嗜酸乳桿菌胞壁肽聚糖對小鼠腸黏膜的免疫調(diào)控
2015-10-29 02:13:42潘道東郭宇星曾小群孫楊贏
食品科學(xué) 2015年13期
吳 振,潘道東,,*,郭宇星,曾小群,孫楊贏
(1.南京師范大學(xué)食品科學(xué)與營養(yǎng)系,江蘇 南京 210097;2. 寧波大學(xué)海洋學(xué)院,浙江 寧波 315211)
嗜酸乳桿菌胞壁肽聚糖對小鼠腸黏膜的免疫調(diào)控
吳振1,潘道東1,2,*,郭宇星1,曾小群2,孫楊贏2
(1.南京師范大學(xué)食品科學(xué)與營養(yǎng)系,江蘇 南京 210097;2. 寧波大學(xué)海洋學(xué)院,浙江 寧波 315211)
通過建立以致病性大腸桿菌感染小鼠為模型,研究氧化修飾前后嗜酸乳桿菌肽聚糖(peptidoglycan,PG)對小鼠腸道上皮細胞病理形態(tài)影響,及對感染大腸桿菌腸黏膜分泌sIgA、血清干擾素-γ(interferon-γ,IFN-γ)和腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)的影響,發(fā)現(xiàn)灌胃PG后能夠促進小腸黏膜分泌sIgA,降低IFN-γ及TNF-α的表達,且選擇性氧化修飾PG比未修飾PG具有優(yōu)勢。可見,嗜酸乳桿菌在腸道免疫方面,可以通過其功能性PG提高對小腸黏膜的免疫調(diào)控作用。
嗜酸乳桿菌;肽聚糖;腸黏膜;免疫調(diào)控
黏膜作為機體的首要防御屏障,其主要功能是排斥和清除有害外源粒子,保護機體免受外源損傷[1]。隨著研究的深入,黏膜免疫保護機制的研究越來越受到國內(nèi)外學(xué)者的廣泛關(guān)注。在腸道中,誘導(dǎo)黏膜免疫的調(diào)控主要發(fā)生在腸道相關(guān)淋巴組織,特別是淋巴集結(jié)(peyer's patches,PP),以及在腸系膜內(nèi)存在的淋巴結(jié)[2]。有關(guān)研究表明,益生菌如乳酸桿菌、雙歧桿菌能夠通過釋放細胞因子白細胞介素-10(interleukin-10,IL-10),抑制腸道致病菌對腸道上皮細胞的黏附作用,保護腸道免受致病菌傷害[3-4]。
急性腸炎是盆腹腔腫瘤放療過程中最常見并發(fā)癥,以腹痛、腹瀉、里急后重、黏液血便為主要表現(xiàn)[5]。臨床中急性腸炎發(fā)病率高達50%~70%,不僅影響原發(fā)病的治療,而且可以引起腸道吸收不良,導(dǎo)致貧血、消瘦,嚴重者導(dǎo)致全身炎癥反應(yīng),多器官功能衰竭,甚至死亡[6-7]。“腸道內(nèi)分泌功能障礙”是指腸道內(nèi)分泌功能不足或紊亂所導(dǎo)致的機體腸組織器官功能改變。這種情況下的腸道損傷,需要補充營養(yǎng)和水及電解質(zhì)來維持機體健康成長[8]。小腸黏膜的組織結(jié)構(gòu)與其功能狀態(tài)有著密切聯(lián)系,腸黏膜形態(tài)結(jié)構(gòu)發(fā)生變化可以通過小腸絨毛高度、黏膜厚度及腸腺深度間接推測出腸道的病變情況[9]。乳酸桿菌等有益菌的代謝產(chǎn)物可以通過腸黏膜處的吞噬細胞進入PP激活Th2細胞,從而產(chǎn)生大量的IL-10,而IL-10能夠進一步促進免疫球蛋白IgA因子的分泌。在Ig產(chǎn)生過程中,腸黏膜上皮細胞分泌物可以與IgA結(jié)合形成sIgA并排列在腸道黏膜上[10-11]。因此,當sIgA與腸黏膜表面的益生菌共同存在時,就可以抑制致病菌在腸黏膜表面的黏附,進而限制了有害菌的繁殖,對腸道菌群及腸黏膜功能的正常發(fā)揮都起到了很好的保護作用[12-13]。
