNA水平與對照組比較差異無統計學意義;48 h后Nrf2 mRNA、TFAM mRNA水平較對照組升高,差異均有統計學意義(P<0.05),而NRF1 mRNA水平與對照組差異無統計學意義。Western印跡法顯示,轉染后24 h,過表達組Nrf2、NRF1、TFAM蛋白表達與對照組比較差異均無統計學意義,48 h后過表達組Nrf2、NRF1、TFAM蛋白表達升高(P<0.05)。過表達組MMP在轉染24 h后較對照組升高2.313%(t=5.546,P=0.005),48 h后升高14.872%(t=8.537,P=0.001)。線粒體合成指標相對mtDNA拷貝數,在轉染24 h后兩組間差異無統計學意義(P>0.05),48 h后過表達組明顯高于對照組(t=5.760,P=0.005);細胞ATP含量在轉染后24 h兩組間差異無統計學意義(P>0.05),48 h后過表達組ATP含量較對照組明顯升高(t=22.040,P=0.008)。結論 過表達Nrf2可以促進黑素細胞線粒體生物合成相關基因和蛋白的表達,進而促進線粒體生物合成,上調線粒體功能相關的指標。
白癜風;黑素細胞;線粒體;NF-E2相關因子2;腺苷三磷酸
白癜風是一種獲得性色素脫失性疾病,其發病機制不明,黑素細胞氧化應激損傷在白癜風的發生、發展中起關鍵作用[1]。線粒體是細胞內重要的能源工廠和活性氧簇(reactive oxygen species,ROS)來源,也是氧化應激損傷首當其沖的攻擊靶點[2]。白癜風患者皮損周圍黑素細胞、角質形成細胞和朗格漢斯細胞以及外周血單一核細胞中線粒體形態及功能均發生異常[3]。臨床研究發現,線粒體刺激劑聯合抗氧化劑可有效改善穩定期白癜風患者癥狀,使皮損區色素得到恢復[4]。核因子 E2相關因子 2(nuclear factor E2-related factor 2,Nrf2)通路是細胞內重要的抗氧化通路,可促進下游多種抗氧化因子的表達,發揮抗氧化作用。在過表達Nrf2后線粒體生物合成增加,功能改善,小鼠肝臟細胞壞死、凋亡減少[5]。Nrf2在肝臟、心肌細胞中促進線粒體生物合成、改善線粒體功能的作用,在白癜風黑素細胞是否存在[5-6]。本研究采用過表達質粒轉染技術上調白癜風患者表皮黑素細胞系(PIG3V)Nrf2的表達,探討對線粒體生物合成和功能的影響。
資料與方法
一、資料與試劑
1.細胞來源:白癜風患者表皮黑素細胞系PIG3V,由美國伊利諾斯州芝加哥Loyola大學Caroline Le Poole博士贈送。該細胞系運用反轉錄病毒載體轉染16型人乳頭狀瘤病毒E6和E7開放讀碼框得來,與正常黑素細胞生物學特性相似。
2.主要儀器和試劑:254培養基、HMGS、脂質體Lipofectamine 2000為美國Gibco公司產品。Nrf2基因全長過表達質粒由上海吉瑪制藥技術有限公司設計、合成,基因長度1 770 bp,克隆載體為pEX-1,克隆位點為XhoI/BamHI;Trizol試劑、反轉錄cDNA試劑盒、SYBR Premix Ex Taq II試劑盒為大連TaKaRa公司產品;Nrf2、核呼吸因子 1(NRF1)、線粒體轉錄因子A(TFAM)、ND4引物由上海生工生物工程股份有限公司合成。Nrf2、NRF1、TFAM兔抗人單克隆抗體為美國Abcom公司產品。抗β肌動蛋白鼠單克隆抗體、辣根過氧化物(HRP)標記羊抗兔(羊抗鼠)IgG產自北京康為世紀生物科技有限公司。MitoTracker?Red CMXRos為美國Invitrogen公司產品。Enliten?ATP檢測試劑盒產自美國Promego公司。Bio-rad IQ5實時熒光定量PCR儀。BD FACSAria流式細胞分選儀。美國promega公司E5311熒光素酶報告系統。
