FUT8基因RNAi慢病毒載體的構(gòu)建及對MCF-7細(xì)胞增殖的影響
2015-10-24 09:19:31溫憲春韓翠翠趙月生于海濤李成沖岳麗玲
生物技術(shù)通報 2015年5期
溫憲春韓翠翠趙月生于海濤李成沖岳麗玲
(1.齊齊哈爾醫(yī)學(xué)院 醫(yī)藥科學(xué)研究中心,齊齊哈爾 161006;2.齊齊哈爾醫(yī)學(xué)院附屬第三醫(yī)院 乳腺外科,齊齊哈爾 161006)
FUT8基因RNAi慢病毒載體的構(gòu)建及對MCF-7細(xì)胞增殖的影響
溫憲春1韓翠翠1趙月生2于海濤1李成沖1岳麗玲1
(1.齊齊哈爾醫(yī)學(xué)院醫(yī)藥科學(xué)研究中心,齊齊哈爾161006;2.齊齊哈爾醫(yī)學(xué)院附屬第三醫(yī)院乳腺外科,齊齊哈爾161006)
旨在構(gòu)建FUT8基因RNA干擾(RNAi)慢病毒載體并觀察其對人乳腺癌細(xì)胞MCF-7增殖的影響。針對FUT8基因設(shè)計3組短發(fā)夾RNA序列,退火合成雙鏈DNA,通過連接線性化的pGC-LV-GFP載體,構(gòu)建miRNA慢病毒載體質(zhì)粒,并將其轉(zhuǎn)化至感受態(tài)細(xì)胞DH5α;測序驗證正確后進行FUT8基因慢病毒載體的包裝及病毒滴度測定,將獲得的重組慢病毒pGC-shFUT8轉(zhuǎn)染MCF-7細(xì)胞,利用Real time-PCR、Western blot分別驗證轉(zhuǎn)染后MCF-7細(xì)胞中FUT8 mRNA及蛋白的表達,MTT法及克隆形成實驗檢測shFUT8對MCF-7細(xì)胞增殖能力的影響。測序證實成功構(gòu)建針對FUT8基因的RNAi慢病毒載體;慢病毒載體經(jīng)293T細(xì)胞包裝成功,測定病毒懸液滴度>5×108TU/mL;熒光顯微鏡下觀察各轉(zhuǎn)染組細(xì)胞GFP的表達,轉(zhuǎn)染效率達90%以上;Real-time PCR、Western blot結(jié)果顯示干擾組FUT8的mRNA及蛋白表達水平較對照組顯著降低,其中pGC-shFUT8-2序列對FUT8基因的干擾效率可達80%,干擾效果最佳,F(xiàn)UT8沉默后MCF-7細(xì)胞增殖能力下降。
FUT8基因;慢病毒載體;RNA干擾;MCF-7細(xì)胞
糖基化是最常見的翻譯后修飾反應(yīng)之一,糖基化異常直接影響細(xì)胞的識別、黏附、遷移及侵襲等生物學(xué)特性,與腫瘤的發(fā)生、發(fā)展密切相關(guān)。α-1,6巖藻糖基轉(zhuǎn)移酶(α1,6-fucosyltransferase,F(xiàn)UT8)是巖藻糖基轉(zhuǎn)移酶基因超家族的一員,其通過形成α-1,6糖苷鍵催化巖藻糖基由GDP-Fuc轉(zhuǎn)移至糖蛋白N-連接寡糖的核心結(jié)構(gòu)GlcNAc上,形成核心巖藻糖[1]。這種核心巖藻糖基化修飾普遍存在于糖蛋白中,被認(rèn)為是對糖蛋白進行重要的翻譯后修飾和功能調(diào)控的一種方式[2]。