西藏藏豬遺傳多樣性研究
2015-10-24 09:19:30郭永博蔡原
生物技術通報 2015年5期
關鍵詞:研究
郭永博 蔡原
(甘肅農業(yè)大學動物科學技術學院,蘭州 730070)
西藏藏豬遺傳多樣性研究
郭永博 蔡原
(甘肅農業(yè)大學動物科學技術學院,蘭州 730070)
采用mtDNA D-loop作為分子標記,對西藏的4個藏豬群體(林芝、山南、昌都和日喀則)遺傳多樣性進行了研究。結果表明,西藏藏豬mtDNA D-loop高變區(qū)A+T含量(62.90%)明顯高于 G+C 含量(37.1%),富含A和T,存在堿基偏倚性。在長度為435 bp的序列中,共檢測到20個變異位點,界定了26個單倍型,單倍型多樣度(Hd)為0.705±0.021,平均核苷酸差異數(k)為1.231,核苷酸多樣度(Pi)為0.002 83。其中,Hd、k和Pi在昌都藏豬群體中最高,日喀則藏豬最低。此外,Hap1和Hap3單倍型是4個群體的共享單倍型,表明4個藏豬群體存在兩個共同的母系祖先單倍型。
藏豬;mtDNA D-loop;單倍型;遺傳多樣性
藏豬是世界上少有的高原型豬種,是唯一能夠適應高原海拔氣候和以放牧為主的豬種[1]。藏豬主產于青藏高原,包括云南迪慶藏豬、四川阿壩及甘孜藏豬、甘肅的合作豬以及分布于西藏自治區(qū)山南、林芝、昌都等地的藏豬類群。由于近年來繁殖育種手段不斷改進,高強度選育,加之生態(tài)環(huán)境的改變,致使許多擁有豐富遺傳資源的地方品種數量銳減,純種藏豬的分布范圍和數量正在不斷減少,已面臨種群滅絕的危險[2]。因此,最大限度地保存藏豬的遺傳多樣性是藏豬保護與利用面臨的突出問題。
mtDNA D-loop,即mtDNA的非編碼區(qū),該區(qū)的進化速率較其他區(qū)域高5-10倍,是目前mtDNA研究的熱點,被廣泛應用于哺乳動物遺傳多樣性和種內、種間親緣關系等方面的研究[3-5],甘佳等[6]以微衛(wèi)星標記測定了四川境內阿壩、稻城和德格3個藏豬遺傳多樣性。近年來,應用mtDNA研究豬的遺傳多樣性及系統(tǒng)發(fā)育比較多[7-9],Gou等[10]評估了隨機和選擇性繁殖豬種群之間的遺傳多樣性,結果表明隨機繁育種群單倍型多樣性顯著大于選擇性繁育種群;Li等[11]研究了西藏野豬的遺傳適應與高海拔有關,表征了家豬唾液分泌增加的遺傳基礎;Jiao等[12]測定藏豬的遺傳多樣性發(fā)現,合作豬目前具有較高的遺傳多樣性,可以進行合理利用;Zhao等[13]利用mtDNA研究了中國豬的起源和演化,表明了中國國內豬可能是起源于野生長江中游地區(qū)及中國南方的公豬;Gou等[14]通過研究藏豬、亞洲家豬和野豬mtDNA高變區(qū),認為在西藏高原和東南亞島嶼地區(qū)存在家豬的兩個新的起源中心;張亞平等[15]測定了云南4 