二、方法
1.Nrf2基因全長過表達質粒瞬時轉染PIG3V細胞體系的建立:轉染需用無抗生素無血清的培養基,所用槍頭、微量離心管均需用DEPC水做無核酶、無DNA處理。瞬時轉染同時在6孔板和25 cm2培養瓶中進行,以便用于線粒體合成相關基因和蛋白表達、mtDNA拷貝數、線粒體膜電位(mitochondrial membrane potential,MMP)、細胞 ATP含量等檢測。實驗分為空白組(不含質粒)、對照組(轉染pEX-1空質粒)、過表達組(轉染pEX-1-Nrf2過表達質粒)。6 孔板每孔質粒 4 μg,脂質體 12 μl;25 cm2培養瓶每瓶質粒 12 μg,脂質體 30 μl。按照Lipofectamine 2000轉染試劑盒說明書進行瞬時轉染,轉染后無血清254培養基37℃培養,6 h后換液,24 h觀察熒光。本質粒帶綠色熒光,成功轉染的細胞在藍光激發下胞質顯示顆粒狀綠色熒光。
2.實時熒光定量PCR檢測與線粒體合成相關的 Nrf2、NRF1、TFAM mRNA 水平:提取處理后PIG3V細胞的mRNA,定量,反轉錄成cDNA。PCR引物序列:Nrf2 正向:5′-CTTGGCCTCAGTGATTCTG AAGTG-3′,反向:5′-CCTGAGATGGTGACAAGGGT TGTA-3′;NRF1 正向:5′-CCACGTTACAGGGAGGT GAG-3′,反向:5′-TGTAGCTCCCTGCTGCATCT-3′;TFAM 正向:5′-ACAGGATGATGACTATGGAA-3′,反向:5′-CAACTCTGAATACAATGTGAATT-3′;β 肌動蛋白正向 5′-CGGGAAATCGTGCGTGACAT-3′,反向:5′-GAAGGAAGGcTGGAAG AGTG-3′。PCR 擴增體系包括:cDNA模板2μl,引物正向和反向各1μl,SYBR Premix Ex Taq II 12.5 μl,ddH2O 8.5 μl。擴增條件:預變性 95 ℃ 30 s;95 ℃ 5 s,55 ℃ 30 s,共 40 個循環,95℃ 1 min,65 ℃ 30 s,72 ℃ 15 s,1 個循環。在實時熒光定量PCR儀上讀取Ct值。以β肌動蛋白作為內參照。采用2-ΔΔCt法計算對特異性擴增的目的基因進行相對定量。
3.Wesrern印跡法檢測線粒體合成相關蛋白的表達:提取處理后PIG3V細胞總蛋白。蛋白定量,煮沸變性,電泳并用半干轉法轉膜,封閉,分別加入兔抗人 Nrf2、NRF1、TFAM 抗體,4 ℃搖床上過夜,洗膜,加二抗,室溫2 h,洗膜,曝光顯影。以β肌動蛋白作為內參照對所得蛋白條帶圖進行灰度分析。