然而,在多種腫瘤惡性演進的過程中,F(xiàn)UT8的活性和表達量會逐漸增加[3-8],但FUT8作用的特異性底物以及核心巖藻糖基化修飾在乳腺細(xì)胞惡性變過程中所發(fā)揮的具體生物學(xué)功能尚不清楚。本研究采用RNA干擾技術(shù)構(gòu)建針對人FUT8基因的慢病毒表達載體,并在人乳腺癌細(xì)胞MCF-7中表達,旨在為進一步深入研究FUT8基因功能及在乳腺癌發(fā)生發(fā)展中的作用奠定實驗基礎(chǔ)。
1 材料與方法
1.1 材料
人乳腺癌MCF-7細(xì)胞、293T細(xì)胞購自上海中科院細(xì)胞庫,DH5α感受態(tài)細(xì)胞由本室保存,慢病毒載體質(zhì)粒pGC-LV-GFP、pHelper1.0、pHelper2.0為上海吉凱基因化學(xué)技術(shù)有限公司產(chǎn)品;T4 DNA連接酶、限制性內(nèi)切酶Age I和EcoR I購自NEB,Lipofectamine 2000、Trizol(Invitrogen公司),質(zhì)粒大提試劑盒(QIAGEN公司),Taq polymerase、凝膠回收試劑盒、PrimeScript RT reagent Kit(TaKaRa公司),胎牛血清、MEM培養(yǎng)基(Gibco公司),F(xiàn)UT8鼠抗人多克隆抗體(Santa公司)、HRP標(biāo)記的羊抗鼠IgG(CST公司)。Positive clone測序由上海美季生物技術(shù)有限公司完成。
1.2.1 靶向FUT8基因的shRNA慢病毒表達載體的構(gòu)建 從GenBank查找FUT8基因序列(GI:NM_004480),應(yīng)用Ambion公司的RNA干擾設(shè)計軟件,設(shè)計3條針對人FUT8基因的特異性shRNA序列(表1),以通用序列(negative control,NC)作為陰性對照。將合成好的單鏈DNA退火形成雙鏈DNA,通過T4 DNA連接酶連接線性化的pGC-LVGFP載體,并轉(zhuǎn)化至感受態(tài)細(xì)胞DH5α,菌落PCR篩選重組陽性克隆進行測序鑒定。

表1 針對FUT8基因的3個siRNA靶序列
1.2.2 RNAi慢病毒的包裝 取對數(shù)生長期的293T細(xì)胞按照1.2×107個接種于15 cm培養(yǎng)皿中,待細(xì)胞達70%-80%融合時進行轉(zhuǎn)染。取20 μg慢病毒表達質(zhì)粒pGC-LV-GFP-shFUT8、15 μg的pHelper 1.0質(zhì)粒、10 μg的pHelper 2.0質(zhì)粒與2.5 mL Opti-MEM混勻后,脂質(zhì)體Lipofectamine 2000介導(dǎo)轉(zhuǎn)染293T細(xì)胞,培養(yǎng)8 h后換為完全培養(yǎng)基,繼續(xù)培養(yǎng)48 h后收集含慢病毒顆粒的細(xì)胞上清液,于4℃,4 000×g離心10 min,上清液用0.45 μm濾器過濾,得到的病毒原液命名為pGC-shFUT8,分裝后-80℃保存?zhèn)溆谩?/p>