乳酸菌細胞壁中肽聚糖的免疫學(xué)研究,目前主要集中在雙歧桿菌細胞壁肽聚糖方面,口服或非胃腸道途徑使肽聚糖進入宿主體內(nèi),可使宿主免疫監(jiān)視功能得到提高,并且促使免疫器官體內(nèi)淋巴細胞產(chǎn)生各種細胞因子和抗體,同時提高巨噬細胞和自然殺傷細胞活性,達到提高機體特異與非特異免疫功能,從而發(fā)揮自身穩(wěn)定的細胞調(diào)節(jié)和抗感染的效果[14-15]。1-氧基-2,2,6,6,-四甲基哌啶(2,2,6,6-tetramethylpiiperidine-1-oxyl,TEMPO)是近年來發(fā)現(xiàn)的一種新型穩(wěn)定的含氧自由基,它在自旋標記法及理論上被廣泛應(yīng)用,目前已成為自由基化學(xué)主要發(fā)展方向之一[16]。TEMPO與NaClO/NaBr構(gòu)成一種新型催化氧化體系,此體系具有高度選擇性氧化功能,將糖類物質(zhì)中的伯羥基氧化成相應(yīng)的羧酸鹽,從而賦予了糖類物質(zhì)一些特殊的性能,如溶解性;而對來源于嗜酸乳桿菌肽聚糖的氧化修飾未見報道。本實驗通過建立以致病性大腸桿菌(Escherichia coli 0111:B4)感染小鼠為模型,研究選擇性氧化修飾前后嗜酸乳桿菌肽聚糖(peptidoglycan,PG)對小鼠腸道上皮細胞病理形態(tài)學(xué)及PG對感染大腸桿菌腸黏膜分泌sIgA、血清干擾素-γ(interferon-γ,IFN-γ)和腫瘤壞死因子-α(tumor necrosis factor-α,TNF-α)細胞因子的影響,探討PG對感染致病性大腸桿菌小鼠腸黏膜的免疫調(diào)節(jié)作用。
1 材料與方法
1.1動物、試劑與儀器
健康成年雄性ICR小鼠(動物合格許可證編號:批號SCXK(HU)2012-0002),平均體質(zhì)量(20±2)g,由寧波大學(xué)動物中心提供。
sIgA、IFN-γ、TNF-α ELISA試劑盒 南京建成生物工程公司;PG由本實驗室從嗜酸乳桿菌中提取,氧化PG由TEMPO/NaClO/NaBr選擇性氧化,保存于-20 ℃?zhèn)溆茫籅CA蛋白檢測試劑盒、RIPA強蛋白裂解液 江蘇碧云天生物試劑公司。
Thermo MK3全波長掃描多功能酶標儀 熱電(上海)儀器有限公司;DW-86L490型超低溫保存箱青島海爾特種電器有限公司;W H-3微型漩渦混合儀 上海滬西分析儀器廠有限公司;LDZX-50KBS立式壓力蒸汽滅菌器 上海申安醫(yī)療器械廠;低速冷凍離心機 北京醫(yī)用離心機廠;十二烷基硫酸鈉-聚丙烯酰胺凝膠電泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis,SDS-PAGE)儀 北京六一儀器廠。
1.2方法
1.2.1分組與造模