4.線粒體生物合成檢測:通過檢測細胞全基因組中mtDNA拷貝數來評價線粒體生物合成水平,選擇線粒體DNA特征性基因ND4作為檢測的目標基因,設計引物。PCR引物序列:ND4基因正向:5′-CCATTCTCCTCCTATCCCTCAAC-3′,反向:5′-CAC AATCTGATGTTTTGGTTAAACTATATTT-3′。提取處理后細胞總DNA,定量。PCR擴增體系包括:DNA模板 0.36 μg(體積不同),10 μmol/L ND4引物 F 端和 R 端各 1 μl,SYBR Ⅱ Premix Ex TaqTM12.5 μl,加ddH2O適量補足反應體系至25 μl。擴增條件、結果分析同上文。相對mtDNA拷貝數=ND4基因DNA拷貝數/β肌動蛋白基因拷貝數。
5.MMP檢測:常規消化細胞,生理氯化鈉液洗1遍,將MitoTracker?Red CMXRos按1∶5 000用無血清254培養基稀釋,用稀釋液重懸細胞,37℃孵育30 min,每10分鐘振蕩1次,其后生理氯化鈉液洗3遍,用適量生理氯化鈉液重懸,上流式細胞儀,檢測PE通道的平均熒光強度值。MMP與平均熒光強度值正相關。
6.線粒體ATP產量檢測:將各組細胞常規消化,用254培養基重懸,使用細胞計數板計數,每組各取4 500個細胞進行下一步實驗。向細胞中加入2%三氯乙酸50 μl/管,室溫靜置10 min后向管中加入50 μl 4%三羥甲基氨基甲烷中和,1 000×g離心3 min,取上清液。使用Enliten?ATP檢測試劑盒,用熒光素酶報告系統檢測RLU值。用試劑盒中ATP標準品設立ATP濃度梯度RLU值,建立RLU值-ATP濃度的標準曲線,按照擬合公式計算各樣品ATP含量。
7.統計學方法:用graphpad prism 5軟件進行數據分析。mRNA表達水平比較、mtDNA拷貝數、MMP、ATP含量比較采用兩樣本t檢驗進行統計學分析,P<0.05為差異有統計學意義。
結 果
一、Nrf2基因全長過表達質粒瞬時轉染PIG3V細胞體系的建立
轉染后24 h在倒置熒光顯微鏡下觀察,對比白光和藍光激發后圖像(圖1)。轉染后的細胞胞質內呈現顆粒狀綠色熒光,胞核部位不顯示熒光,提示質粒轉染成功。
二、Nrf2過表達對線粒體合成相關基因mRNA和蛋白水平的影響
Nrf2過表達質粒轉染PIG3V細胞后24 h,過表達組Nrf2 mRNA、NRF1 mRNA水平均較對照組明顯升高(均P<0.001),而TFAM mRNA水平較對照組差異無統計學意義(P>0.05)。48 h后Nrf2 mRNA、TFAM mRNA水平較對照組升高(均P<0.05),而NRF1 mRNA水平較對照組差異無統計學意義(P>0.05),見表1。Western印跡法結果顯示(圖 2),轉染后 24 h 時,過表達組 Nrf2、NRF1、TFAM蛋白表達與對照組相比差異均無統計學意義(P>0.05);48 h 時,過表達組 Nrf2、NRF1、TFAM 蛋白均較對照組明顯升高(P<0.05)。見表2。
三、Nrf2過表達后細胞相對mtDNA拷貝數變化
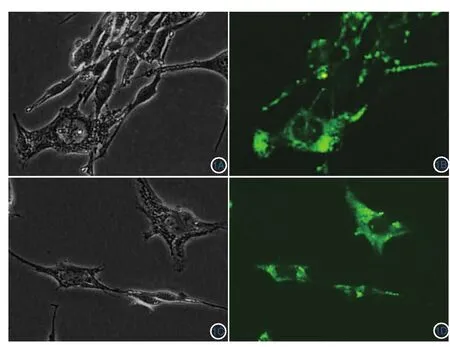
圖1 轉染質粒后細胞熒光圖(×400):本質粒帶綠色熒光,成功轉染的細胞在藍光激發下,胞質呈現綠色顆粒狀熒光,胞核無熒光 1A:pEX-1空質粒組白光下圖像;1B:pEX-1空質粒組藍光激發下圖像;1C:轉染pEX-1-Nrf2過表達質粒組白光下圖像;1D:轉染pEX-1-Nrf2過表達質粒組藍光下圖像