1.2.3 孔稀釋法測定病毒滴度 滴度測定前1 d,以8×103cells/孔的密度接種293T細(xì)胞于96孔板,體積100 μL。將10 μL病毒儲存液加入90 μL的無血清培養(yǎng)基中,在EP管中做10倍梯度稀釋,連續(xù)10個稀釋度。選取細(xì)胞孔,吸去90 μL的原培養(yǎng)基,然后在每孔中加入90 μL慢病毒稀釋液,每個稀釋度重復(fù)3孔,24 h后每孔加入100 μL完全培養(yǎng)基。4 d后根據(jù)GFP表達情況,以最大稀釋倍數(shù)孔計算病毒原液滴度,計算公式:病毒滴度(TU/mL)=熒光細(xì)胞個數(shù)×1 000/孔的病毒原液量(μL)。
1.2.4 重組慢病毒感染MCF-7細(xì)胞 將人乳腺癌MCF-7細(xì)胞以4×104cells/孔的密度接種于6孔板培養(yǎng),細(xì)胞80%以上融合時進行病毒感染。以預(yù)試驗得到的重組慢病毒的最佳MOL值100侵染人MCF-7細(xì)胞,同時設(shè)立不加病毒的空白對照組和轉(zhuǎn)染病毒空載體的陰性對照組,8 h后更換細(xì)胞上清為新鮮培養(yǎng)基。
1.2.5 實時PCR檢測MCF-7細(xì)胞FUT8 mRNA表達 轉(zhuǎn)染96 h后收集細(xì)胞,Trizol提取細(xì)胞總RNA。取1 μg總RNA進行逆轉(zhuǎn)錄反應(yīng),PCR反應(yīng)利用ABI7300熒光定量PCR儀進行。PCR反應(yīng)引物序列如下:FUT8上游引物:5'-CCATTTCAGGTTTGTTTGGTAG-3';下游引物5'-ATTGGTCCCGCTTCTCACTT-3';內(nèi)參β-actin上游引物為5'-CTGGGACGACATGGAGAAAA-3',下游引物為5'-AAGGAAGGCTGGAAGAGTGC-3'。PCR反應(yīng)條件:95℃預(yù)變性30 s;95℃變性5 s,60℃退火30 s,共40個循環(huán)。
1.2.6 Western blot檢測MCF-7細(xì)胞FUT8蛋白表達
轉(zhuǎn)染96 h后收集各組細(xì)胞,加入RIPA裂解液,提取細(xì)胞總蛋白,BCA法測定蛋白濃度。取20 μg總蛋白進行SDS-PAGE電泳,轉(zhuǎn)PVDF膜,封閉液室溫封閉2 h后依次加入鼠抗人FUT8一抗(1∶200),HRP標(biāo)記的羊抗鼠二抗(1∶3 000),TBST洗膜后ECL顯色,暗室曝光。以GAPDH作為內(nèi)參,檢測各蛋白條帶吸光強度,AFUT8/GAPDH的比值表示FUT 8蛋白相對表達量。
1.2.7 MTT法分析細(xì)胞增殖能力 各組分別取對數(shù)生長期細(xì)胞,以每孔3×103cells/200 μL接種于96孔板,37℃常規(guī)培養(yǎng)7 d,每天每組取4個平行孔細(xì)胞,加入MTT液(20 μL/孔),繼續(xù)培養(yǎng)4 h后,棄上清,每孔加入150 μL DMSO,充分震蕩后酶標(biāo)儀490 nm波長處測定OD值,繪制細(xì)胞生長曲線圖。試驗重復(fù)3次。