個地方豬品種的mtDNA D-loop高變區(qū),結果顯示mtDNA 多態(tài)能有效作為品種內遺傳多樣性指標,及云南4個地方豬品種大多數個體起源于一個野豬亞種[15]。以mtDNA分子標記研究西藏藏豬遺傳多樣性報道較少,因此,本研究對分布在西藏4個藏豬mtDNA D-loop高變區(qū)序列進行研究,以期為藏豬資源保護提供參考依據。
1 材料與方法
1.1 材料
本研究采集了西藏林芝藏豬、山南藏豬和昌都藏豬3個群體共267頭藏豬血液樣品,詳見表1。
1.2 方法
1.2.1 試驗方法 參照 《分子克隆實驗指南》[16],采用常規(guī)酚-氯抽提法提取基因組DNA。根據發(fā)表在GenBank中的西藏藏豬mtDNA D-loop序列(Accession:AF486868)設計引物:上游引物(PL16405):5'-ATACCAATCACTAGCATCAT-3';下游引物(P-H653):3'-CCGGATCATGAGTTCCATGAAGT-5'。
進行聚合酶鏈式反應,PCR擴增反應體系體積 為:60 μL, 其 中:10×Buffer 5.0 μL(Mg2+1.5 mmol/L)、dNTP(2.5 mmol/L)1.0 μL、上下游引物各5 μL、Taq DNA聚合酶(5 U/μL)0.25 μL、模板DNA 1 μL,加雙蒸水至28.0 μL。擴增條件:94℃ 2 min;94℃ 20 s,58℃ 30 s,72℃ 1.5 min,35個循環(huán);72℃ 10 min,4℃保存。PCR產物經1%的瓊脂糖凝膠電泳檢測,擴增產物送上海生工生物工程股份有限公司純化并進行雙向測序。
1.2.2 數據處理 測得的原始序列,通過Chromas-Version2.33(http://www.technelysium. com.au/chromas. html)進行人工編輯,校對電泳峰圖與堿基對應關系,剪切非研究區(qū)域序列。分析中引用來自于GenBank(http://www.ncbi.nlm.nih.gov/Genbank/index.html)的西藏藏豬相關序列,見表1。