轉染后24 h,過表達組相對mtDNA拷貝數(0.247±0.172)與對照組(0.515±0.422)相比差異無統計學意義(t=0.820,P=0.473);轉染后 48 h,過表達組(8.167±1.620)高于對照組(2.779±0.013),差異有統計學意義(t=5.760,P=0.005)。
表1 黑素細胞過表達Nrf2后相關基因mRNA相對表達量(2-ΔΔCt,±s)
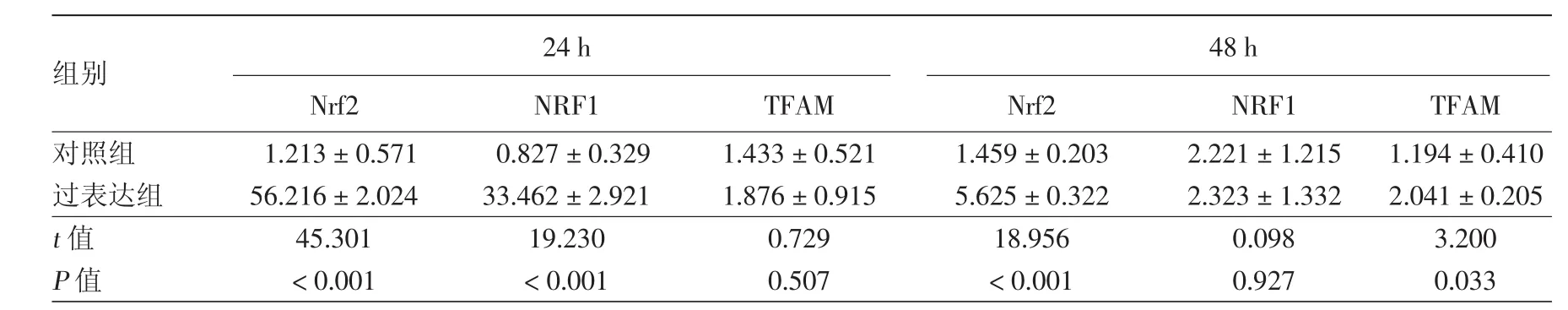
表1 黑素細胞過表達Nrf2后相關基因mRNA相對表達量(2-ΔΔCt,±s)
注:n=4。Nrf2:核因子E2相關因子2;NRF1:核呼吸因子1;TFAM:線粒體轉錄因子A
組別 24 h 48 h Nrf2 NRF1 TFAM Nrf2 NRF1 TFAM對照組 1.213±0.571 0.827±0.329 1.433±0.521 1.459±0.203 2.221±1.215 1.194±0.410過表達組 56.216±2.024 33.462±2.921 1.876±0.915 5.625±0.322 2.323±1.332 2.041±0.205 t值 45.301 19.230 0.729 18.956 0.098 3.200 P值 <0.001 <0.001 0.507 <0.001 0.927 0.033
四、Nrf2過表達后MMP的變化
Nrf2過表達后24 h,過表達組MMP平均熒光強度值(73.974±0.44)較對照組(72.302±0.28)升高2.313%,差異有統計學意義(t=5.546,P=0.005)。48 h后過表達組 MMP(16.537± 0.38)較對照組(14.396±0.21)升高14.872%,差異有統計學意義(t=8.537,P=0.001)。
五、Nrf2過表達后線粒體ATP產量的變化
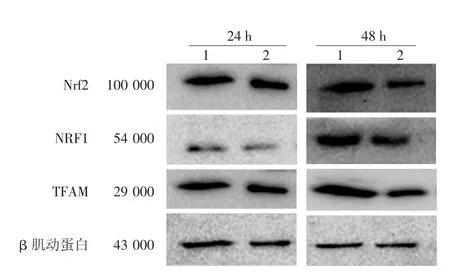
圖2 過表達Nrf2后線粒體合成相關蛋白表達水平的變化1:過表達組;2:對照組。Nrf2:核因子E2相關因子2;NRF1:核呼吸因子1;TFAM:線粒體轉錄因子A
轉染后24 h,Nrf2過表達組ATP含量(1.593±2.122)×10-13mol/細胞與對照組(1.111±0.902)×10-13mol/細胞相比差異無統計學意義(t=0.418,P=0.691);轉染后 48 h,Nrf2過表達組 ATP含量(12.820±0.561)×10-13mol/細胞較對照組(7.437±0.556)× 10-13mol/細胞明顯升高 (t=22.040,P=0.008)。
討 論
穩定期白癜風患者白斑區皮膚內角質形成細胞線粒體腫脹,線粒體嵴崩裂[7]。進展期白癜風患者白斑周邊皮膚內黑素細胞線粒體腫脹,線粒體嵴重排;白斑周邊皮膚內角質形成細胞中的大部分線粒體內膜、膜間隙和基質腫脹,線粒體嵴重排,而正常對照皮膚相應細胞中線粒體結構正常[3]。線粒體損傷誘導的線粒體凋亡途徑是細胞凋亡主要途徑之一。因為線粒體損傷后MMP下降,ATP產生減少,膜通透性轉換小孔開放,細胞色素C(Cyt-C)釋放至胞質,形成Cyt-C/Apaf-1/procaspase-3凋亡復合體,引起procaspase-3激活成凋亡蛋白Caspase-3,進而引起細胞的凋亡[8]。我們前期研究發現,黃芩素可以抑制線粒體Cyt-C釋放,從而減少氧化應激誘導的人黑素細胞凋亡,提示改善線粒體功能可提高黑素細胞的抗氧化應激能力[9]。