“感謝大家收看我們的演出,希望這次演出能給各位帶來一次與眾不同的體驗。”厚重而充滿磁性的男聲再次響起,我和抽煙男牽起手,一起做了一個謝幕的姿勢,幕簾緩緩閉合,女孩和那兩個男人臉上露出恍然大悟的表情。
1.2.8 平板克隆形成試驗 取對數(shù)生長期的細(xì)胞,胰酶消化并吹打成單細(xì)胞懸液,以 3×103cells/孔的細(xì)胞數(shù)接種于6孔板中,每組細(xì)胞設(shè)3個復(fù)孔。37℃,5% CO2靜置培養(yǎng)2-3周。PBS 清洗,甲醇固定15 min,0.2%結(jié)晶紫染色20 min,流水沖去染液。顯微鏡下計數(shù)大于50個細(xì)胞的克隆數(shù),計算克隆形成率:克隆形成率(%)=克隆數(shù)/接種細(xì)胞數(shù)×100%。
2 結(jié)果
2.1 重組慢病毒載體的測序鑒定
將3組重組載體的陽性克隆進行DNA測序,測序結(jié)果表明插入片段與設(shè)計的shRNA核苷酸序列完全一致(圖1),說明已成功構(gòu)建針對FUT 8基因的重組慢病毒載體pGC-LV-GFP-shFUT8。

圖1 慢病毒重組載體pGC-LV-GFP-shFUT8的測序結(jié)果
2.2 重組慢病毒的滴度測定
熒光顯微鏡下觀察,重組慢病毒載體轉(zhuǎn)染293T細(xì)胞48 h后各轉(zhuǎn)染組細(xì)胞均可見強綠色熒光(圖2),說明轉(zhuǎn)染成功。將病毒原液倍比稀釋分別感染293T細(xì)胞后,計數(shù)各孔中表達綠色熒光的細(xì)胞數(shù),本試驗最終測得pGC-shFUT8-1、2、3病毒原液滴度值分別為8×108、1×109和5×108TU/mL,表明病毒包裝成功。
圖2 重組慢病毒載體轉(zhuǎn)染293T細(xì)胞后觀察綠色熒光蛋白表達(40×)
2.3 重組慢病毒感染MCF-7細(xì)胞的結(jié)果
重組慢病毒感染MCF-7細(xì)胞96 h后,熒光顯微鏡下可見90%以上細(xì)胞發(fā)出綠色熒光(圖3),細(xì)胞傳代后綠色熒光仍可持續(xù)表達,證明重組慢病毒成功轉(zhuǎn)染MCF-7細(xì)胞。
圖3 慢病毒轉(zhuǎn)染MCF-7細(xì)胞后熒光顯微鏡觀察綠色熒光(100×)
2.4 慢病毒轉(zhuǎn)染MCF-7細(xì)胞后下調(diào)FUT8的mRNA表達
Real-time PCR結(jié)果(圖4)顯示,空白對照組與陰性對照組在mRNA表達水平方面差異無統(tǒng)計學(xué)意義(P>0.05);與對照組相比,pGC-shFUT8實驗組的FUT8 mRNA表達水平均有不同程度降低,shFUT8-1、2、3的抑制率分別為30%、54%和40.82%,其中pGC-shFUT8-2在mRNA表達水平抑制效果最為顯著(P<0.05)。