并用 MEGA 5.0(http://www.megasoftware.net)建立序列數據庫,采用 Clustal X 1.8(http://www.igbmc.ustrasbg.fr/pub/ ClustalX)進行同源序列比對分析。采用 DnaSP 5.10.1(http://www.ub.edu/dnasp)進行核苷酸變異位點、單核苷酸多態(tài)性、單倍型數目、單倍型多樣性分析以及計算單倍型多樣度、核苷酸多樣度和平均核苷酸差異度。
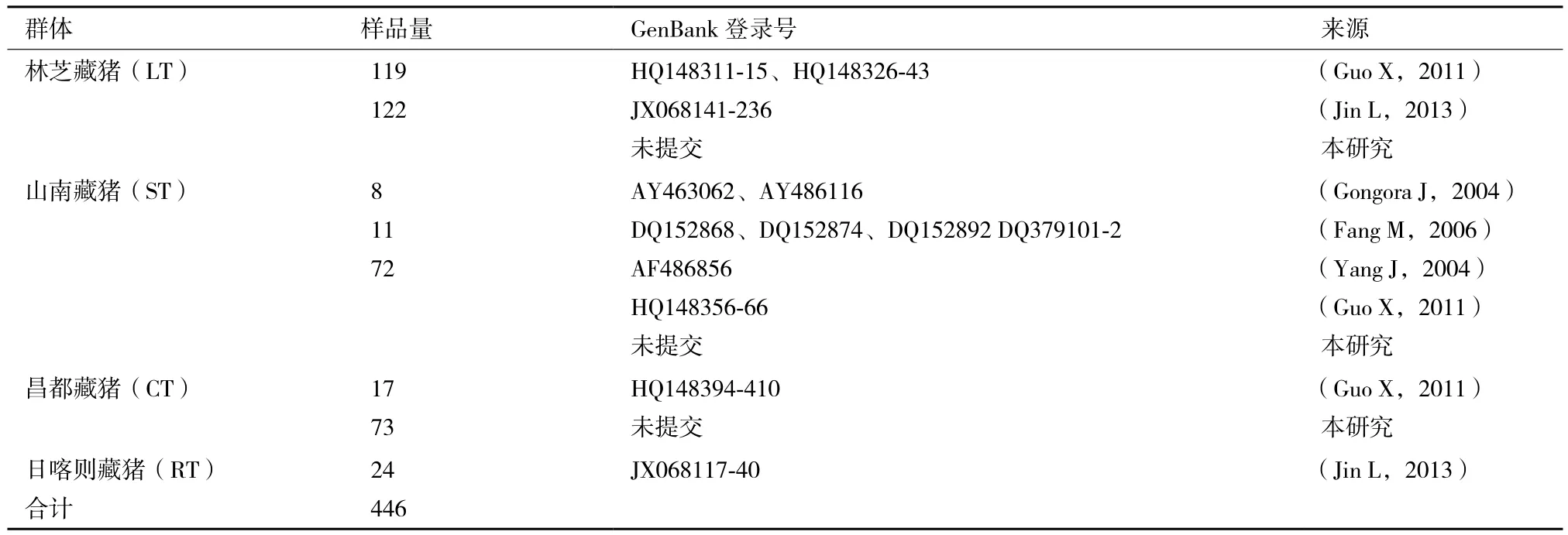
表1 樣品信息
2 結果
2.1 mtDNA D-loop高變區(qū)SNPs分析
西藏藏豬mtDNA D-loop高變區(qū)經擴增、 測序、編輯后,獲得長度為435 bp核苷酸序列。用MAGE5.0軟件進行堿基分析發(fā)現,T、C、A和G 4種核苷酸的平均比例分別為27%、24%、35.90%和13.10%,A+T 含量(62.90%)明顯高于 G+C 含量(37.1%),可見藏豬mtDNA D-loop高變區(qū)富含A和T,同時體現了哺乳動物線粒體DNA堿基組成的基本特點。利用Dnasp5.10.1軟件對4個藏豬446個個體的 mtDNA D-loop 區(qū)進行單核苷酸多態(tài)性分析(single nucleotide polymorphism,SNP),共檢測到20個多態(tài)位點,占所測核苷酸的4.60%,說明該核苷酸序列突變率小,有較好的穩(wěn)定性,其中有3個單一多態(tài)位點和17個簡約信息位點(表2)。
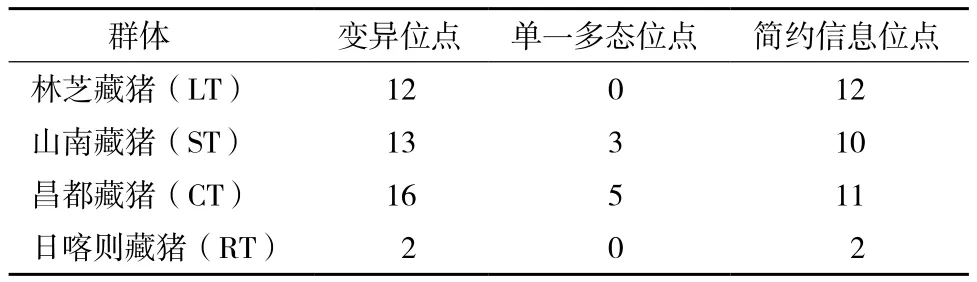
表2 西藏藏豬mtDNA D-loop高變區(qū)變異位點數統(tǒng)計
堿基變異數(S)分布結果(圖1-A)表明,在第1-40個堿基之間沒有發(fā)現堿基變異,說明這是一個相對保守的區(qū)域,變異區(qū)在第41個堿基與435個堿基之間,而高變區(qū)集中在第 157-417 堿基之間。核苷酸多樣度(Pi:序列間每個位點的平均核苷酸差異數)變化趨勢(圖 1-B)顯示,D-loop區(qū)核苷酸多樣度最高峰值出現在第250堿基左右(圖1)。
圖 1 mtDNA D-loop高變區(qū)序列堿基變異(A)及核苷酸多樣度(B)分布
2.2 遺傳多樣性分析