ICR小鼠隨機分為對照組、感染組(E. coli 0111:B4感染組)、PG治療組(高劑量0.5 mg/g,中劑量0.25 mg/g,低劑量0.125 mg/g)及氧化PG治療組(高劑量0.5 mg/g,中劑量0.25 mg/g,低劑量0.125 mg/g);每組7 只,共計8 組。飼養(yǎng)5 d后,對照組每只灌服生理鹽水0.2 mL;感染組每只用改良的注射器灌胃0.2 mL E. coli 0111:B4菌液(1×109CFU/mL),次日起每天灌服肽聚糖溶液0.2 mL,連續(xù)5 d;治療組第5天灌服0.2 mL E. coli 0111:B4菌液(1×109CFU/mL),1 h后,灌服肽聚糖溶液0.2 mL,次日起每天灌服肽聚糖溶液0.2 mL,連續(xù)5 d。末次灌胃后12 h處死動物,采集組織樣本[17]。
1.2.2腸黏膜形態(tài)學(xué)觀察
取ICR小鼠2~3 cm回腸組織,迅速用新配制的生理鹽水沖洗腸內(nèi)容物,并用質(zhì)量分數(shù)為3%的明膠溶液包埋,放入液氮中冰凍切片,染色,顯微鏡下觀察組織學(xué)變化(放大倍數(shù)為×100),圖像分析儀隨機全盲測量腸絨毛高度,隱窩深度及黏膜厚度[18]。
1.2.3小腸相關(guān)蛋白SDS-PAGE分析
取不同處理組相同部位的小腸組織1 g,迅速用無菌磷酸鹽緩沖溶液沖洗腸內(nèi)容物,之后用無菌手術(shù)剪剪碎組織,并用蛋白裂解液進行裂解處理(4 ℃,30 min)。收取裂解液,12 000 r/min離心15 min,回收上清液,經(jīng)BCA試劑盒測定,統(tǒng)一各組的蛋白質(zhì)量濃度為5 mg/mL。之后蛋白樣品加入緩沖液后水浴5 min。處理好的蛋白樣本放入-20 ℃冰箱備用[19]。
1.2.4細胞因子檢測
取ICR小鼠回腸末端5 cm,平鋪于濾紙上,縱行剪開,無菌磷酸鹽緩沖溶液反復(fù)沖洗腸腔表面后,刮下黏膜,研磨成質(zhì)量分數(shù)為10%的勻漿,4 000 r/min、4 ℃離心15 min,取0.2 mL上清液至-80 ℃冰箱待檢。取上清液及標準品各100 μL置于96 孔板中,37 ℃溫育30 min,甩干液體,洗滌液進行反復(fù)洗滌5 次后,加入酶標耦合液100 μL,37 ℃條件下溫育30 min后,甩干液體,洗滌液進行反復(fù)沖洗5 次,每孔加入底物A、B液各50 μL,37 ℃條件下反應(yīng)15 min,之后加入終止液50 μL,于450 nm波長處用酶標儀測定sIgA、IFN-γ和TNF-α含量。以上實驗步驟均嚴格按照試劑盒說明操作。
1.3統(tǒng)計學(xué)分析
2 結(jié)果與分析
2.1小腸形態(tài)學(xué)指標的測定
實驗小鼠灌服大腸桿菌后均有不同程度的腹瀉。感染組小鼠精神萎靡、飲食活動量減少、嗜睡、對外界反應(yīng)遲鈍,其中有3 只小鼠出現(xiàn)死亡;而PG及氧化PG治療組均未出現(xiàn)小鼠死亡現(xiàn)象。光鏡觀察結(jié)果如圖1所示:感染組小鼠絨毛水腫,部分小鼠絨毛上皮細胞脫落,絨毛高度和隱窩深度降低。PG治療組(高劑量0.5 mg/g)及氧化PG治療組(高劑量0.5 mg/g)小腸絨毛均完整,水腫較輕,絨毛高度和隱窩深度較大。

圖1 不同處理下的小腸黏膜形態(tài)Fig.1 Morphology of intestinal mucosa in mice observed under different treatment conditions