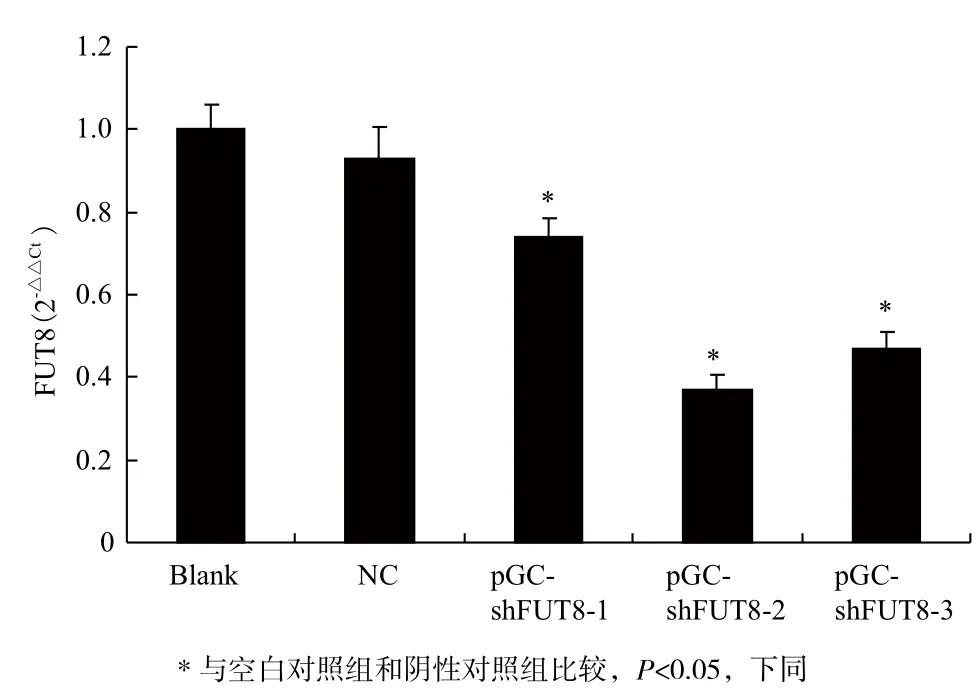
圖4 Real-time PCR檢測FUT8基因的mRNA表達水平
2.5 慢病毒轉(zhuǎn)染MCF-7細(xì)胞后下調(diào)FUT8的蛋白表達
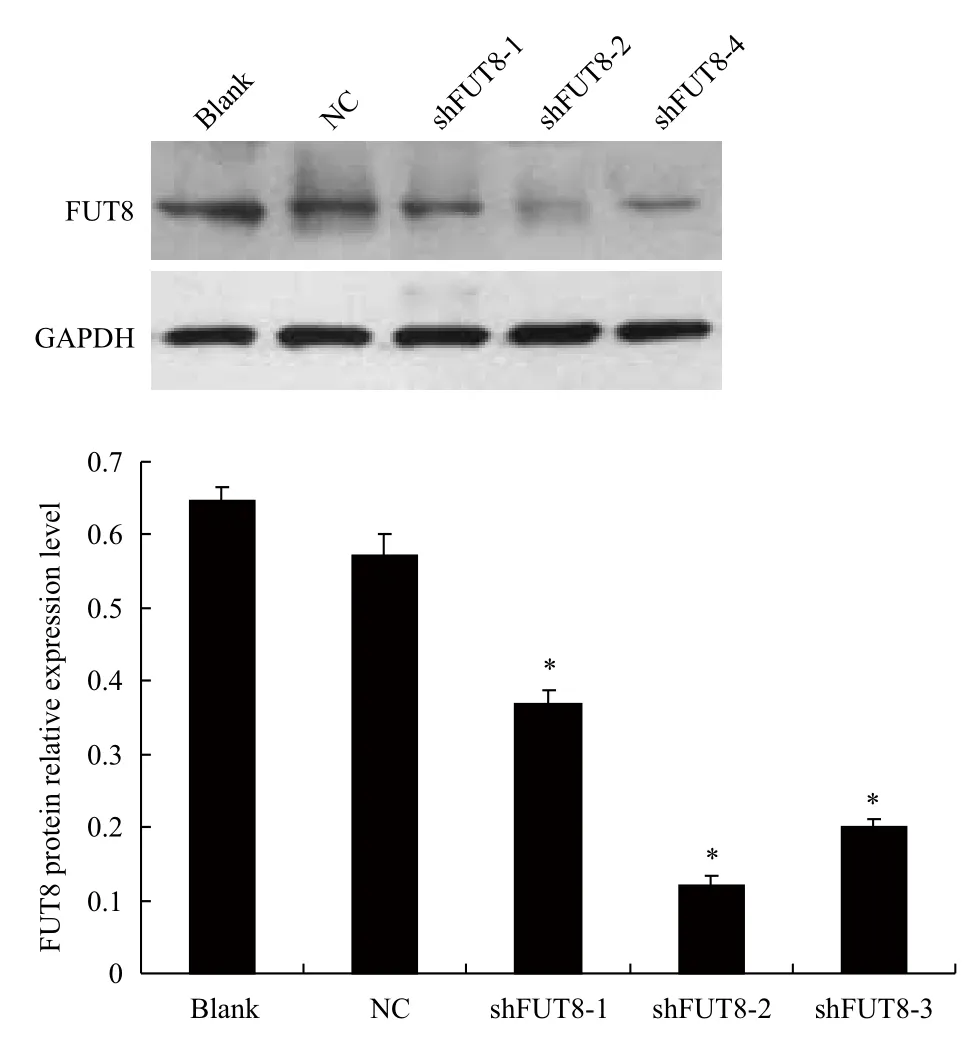
圖5 Western blotting 檢測FUT8蛋白表達水平
重組慢病毒載體轉(zhuǎn)染MCF-7細(xì)胞后Western blotting檢測結(jié)果(圖5)顯示,3個RNAi組的FUT8蛋白相對表達量較空白對照組與陰性對照組明顯降低,shFUT8-1、2、3組分別下降了43.08%、80%和69.23%,其中pGC-shFUT8-2組抑制效果最佳(P<0.05);空白對照組與陰性對照組間FUT8蛋白表達量無顯著性差異(P>0.05)。
2.6 細(xì)胞體外增殖分析
MTT檢測結(jié)果發(fā)現(xiàn),接種第3天起,pGC-shFUT8-2組細(xì)胞增殖速率明顯低于空白對照組與陰性對照組(P<0.05),而陰性對照組與空白對照組相比較,增殖未見改變(P>0.05,圖6)。同時,克隆形成試驗也顯示,轉(zhuǎn)染pGC-shFUT8-2組細(xì)胞克隆形成率(16.96%)遠(yuǎn)低于空白對照組及陰性對照組(分別為41.38%、39.67%),差異顯著(P<0.05),而陰性對照組與空白對照組比較無顯著性差異(P>0.05,圖7)。
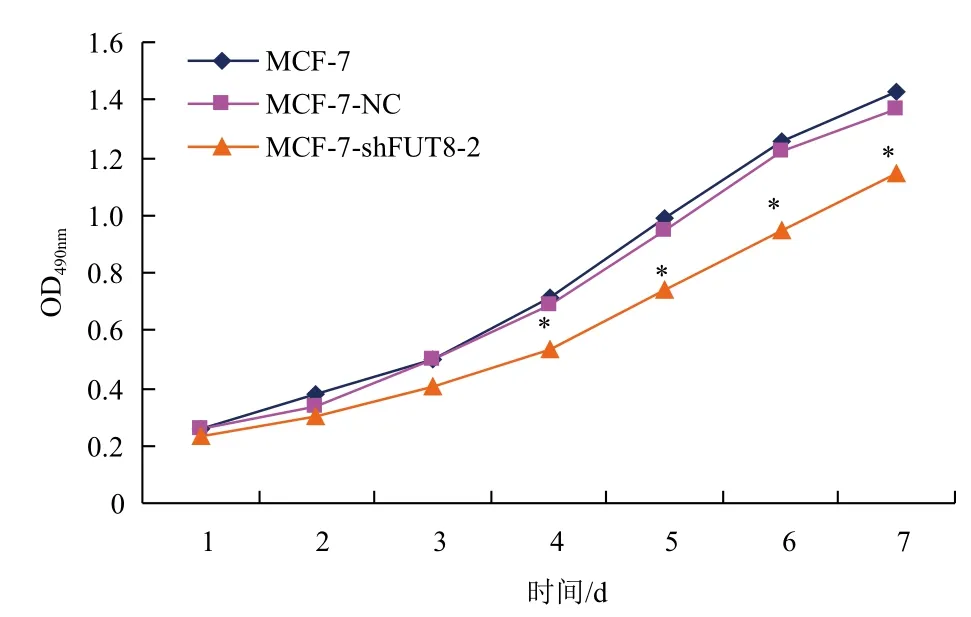
圖6 各組MCF-7細(xì)胞生長曲線圖
3 討論
細(xì)胞惡性轉(zhuǎn)化過程中總伴有糖鏈結(jié)構(gòu)的改變,并且這種改變與腫瘤細(xì)胞的黏附、遷移及侵襲等多種運動功能相關(guān)[9]。核心巖藻糖是位于N-型糖鏈核心部位的唯一的巖藻糖糖基,F(xiàn)UT8是催化核心巖藻糖基反應(yīng)的酶[10]。現(xiàn)已證明核心巖藻糖基對多種生長因子和黏附分子(如EGF、E-cadherins、TGF-β和Integrin等)的信號傳導(dǎo)是必不可少的[11-15]。如FUT8基因缺失小鼠表現(xiàn)出肺氣腫樣改變的表型和嚴(yán)重的發(fā)育遲緩,出生后3 d之內(nèi)有70%的死亡率[16]。在腫瘤細(xì)胞常常可以見到FUT8的改變,其通過催化多種底物蛋白,如芳基硫酸酯A(ARSA)、5T4糖蛋白癌胚抗原、甲胎蛋白、纖連蛋白和鈣粘蛋白等,參與底物分子對腫瘤發(fā)生、發(fā)展及轉(zhuǎn)移潛能相關(guān)的作用途徑[17,18]。因此,F(xiàn)UT8基因在腫瘤細(xì)胞的生長和轉(zhuǎn)移過程中發(fā)揮著重要的調(diào)節(jié)作用。
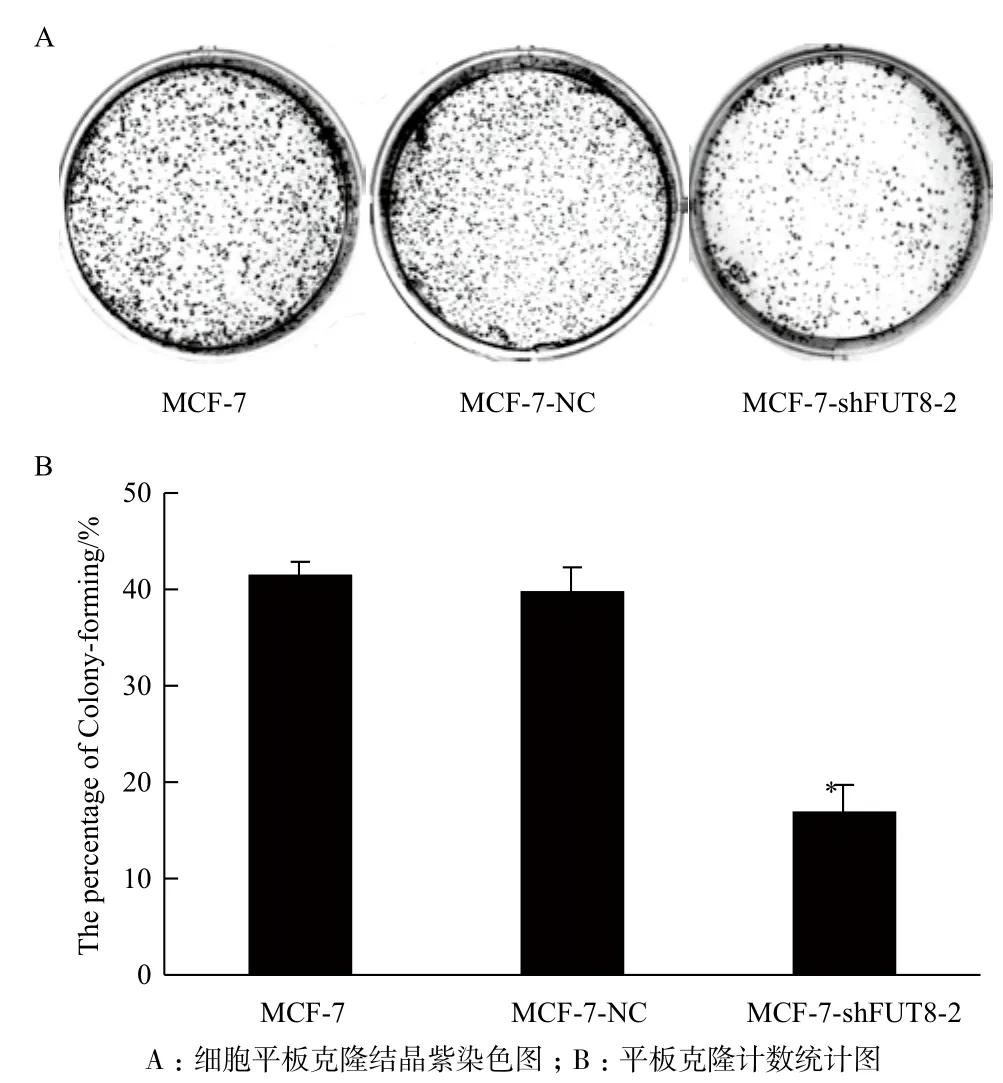
圖7 各組MCF-7細(xì)胞平板克隆形成能力分析