表2 黑素細胞過表達Nrf2后相關蛋白灰度分析(±s)
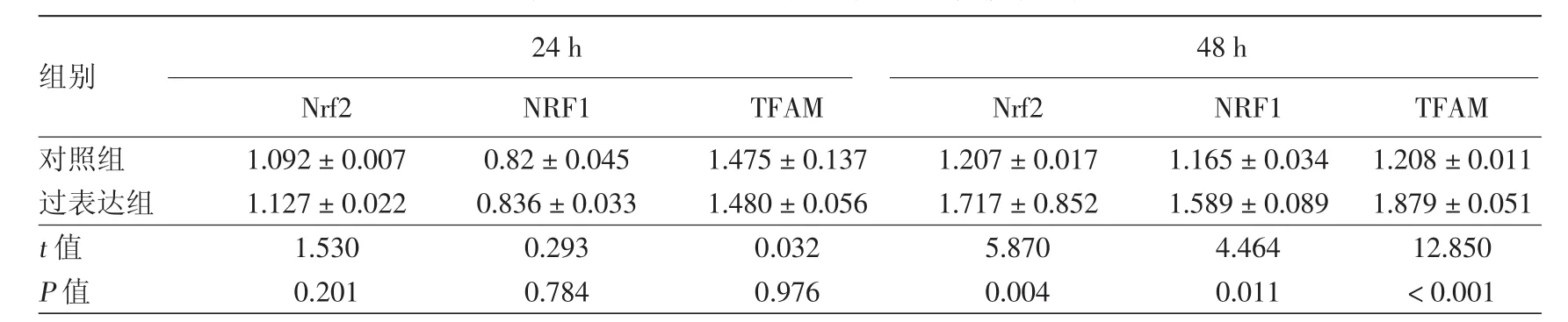
表2 黑素細胞過表達Nrf2后相關蛋白灰度分析(±s)
注:n=4。Nrf2:核因子E2相關因子2;NRF1:核呼吸因子1;TFAM:線粒體轉錄因子A
組別 24 h 48 h Nrf2 NRF1 TFAM Nrf2 NRF1 TFAM對照組 1.092±0.007 0.82±0.045 1.475±0.137 1.207±0.017 1.165±0.034 1.208±0.011過表達組 1.127±0.022 0.836±0.033 1.480±0.056 1.717±0.852 1.589±0.089 1.879±0.051 t值 1.530 0.293 0.032 5.870 4.464 12.850 P值 0.201 0.784 0.976 0.004 0.011 <0.001
研究發現,Nrf2可直接調節小鼠神經元和成纖維細胞線粒體呼吸作用,上調Nrf2后MMP升高,ATP生成增加,氧化磷酸化效率提高[10]。本研究在白癜風患者黑素細胞中觀察到,過表達Nrf2后,MMP升高,細胞內ATP含量增加,與文獻報道結果類似[10]。在心肌細胞中,Nrf2入核與抗氧化反應元件(antioxidant response element,ARE)啟動子區域結合,使NRF1上調,上調的NRF1促進TFAM表達,TFAM使線粒體合成增加[6]。本研究發現,在轉染Nrf2過表達質粒后24 h,NRF1 mRNA即明顯升高,而TFAM mRNA無明顯變化;在48 h后NRF1 mRNA降至正常水平時,TFAM mRNA卻明顯升高。TFAM mRNA升高滯后于NRF1 mRNA升高的現象,提示NRF1是作為上游基因調控TFAM的升高,與小鼠肝臟細胞、心肌細胞內的研究結果一致[5-6]。
Nrf2通路是細胞內抗氧化應激的主要通路。Nrf2在合成后與Keap1結合而處于無功能狀態,在ROS的作用下其與Keap1解離,進而被磷酸化而激活。磷酸化的Nrf2發生核轉位,與ARE相結合,促進一系列的抗氧化基因的表達,NRF1就是其中之一。NRF1上調后進一步促進TFAM表達,進而使線粒體生物合成增加,功能障礙改善。另外,Nrf2基因可被 PI3K、PKC、JNK、ERK、P38 等信號分子激活[11-12]。我們前期研究發現,過表達Nrf2可對氧化應激作用下人黑素細胞發揮保護作用,并且我們證實血紅素單加氧酶-1(heme oxygenase,HO-1)等抗氧化因子的上調是其發揮保護作用的機制[12-13]。作為細胞內作用廣泛的轉錄因子,Nrf2保護黑素細胞的機制可能不僅限于此。通過本實驗我們證實,在白癜風黑素細胞中Nrf2改善線粒體功能,間接抑制線粒體凋亡途徑,可能是其抗氧化應激作用的另一重要機制。
[1]Denat L,Kadekaro AL,Marrot L,et al.Melanocytes as instigators and victims of oxidative stress[J].J Invest Dermatol,2014,134(6):1512-1518.
[2]Murphy MP.How mitochondria produce reactive oxygen species[J].Biochem J,2009,417(1):1-13.