感染組小腸絨毛高度、隱窩深度及黏膜厚度均低于對照組,差異顯著(P<0.05)。感染組和治療組均高于對照組(P<0.05),說明PG能夠提高小鼠腸絨毛高度、隱窩深度及黏膜厚度。感染組與治療組之間沒有顯著差異(P>0.05),說明感染組與治療組療效相當;而修飾前后PG治療組與感染組沒有顯著差異(P>0.05),說明選擇性氧化修飾前后PG對預(yù)防與治療大腸桿菌感染腸黏膜療效沒有差異,結(jié)果如表1所示。由于氧化修飾后PG的提高了PG的溶解性,因此,PG的選擇性氧化修飾對于PG的溶解性能改良是一種很好的方式。

表1 各組小腸組織形態(tài)學(xué)指標的比較Table1 Comparison of intestinal morphology in different groups
2.2氧化修飾PG對小腸相關(guān)蛋白表達量的影響
根據(jù)對照組、感染組及氧化PG治療組的SDS-PAGE分析,通過上樣同等濃度的蛋白樣本(5 μg),結(jié)果發(fā)現(xiàn)在70 kD和45 kD的分子質(zhì)量區(qū)域,治療組與感染組的部分蛋白表達存在差異,而且這種差異與氧化PG的劑量存在依賴性。由此可見,氧化PG對于小腸黏膜的免疫調(diào)控作用是存在的,而且這種調(diào)控作用可能與NF-κB通路蛋白相關(guān)[20]。
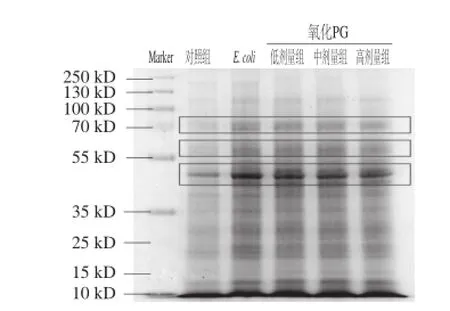
圖2 不同條件下小腸蛋白變化的SDS-PAGE分析Fig.2 Changes in total protein abundance in mouse small intestine after different treatments
2.3氧化修飾前后PG對腸黏膜分泌細胞因子的影響
2.3.1不同濃度肽聚糖對小鼠腸黏膜分泌sIgA的影響
sIgA是腸黏膜上皮細胞分泌產(chǎn)生的小體IgA與雙體IgA分子結(jié)合形成,能降低腸道蛋白酶的分解,從而形成黏膜上的抗體。由于多糖含有其他菌群的共同抗原,因此sIgA能與腸道內(nèi)有害物質(zhì)進行免疫反應(yīng),阻止這些物質(zhì)在腸道上皮的黏附和穿透,中和毒素、緩解腹瀉、胃炎及過敏反應(yīng)等癥狀[21]。由圖3可知:低劑量肽聚糖與感染組相比沒有明顯差異;原PG與氧化后PG都能夠促進黏膜分泌sIgA(0.01<P<0.05),且PG灌胃濃度越高,分泌sIgA因子越多;氧化后PG促進分泌sIgA的能力優(yōu)于原PG,與感染組之間存在極顯著差異(P<0.01)。通過灌胃PG及氧化PG,都能很好的降低E. coli 0111:B4引起的腸道炎癥反應(yīng),而且PG氧化后沒有對其功能產(chǎn)生影響,且氧化PG對sIgA分泌的促進作用更加明顯。可見PG可以通過促進sIgA的生成來提高腸道的抗炎調(diào)節(jié)作用。
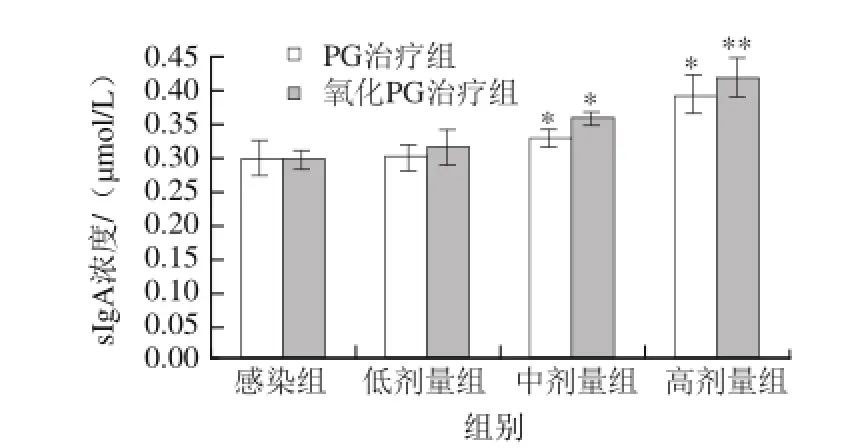
圖3 PG及氧化PG對小鼠腸黏膜組織sIgA的影響Fig.3 Effect of PG and oxidized PG on the release of sIgA in mouse intestinal mucosa
2.3.2不同肽聚糖對小鼠腸黏膜分泌IFN-γ的影響