核苷酸多樣性分析表明,西藏藏豬單倍型多樣度(Hd±Sd)、核苷酸多樣度(Pi)和平均核苷酸差異數(k)分別為0.705±0.021、0.002 83和1.231。其中,山南藏豬和昌都藏豬兩個群體的3個多樣性參數均高于西藏藏豬的整體水平,而林芝藏豬和日喀則藏豬群體均低于整體水平(表3)。無論從單倍型數、單倍型多樣度還是核苷酸多樣度來看,昌都藏豬遺傳多樣性最高,而日喀則藏豬最低。
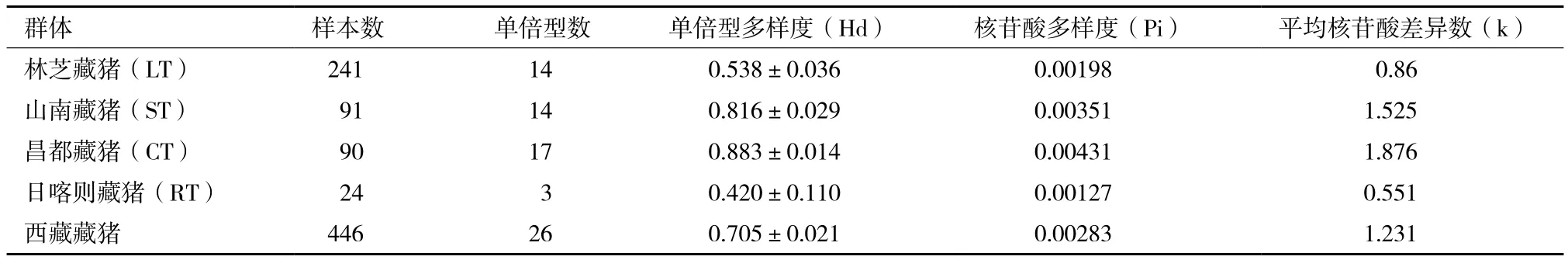
表3 西藏藏豬mtDNA D-loop遺傳多樣性
2.3 單倍型分析
西藏藏豬單倍型分析表明,根據檢測到的20個多態(tài)位點,界定了26個單倍型(Hap1-Hap26,表4)。其中,兩個及以上群體的共享單倍型共有13個,而4個群體共享的只有2個(Hap1和Hap3)。群體間的特有單倍型數差異較大,除日喀則藏豬沒有特有單倍型,其他3個群體都存在特有單倍型,其中林芝藏豬特有單倍型數最多(6個),占單倍型總數的23.08%,山南藏豬特有單倍型最少(3個)。單倍型頻率差異較大,Hap1單倍型頻率最高,有229個個體,Hap6、Hap3、Hap9和Hap16頻率依次降低,個體數分別為51、42、32和29。其余11個單倍型頻率都低于10。
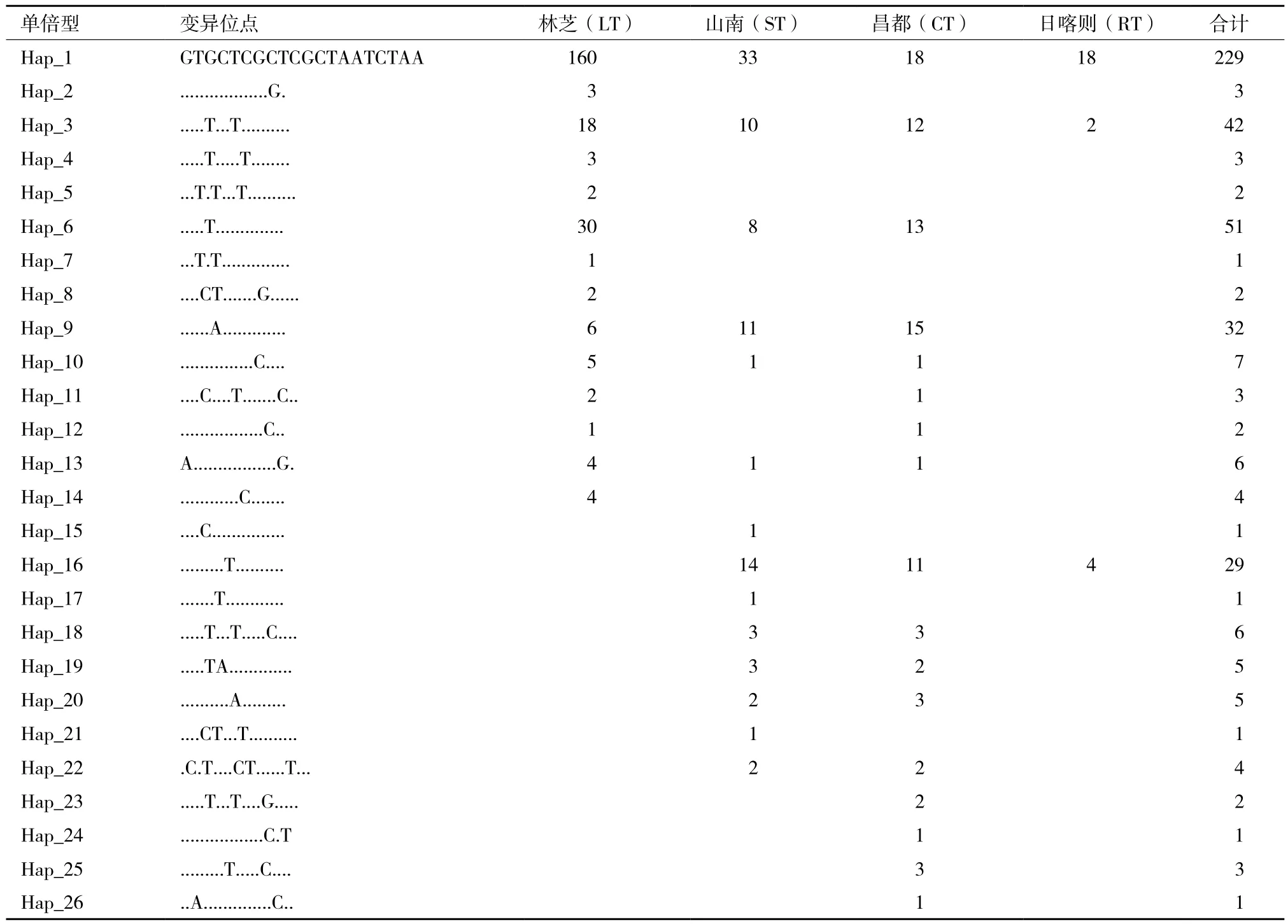
表4 西藏藏豬mtDNA D-loop高變區(qū)變異位點分布
3 討論
3.1 藏豬mtDNA D-loop高變區(qū)序列特征
由于飼養(yǎng)量下降和雜交改良等因素,藏豬資源受到嚴重威脅[17],現已被列入《 國家級畜禽遺傳資源保護名錄》。開展藏豬種質資源調查研究,可為其遺傳資源保護提供科學依據。本研究通過西藏藏豬mtDNA D-loop序列遺傳多樣性來揭示遺傳資源的保護必要性。在分析的446頭西藏藏豬mtDNA D-loop長度為435 bp的核苷酸序列中,不考慮插入、缺失和 Poly(C)末端長度變異的條件下,共檢測到20個變異位點,占核苷酸總數的4.60%,表明西藏藏豬具有較低的遺傳變異,其中,昌都藏豬堿基變異位點數最多(16個),日喀則最少(2個),說明昌都藏豬受選育程度和人工選擇機率較其他3個群體高,而日喀則藏豬可能由于地域差異、長期近交使得基因群體內具有較高的遺傳一致性,這與蔡原等[7]研究一致。mtDNA分子是由很不均一的片段構成的,G+C含量在21%-50%,其中無脊椎動物為21%-43%,脊椎動物為37%-50%[18],本試驗中為37.10%,略高于變化范圍最低值。堿基組成分析表明,A+T平均含量62.90%,顯著高于G+C含量37.10%。其中,A+T的含量可以反映出序列的變異性,具有高比例的A+T可能是 D-loop 序列變異較快的原因之一,同時也說明mtDNA D-loop高變區(qū)富含A、T,表現出很強的堿基偏倚性,這一結果與mtDNA D-loop 序列進行家禽[19,20]、羊[21,22]、牛[23,24]和豬[25,26]等家養(yǎng)動物的研究結果一致。