[3]Prignano F,Pescitelli L,Becatti M,et al.Ultrastructural and functional alterations of mitochondria in perilesional vitiligo skin[J].J Dermatol Sci,2009,54(3):157-167.
[4]Rojas-Urdaneta JE,Poleo-Romero AG.Evaluation of an antioxidant and mitochondria-stimulating cream formula on the skin of patients with stable common vitiligo[J].Invest Clin,2007,48(1):21-31.
[5]MacGarvey NC,Suliman HB,Bartz RR,et al.Activation of mitochondrial biogenesis by heme oxygenase-1-mediated NF-E2-related factor-2 induction rescues mice from lethalStaphylococcus aureussepsis[J].Am J Respir Crit Care Med,2012,185(8):851-861.
[6]Piantadosi CA,Carraway MS,Babiker A,et al.Heme oxygenase-1 regulates cardiac mitochondrial biogenesis via Nrf2-mediated transcriptional control of nuclear respiratory factor-1[J].Circ Res,2008,103(11):1232-1240.
[7]Panuncio AL,Vignale R.Ultrastructural studies in stable vitiligo[J].Am J Dermatopathol,2003,25(1):16-20.
[8]Stevens JM.Cytochrome c as an experimental model protein[J].Metallomics,2011,3(4):319-322.
[9]Liu B,Jian Z,Li Q,et al.Baicalein protects human melanocytes from H2O2-induced apoptosis via inhibiting mitochondriadependent caspase activation and the p38 MAPK pathway[J].Free Radic Biol Med,2012,53(2):183-193.
[10]Holmstr?m KM,Baird L,Zhang Y,et al.Nrf2 impacts cellular bioenergetics by controlling substrate availability for mitochondrial respiration[J].Biol Open,2013,2(8):761-770.