在生物體內(nèi),IFN-γ調(diào)控一系列的生化過程,發(fā)揮著抗菌、抗癌以及相關(guān)免疫調(diào)節(jié)活性。IFN-γ的這些生物活性發(fā)揮依賴于胞內(nèi)信號途徑參與的基因調(diào)控[22]。由圖4可知:感染組與低劑量組比較,IFN-γ含量無顯著變化;中劑量組與高劑量組相對感染組,IFN-γ含量顯著下降(P<0.05);感染大腸桿菌后,小腸黏膜層細胞釋放IFN-γ細胞因子的含量升高;但隨著PG給藥濃度的升高,IFN-γ含量逐漸減低,且氧化PG治療組的效果更加顯著。表明氧化修飾對于PG功能的發(fā)揮具有很好的促進作用。

圖4 PG及氧化PG對小鼠腸黏膜組織IFN--γ的影響Fig.4 Effect of PG and oxidized PG on the release of IFN-γ in mouse intestinal mucosa
2.3.3不同濃度肽聚糖對小鼠腸黏膜分泌TNF-α的影響
大腸桿菌感染誘發(fā)腸黏膜巨噬細胞等分泌TNF-α,啟動腸道內(nèi)炎癥反應(yīng),導(dǎo)致黏膜脫落、壞死,腸道機械屏障功能受損[23]。由圖5可知,與感染組比較,低劑量組中TNF-α含量變化不大,無顯著的統(tǒng)計學(xué)差異。中劑量組與高劑量組相對感染組,TNF-α含量顯著下降(P<0.05)。實驗小鼠灌胃E. coli 0111:B4后,腸道細胞內(nèi)TNF-α含量升高,但隨著PG的濃度梯度治療作用,TNF-α含量逐漸降低。TNF-α作為重要的炎癥釋放因子,對于炎癥的產(chǎn)生,具有重要作用,它可以介導(dǎo)響應(yīng)NF-κB信號途徑,激活一系列的炎癥因子釋放[24]。本實驗中的氧化PG,作為炎癥因子的抑制劑,對于E. coli 0111:B4介導(dǎo)的小鼠腸黏膜炎癥反應(yīng)具有顯著的抑制作用,為研究嗜酸乳桿菌對腸道菌群平衡的免疫調(diào)節(jié)活性提供了良好的理論依據(jù)。
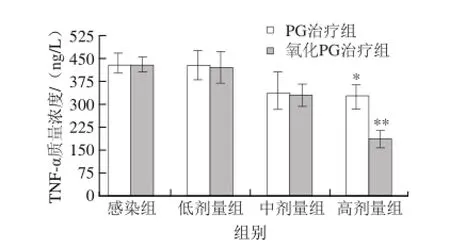
圖5 PG及氧化PG對小鼠腸黏膜組織 TNF-α的影響Fig.5 Effect of PG and oxidized PG on the release of TNF-α in mouse intestinal mucosa
3 結(jié) 論
在嗜酸乳桿菌胞壁肽聚糖對小鼠腸黏膜的免疫調(diào)控中,相比E. coli 0111:B4感染的小腸絨毛水腫,部分小鼠絨毛上皮細胞脫落, 絨毛高度和隱窩深度降低。PG及氧化PG能夠保持小腸絨毛均完整,降低大腸桿菌感染引起的水腫,絨毛高度和隱窩深度也較感染組深。在小腸黏膜免疫因子的檢測中,結(jié)果發(fā)現(xiàn),氧化后PG促進分泌sIgA的能力優(yōu)于原PG,與對照組之間存在極顯著差異(P<0.01)。灌胃PG后能夠降低IFN-γ及TNF-α的表達,且選擇性氧化修飾后PG在抑制腸炎反應(yīng)較PG具有優(yōu)勢;氧化PG不僅提高了PG的溶解度,同時高劑量組的氧化修飾PG可以顯著降低IFN-γ及TNF-α的釋放。
[1] WILSON B A, SALYERS A A, WHITT D D, et al. Bacterial pathogenesis: a molecular approach[M]. 3rd ed. Washington:American Society for Microbiology, 2011: VV055.