3.2 西藏藏豬遺傳多樣性分布規(guī)律
單倍型多樣度(Hd)和核苷酸多樣度(Pi)是衡量一個mtDNA 變異程度的兩個重要指標,Hd值和Pi值越大,多樣性程度越高,遺傳多樣性越豐富,反之,多樣性程度越低,遺傳多樣性越貧乏。另外,mtDNA的單倍型多樣性指數也可以衡量種內的變異程度,它的含義類似于核DNA的雜合度[27]。據統(tǒng)計同種動物的個體之間的平均核苷酸順序奇異值在0.3%-4.0%,有時甚至可高達10%[28]。本研究的西藏藏豬4個,總的單倍型數20個,單倍型多樣度(Hd±Sd)為0.705±0.021,平均核苷酸差異數(k)為1.231,核苷酸多樣度(Pi)為 0.002 83。研究結果顯示,昌都藏豬、山南藏豬單倍型多樣度、平均核苷酸差異數、核苷酸多樣度均高于整體水平,而林芝藏豬低于整體水平,日喀則藏豬明顯低于整體水平,并且依次降低,證明昌都和山南藏豬品種的遺傳變異程度較高,遺傳基礎比較廣泛,具有豐富的遺傳多樣性[29],可能由于昌都藏豬的群體數量較大且分布范圍較廣,而日喀則藏豬則局限在一個較為封閉的環(huán)境中,與其它群體之間基因交流的機會極少[30]。基于mtDNA D-loop序列對梅山豬、二花臉、蘇鐘豬和保山豬遺傳多樣性分析表明,其Hd和Pi值分別為0.784、0.400、0.838、0.883和0.005 6、0.000 9、0.004 7、0.003 9,表明二花臉的多樣性低于其他3個豬種[31]。采用相同的研究方法研究表明合作、迪慶、甘孜、阿壩4個藏豬群體Hd分別為0.963、0.930、0.303和0.830,Pi分別為0.008 2、0.005 5、0.000 7和0.003 6[12]。本研究中,藏豬Hd和Pi分別為0.705和0.002 83,與梅山豬、二花臉、蘇鐘豬和保山豬及藏豬合作群體、迪慶群體和阿壩群體相比,其遺傳多樣性僅高于二花臉豬和藏豬中的甘孜群體,說明了西藏藏豬群體受人工選擇的影響小,長期自然條件產生的突變類型在該群體中得到積累,但可能由于與外界交流少,群體內近親高度繁殖,使其遺傳多樣性比較有限[32]。同時,瓶頸效應與人工選擇共同導致了西藏藏豬等在內的中國家豬遺傳多樣性的降低,特別是高強度選擇豬種對地方豬種的雜交產生的后果更為嚴重。