[11]No JH,Kim YB,Song YS.Targeting nrf2 signaling to combat chemoresistance[J].J Cancer Prev,2014,19(2):111-117.
[12]Jian Z,Li K,Song P,et al.Impaired activation of the Nrf2-ARE signaling pathway underminesH2O2-induced oxidative stress response:a possible mechanism for melanocyte degeneration in vitiligo[J].J Invest Dermatol,2014,134(8):2221-2230.
[13]Jian Z,Li K,Liu L,et al.Heme oxygenase-1 protects human melanocytes from H2O2-induced oxidative stress via the Nrf2-ARE pathway[J].J Invest Dermatol,2011,131(7):1420-1427.
2014-10-15)
(本文編輯:顏艷)
Effects of up-regulation of NF-E2-related factor 2 on mitochondrial biosynthesis and function in an immortalized human vitiligo melanocyte cell line PIG3V
Zhu Longfei,Tian Jun,Jian Zhe,Liu Bangmin,Xiao Qian,Li Chunying,Gao Tianwen.Department of Dermatology,Xijing Hospital,Fourth Military Medical University,Xi′an 710032,China
Gao Tianwen,Email:gaotw@fmmu.edu.cn
ObjectiveTo explore the effects of NF-E2-related factor 2(Nrf2)overexpression on mitochondrial biosynthesis and function in melanocytes.MethodsAn immortalized human vitiligo melanocyte cell line PIG3V was used in this study.An overexpression plasmid Nrf2-pEX-1 containing the full-length Nrf2 gene was constructed.PIG3V cells were divided into 3 groups:blank group receiving no treatment,control group transfected with the pEX-1 plasmid,overexpression group transfected with the Nrf2-pEX-1 plasmid.After transfection,real-time quantitative reverse transcription-PCR (RT-PCR) and Western blot were performed to determine the mRNA and protein levels of mitochondrial biosynthesis-related factors(including Nrf2,nuclear respiratory factor 1 (NRF1)and mitochondrial transcription factor A (TFAM))respectively;RT-PCR was also conducted to measure the copy number of mitochondrial DNA(mtDNA),and flow cytometry to estimate mitochondial membrane potential(MMP);luciferase reporter system was used to estimate the intracellular adenosine triphosphate(ATP)level.Statistical analysis was carried out by using a twosamplet-test.ResultsAfter transfection,a significant increase was observed in the mRNA expression levels of Nrf2 and NRF1 at 24 hours(bothP<0.001)and in those of Nrf2 and TFAM at 48 hours(bothP<0.05),but no significant change was noted in the mRNA expression level of TFAM at 24 hours(P>0.05)or in that of NRF1 at 48 hours(P>0.05)in the overexpression group compared with the control group.In the case of Nrf2,NRF1 and TFAM protein levels,the overexpression group showed significant increases compared with the control group at 48 hours after transfection(allP< 0.05),while no significant difference was noted between the two groups at 24 hours.Compared with the control group,MMP in the overexpression group increased by 2.313%at 24 hours(t=5.546,P=0.005)and by 14.872%at 48 hours(t=8.537,P=0.001)after transfection.Both the relative copy number of mtDNA and ATP level were similar between the overexpression group and control group at 24 hours after transfection (bothPgt;0.05),but significantly higher in the overexpression group than in the control group at 48 hours (t=5.760,P=0.005;t=22.040,P=0.008).ConclusionUp-regulation of Nrf2 pathway can improve mitochondrial function and biosynthesis in PIG3V cells likely by promoting the expressions of mitochondrial biosynthesis-related genes and proteins.
Vitiligo;Melanocytes;Mitochondria;NF-E2-related factor 2;Adenosine triphosphate
10.3760/cma.j.issn.0412-4030.2015.06.002
國家自然科學基金面上項目(81373844);國家自然科學基金青年基金(81102669、81402599)
710032西安,第四軍醫大學西京皮膚醫院
高天文,Email:gaotw@fmmu.edu.cn