[2] BRANDTZAEG P. The mucosal immune system and its integration with the mammary glands[J]. Journal of Pediatrics, 2010, 156(Suppl 2):8-15.
[3] WELLS J. Mucosal vaccination and therapy with genetically modifi ed lactic acid bacteria[J]. Annual Review of Food Science and Technology, 2011, 2: 423-445.
[4] IVANOV I I, LITTMAN D R. Modulation of immune homeostasis by commensal bacteria[J]. Current Opinion in Microbiology, 2011, 14(1):106-114.
[5] 黎軼麗, 王細文. 健康教育保留灌腸法治療急性放射性腸炎25例效果觀察[J]. 齊魯護理雜志: 外科護理, 2010, 16(4): 115-116.
[6] CASTILLO N A, de LEBLANC A M, GALDEANO C M, et al. Probiotics: an alternative strategy for combating salmonellosis:immune mechanisms involved[J]. Food Research International, 2012,45(2): 831-841.
[7] GARDEN A S. Mucosittis: current management and investigations[J]. Seminars in Radiation Oncology, 2003, 13(3): 267-273.
[8] 李幼生. 腸道內(nèi)分泌功能障礙:腸功能障礙重要組成[J]. 腸外與腸內(nèi)營養(yǎng), 2014, 21(1): 1-3.
[9] KI Y, KIM W, CHO H, et al. The effect of probiotics for preventing radiation-induced morphological changes in intestinal mucosa of rats[J]. Journal of Korean Medical Science, 2014, 29(10): 1372-1378.
[10] MANTIS N J, ROL N, CORTH?SY B. Secretory IgA's complex roles in immunity and mucosal homeostasis in the gut[J]. Mucosal Immunology, 2011, 4(6): 603-611.
[11] 雷春龍, 董國忠. 腸道菌群對動物腸黏膜免疫的調(diào)控作用[J]. 動物營養(yǎng)學(xué)報, 2012, 24(3): 416-422.
[12] 王克儉, 曹玉根. 益生菌腸內(nèi)營養(yǎng)制劑在胃癌術(shù)后患者應(yīng)用的臨床研究[J]. 實用醫(yī)技雜志, 2007, 14(29): 5977-5979.
[13] 王友湘, 陳慶森. 益生菌和腸道粘膜免疫關(guān)系的研究進展[J]. 食品科學(xué), 2007, 28(8): 537-542.
[14] 孫進, 常桂芳, 樂國偉, 等. 乳桿菌肽聚糖調(diào)節(jié)小鼠免疫細胞基因表達的通路分析[J]. 細胞與分子免疫學(xué)雜志, 2008, 24(6): 553-556.
[15] 譙仕彥, 侯成立, 曾祥芳. 乳酸菌對豬腸道屏障功能的調(diào)節(jié)作用及其機制[J]. 動物營養(yǎng)學(xué)報, 2014, 26(10): 3052-3063.
[16] 楊貫羽, 郭彥春, 武光輝, 等. 氮氧自由基 TEMPO: 選擇氧化醇的高效有機小分子催化劑[J]. 化學(xué)進展, 2007, 19(11): 1727-1735.
[17] CRONIN J G, TURNER M L, GOETZE L, et al. Toll-like receptor 4 and MYD88-dependent signaling mechanisms of the innate immune system are essential for the response to lipopolysaccharide by epithelial and stromal cells of the bovine endometrium[J]. Biology of Reproduction, 20 12, 86(2): 51. doi: 10.1095/biolreprod.111.092718.