RNA干擾(RNAi)是一種準(zhǔn)確和有效的基因沉默的方法,是由短雙鏈RNA(dsRNA)誘導(dǎo)的同源mRNA高效特異性降解,從而導(dǎo)致目的基因表達沉默[19]。常用的siRNA的表達載體有質(zhì)粒、腺病毒、慢病毒,其中病毒載體感染效率更高。慢病毒作為外源基因載體具有感染范圍廣、表達穩(wěn)定長效、基因裝載量大等優(yōu)點,是目前進行基因功能研究和基因治療的有力工具。本研究設(shè)計合成3對靶向FUT8基因的shRNA,構(gòu)建siRNA表達的慢病毒干擾載體,并利用病毒感染人乳腺癌MCF-7細(xì)胞,以期通過RNAi機制實現(xiàn)對MCF-7細(xì)胞中FUT8基因的沉默。經(jīng)熒光顯微鏡觀察顯示,本試驗構(gòu)建的3組慢病毒干擾載體pGC-shFUT8均可高效率轉(zhuǎn)染MCF-7細(xì)胞,轉(zhuǎn)染效率均在90%以上,并能有效抑制FUT8的mRNA及蛋白的表達,其中pGC-shFUT8-2為最佳靶向FUT8的干擾序列。另外,我們對FUT8基因的功能進行了初步研究,選用干擾效果最佳的pGC-shFUT8-2序列轉(zhuǎn)染MCF-7細(xì)胞,MTT檢測發(fā)現(xiàn)FUT8干擾的MCF-7細(xì)胞生長增殖能力較正常的MCF-7明顯下降;同樣反映細(xì)胞增殖能力的克隆形成試驗也顯示轉(zhuǎn)染pGC-shFUT8-2后,MCF-7細(xì)胞的克隆形成率比轉(zhuǎn)染前顯著降低,表明FUT8干擾后對乳腺癌細(xì)胞增殖具有抑制作用,而FUT8基因的其它生物學(xué)功能及其調(diào)控機制尚需進一步深入研究。
4 結(jié)論
本研究成功構(gòu)建靶向人FUT8基因的RNAi慢病毒載體,并篩選出高效干擾MCF-7細(xì)胞FUT8基因表達的有效靶點,初步證實沉默F(xiàn)UT8對乳腺癌細(xì)胞具有增殖抑制作用。
[1] Sasaki H, Toda T, Furukawa T,et al. α-1, 6-fucosyltransferase(FUT8)inhibits hemoglobin production during differentiation of murine and K562 human erythroleukemia cells[J]. Journal of Biological Chemistry, 2013,288(7):16839-16847.
[2] Uozumi N, Yanagidani S, Miyoshi E, et al. Purification and cDNA cloning of porcine brain GDP-L-Fuc:N-acetyl-β-D-glucosaminide α1→ 6fucosyltransferase[J]. J Biol Chem, 1996, 271(44):27810-27817.
[3] Hutchinson WL, Du MQ, Johnson PJ, et al. Fucosyltransfera-ses:differential plasma and tissue alterations in hepatocellular carcinoma and cirrhosis[J]. Hepatology, 1991, 13(4):683-688.