對26個單倍型進行區(qū)域分布研究發(fā)現,昌都藏豬單倍型多樣度最高,也高于西藏藏豬的整體水平,同時相對于其它3個群體,單倍型數最多,也說明昌都藏豬的遺傳多樣性最豐富。聯合國糧農組織歷年關于家養(yǎng)動物報告中都涉及到有關遺傳多樣性調查的內容,不斷改進和完善保存理論和保存機制,不斷加大各區(qū)域保種項目的投資力度[33],因此,在藏豬遺傳資源保護過程中應該遵循遺傳多樣性原則進行優(yōu)先保護。
3.3 西藏藏豬系統(tǒng)發(fā)生關系
Hap1和Hap3是4個群體的共享單倍型,說明可能具有共同的兩個母系起源。有研究表明,包括藏豬在內的家豬來源于歐洲野豬和亞洲野豬[34-36],而且是多次馴化事件的結果[37],就不同藏豬群體而言存在著共同的母系起源,但在幾千年的馴化中形成了不同的進化分枝,由于生活環(huán)境差異各群體也存在一定數量的特有單倍型,且頻率差異較大[25],其中藏豬的進化分支有一定的地理特征,即群體特異性,歐洲起源家豬與亞洲起源家豬及藏豬明顯的分為兩個不同的進化枝,在藏豬群體中西藏藏豬與甘孜豬處于同一個進化枝,而合作豬形成了一個相對獨立的進化分枝,阿壩豬在各個進化枝中都有出現[12],而西藏藏豬中除日喀則藏豬沒有特有單倍型,其它3個都存在特有單倍型,其中林芝藏豬特有單倍型數最多(6個),占單倍型總數的23.08%,山南藏豬特有單倍型最少(3個)。可能由于日喀則藏豬沒有經過長期的自然選擇和人工選擇,受人工選育的程度低,致使該群體沒有形成各自獨特的遺傳結構,其余3個群體雖然在遺傳結構上有獨特性,但它們都屬于一個地區(qū),與日喀則藏豬群體有密切的關系[38,39]。
4 結論
本研究對西藏藏豬mtDNA D-loop高變區(qū)特征、遺傳多樣性分布規(guī)律和系統(tǒng)發(fā)生關系研究表明,昌都藏豬的遺傳多樣性最高,山南藏豬、林芝藏豬、日喀則藏豬遺傳多樣性依次降低,說明昌都藏豬的群體數量較大且分布的范圍較廣,而日喀則藏豬則局限在一個較為封閉的環(huán)境中,群體間相距較遠,基因交流的機會較少,外界交流較少,群體內高度近交繁殖,受人工選育程度低,但它們可能具有共同兩個母系起源,在遺傳資源保護過程中應遵循遺傳多樣性原則進行優(yōu)先保護。
[1] 辛盛鵬, 石達, 晉美加措, 等. 西藏自治區(qū)藏豬遺傳資源保護與開發(fā)利用研究[J]. 中國牧業(yè)通訊, 2011(4):49-51.