[18] 潘耀謙, 劉興友, 陳金山, 等. 急性犬瘟熱伴發(fā)寄生性腸炎的病理形態(tài)學(xué)觀察[J]. 中國獸醫(yī)科學(xué), 2012, 41(11): 1182-1186.
[19] 蘭海燕, 李立會. 蛋白質(zhì)凝膠電泳技術(shù)在作物品種鑒定中的應(yīng)用[J].中國農(nóng)業(yè)科學(xué), 2002, 35(8): 916-920.
[20] ABREU M T, FUKATA M, ARDITI M. TLR signaling in the gut in health and disease[J]. Journal of Immunology, 2005, 174(8): 4453-4460.
[21] FAMULARO G, MORETTI S, MARCELLIN I, et al. Stimulation of immunity by probiotics[M]. London: Chapman and Hill, 1997: 133-161.
[22] NGIOW S F, von SCHEIDT B, AKIBA H, et al. Anti-TIM3 antibody promotes T cell IFN-γ-mediated antitumor immunity and suppresses established tumors[J]. Cancer Research, 2011, 71(10): 3540-3551.
[23] 李永渝. 腸道屏障功能障礙的病理生理機制[J]. 胃腸病學(xué), 2006,11(10): 629-630.
[24] 秦琦瑜, 李勇, 陳虎, 等. TNF-α與IL-6在大鼠急性重癥胰腺炎并發(fā)全身炎性反應(yīng)綜合征中的作用[J]. 河北醫(yī)藥, 2008, 2(3): 140-141.
Immunoregulatory Activity of Lactobacillus acidophilus Peptidoglycan in Mouse Intestinal Mucosa
WU Zhen1, PAN Daodong1,2,*, GUO Yuxing1, ZENG Xiaoqun2, SUN Yangying2
(1. Department of Food Science and Nutrition, Nanjing Normal University, Nanjing 210097, China;2. School of Marine Science, Ningbo University, Ningbo 315211, China)
As an important immune system, mucosal immune system is the first line to protect the mucous membranes against colonization and invasion by potentially dangerous microbes that may be encountered. In this study, the immunoregulatory activity of Lactobacillus acidophilus peptidoglycan (PG) was investigated on pathogenic Escherichia coli(E. coli)-induced mouse model. Compared with PG, the oxidative modification of PG had a significant effect on the secretion of sIgA, reduced the expression of interferon-γ (IFN-γ) and tumor necrosis factor-α (TNF-α) in the intestinal mucosa. PG derived from Lactobacillus acidophilus had the capability of enhancing immune regulation of intestinal mucosa.
Lactobacillus acidophilus; peptidoglycan; intestinal mucosa; immunoregulatory activity
TS201.3
A
1002-6630(2015)13-0207-04
10.7506/spkx1002-6630-201513038
2014-12-26
國家自然科學(xué)基金面上項目(41276121;31471598);江蘇省自然科學(xué)基金面上項目(BK20141447);浙江省自然科學(xué)基金青年基金項目(Q16C200005)
吳振(1985—),男,博士研究生,研究方向為乳品科學(xué)。E-mail:woodsen@163.com
潘道東(1964—),男,教授,博士,研究方向為乳品科學(xué)。E-mail:daodongpan@163.com
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
課堂內(nèi)外·初中版(科學(xué)少年)(2023年10期)2023-12-10 00:43:06
全科護理(2022年10期)2022-12-26 21:19:15
中國合理用藥探索(2022年1期)2022-11-26 00:22:32
鄉(xiāng)村科技(2021年33期)2021-03-16 02:26:54
國際放射醫(yī)學(xué)核醫(yī)學(xué)雜志(2021年10期)2021-02-28 08:41:58
學(xué)苑創(chuàng)造·A版(2020年9期)2020-10-13 09:41:02
小學(xué)生學(xué)習指導(dǎo)(低年級)(2017年10期)2017-10-10 01:00:05
云南中醫(yī)學(xué)院學(xué)報(2014年3期)2014-07-31 18:57:34
中國合理用藥探索(2014年11期)2014-03-11 20:30:20