[4] Miyoshi E, Noda K, Ko JH, et al. Overexpression of α1-6 fucosyltransferase in hepatoma cells suppresses intrahepatic metastasis after splenic injection in athymic mice[J]. Cancer Res, 1999, 59(9):2237-2243.
[5] Ito Y, Miyauchi A, Yoshida H, et al. Expression of alpha1,6-fucosyltransferase(FUT8)in papillary carcinoma of the thyroid:its linkage to biological aggressiveness and anaplastic transformation[J]. Cancer Lett, 2003, 200(2):167-172.
[6] Takahashi T, Ikeda Y, Miyoshi E, et al. α1, 6 fucosyltransferase is highly and specifically expressed in human ovarian serous adenocarcinomas[J]. Int J Cancer, 2000, 88(6):914-919.
[7] Muinelo-Romay L, Villar-Portela S, Cuevas E, et al. Identification of α(1, 6)fucosylated proteins differentially expressed in human colorectal cancer[J]. BMC Cancer, 2011, 11(10):508-519.
[8] Potapenko IO, Haakensen VD, Lüders T, et al. Glycan gene expression signatures in normal and malignant breast tissue;possible role in diagnosis and progression[J]. Molecular Oncology, 2010, 4(2):98-118.
[9] Ito H, Hiraiwa N, Sawada-Kasugai M, et al. Altered mRNA expression of specific molecular species of fucosyl-and sialyltransferases in human colorectal cancer tissues[J]. Int J Cancer,1998, 71(4):556-564.
[10] Miyoshi E, Moriwaki K, Nakagawa T, et al. Biological function of fucosylation in cancer biology[J]. J Biochem,2008, 143(6): 725-729.
[11] Wang X, Gu J, Ihara H, et al. Core fucosylation regulates epidermal growth factor receptor-mediated intracellular signaling[J]. J Biol Chem, 2006, 281(5):2572-2577.
[12] Matsumoto K, Yokote H, Arao T, et al. N-Glycan fucosylation of epidermal growth factor receptor modulates receptor activity and sensitivity to epidermal growth factor receptor tyrosine kinase inhibitor[J]. Cancer Sci, 2008, 99(8):1611-1617.
[13] Liu YC, Yen HY, Chen CY, et al. Sialylation and fucosylation of epidermal growth factor receptor suppress its dimerization and activation in lung cancer cells[J]. Proc Natl Acad Sci USA,2011, 108(28):11332-11337.
[14] Lin H, Wang D, Wu T, et al. Blocking core fucosylation of TGF-β1 receptors downregulates their functions and attenuates the epithelial-mesenchymal transition of renal tubular cells[J]. Am J Physiol Renal Physiol, 2011, 300(4):1017-1025.
[15] Venkatachalam MA, Weinberg JM. New wrinkles in old receptors: core fucosylation is yet another target to inhibit TGF-β signaling[J]. Kidney International, 2013, 84(1):11-14.
[16] Zhao Y, Itoh S,Wang X, et al. Deletion of core fucosylation on α3β1 integrin down-regulates its functions[J]. J Biol Chem,2006, 281(50):38343-38350.
[17] Wang M, Long RE, Comunale MA, et al. Novel fucosylated biom arkers for the early detection of hepatocellular carcinoma[J]. Cancer Epidemiol Biomarkers Prev, 2009, 18(6):1914-1921.
[18]Geng F, Shi BZ, Yang YF, Wu XZ. The expression of core fucosylated E-cadherin in cancer cells and lung cancer patients:prognostic implications[J]. Cell Res, 2004, 14(5):423-433.
[19] Pushparaj PN, Aarthi JJ, Manikandan J, et al. siRNA, miRNA, and shRNA:in vivo applications[J]. J Dent Res, 2008, 87(11):992-1003.
(責(zé)任編輯 馬鑫)
Construction of FUT8 Gene Lentiviral RNA Interference Vector and Regulation on Proliferation of Human Breast Cancer Cells MCF-7
Wen Xianchun1Han Cuicui1Zhao Yuesheng2Yu Haitao1Li Chengchong1Yue Liling1
(1. Research Center of Medical Science,Qiqihar Medical College,Qiqihar161006;2. Breast Surgery,the Third Affiliated Hospital of Qiqihaer Medical College,Qiqihar161006)
This study aimed to construct the lentiviral RNA interference(RNAi)vector of human FUT8 gene and determine its effect on proliferation of human breast cancer cells MCF-7. Designing three short hairpin RNA(shRNA)sequence targeting the FUT8 gene, and then synthesizing the complementary DNA chains containing the target sequence, miRNA lentiviral vector plasmid with linear pGC-LV-GFP carriers were constructed and then transformed to DH5α cells. After identified by sequencing, then packaging lentiviral vectors and measuring the virus titer were done. The recombinant lentiviral vector pGC-shFUT8 was transfected into human MCF-7 cells, and then the expression of FUT8 in transfected MCF-7 cells was detected by real time-PCR and Western Blotting. The effect of shFUT8 on MCL-7 cell proliferation was measured by MTT assay and colony formation experiment. Gene sequencing confirmed the successful construction of RNAi lentiviral vector targeting the FUT8 gene. The lentiviral vectors were packed successfully by 293T cells and the virus titer was more than 5×108TU/mL. Expressed GFP in transfected cells were observed under the fluorescence microscope, and the transfection efficiency was over 90%. Real-time PCR and Western Blotting analysis indicated that the expression levels of FUT8 mRNA and protein in the transfected group significantly reduced when comparing with the negative control group. Particularly the pGC-shFUT8-2 sequence could interfere 80 % of FUT8 gene expression. The proliferation of MCF-7 decreased after FUT8 was inactive.
FUT8 gene;Lentiviral vector;RNA interference;MCF-7cells
10.13560/j.cnki.biotech.bull.1985.2015.05.035
2014-08-25
國家自然科學(xué)基金項目(81202084),教育部科學(xué)技術(shù)研究重點項目(212047)
溫憲春,男,碩士,研究方向:腫瘤藥理學(xué);E-mail:wenxianchun@sina.com
岳麗玲,女,教授,碩士生導(dǎo)師,研究方向:腫瘤分子生物學(xué);E-mail:yuell1025@126.com