[2] 強巴央宗, 謝莊, 田發(fā)益. 高原藏豬現狀與保種策略[J]. 中國畜牧雜志, 2001, 37(6):46-47.
[3] Brown WM. Mechanism of evolution in animal mitochondrial DNA[J]. The New York Academy of Sciences, 1981, 361(1):119-134.
[4] Grossi SF, Lui JF, Garcia JE, et al. Genetic diversity in wild(Sus scrofa scrofa)and domestic(Sus scrofa domestica)pigs and their hybrids based on polymorphism of a fragment of the D-loop region in the mitochondrial DNA[J]. Genet Mol Res, 2006, 5(4):564-568.
[5] Royo LJ, Alvarez I, Beja-Pereira A, et al. The origins of Iberian horses assessed via mitochondrial DNA[J]. Journal of Heredity,2005, 96(6):663-669.
[6] 甘佳, 帥素容, 江建平, 李書偉. 四川三個藏豬的微衛(wèi)星遺傳多樣性分析[J]. 四川畜牧獸醫(yī), 2010, 37(12):27-29.
[7] 蔡原, 趙生國. 藏豬遺傳多樣性研究及系統(tǒng)關系研究[J]. 國外畜禽學:豬與禽, 2011(5):60-62.
[8] 楊具田, 臧榮鑫, 盧建雄. 蕨麻豬種質資源保護與開發(fā)利用[J].畜牧與獸醫(yī), 2003, 35(12):15-16.
[9] 孫俊麗, 張冰, 馬青艷, 等. 陸川豬 mtDNA D-loop序列遺傳多樣性分析[J]. 中國畜牧獸醫(yī), 2010, 37(6):122-125.
[10] Qu KX,Wu GS,Gou X,et al. Genetic differentiations between randomly and selectively breed pig populations in Yunnan,China[J]. Zoological Research, 2011, 32(3):255-261.
[11] Li M, Tian S, Jin L, et al. Genomic analyses identify distinct patterns of selection in domesticated pigs and Tibetan wild boars[J]. Nat Genet, 2013, 45(12):1431-1438.
[12] Jiao T, Zhao S, Wang C, et al. Mitochondrial DNA D-Loop diversity of Tibetan Pig populations[J]. The Philippine Agricultural Scientist, 2009, 92(4):362-369.
[13] Yu G, Xiang H, Wang J, et al. The phylogenetic status of typical Chinese native pigs:analyzed by Asian and European pig mitochondrial genome sequences[J]. Journal of Animal Science and Biotechnology, 2013, 4(1):9.
[14] Yang S, Zhang H, Mao H, et al. The local origin of the Tibetan Pig and additional insights into the origin of Asian Pigs[J]. PLoS One, 2011, 6(12):e28215.
[15] 茍瀟, 亐開興, 吳桂生, 等. 云南4個地方豬品種 mtDNA 多態(tài)及其起源分化研究[C]// 中國畜牧獸醫(yī)學會 2004 年學術年會暨第五屆全國畜牧獸醫(yī)青年科技工作者學術研討會論文集(上冊). 北京:中國畜牧獸醫(yī)學會, 2004:237-241.
[16] Sambrook J, Russell DW. Molecular Cloning:A Laboratory Manual[J]. 3rd. New York:Cold SpringHarbor Laboratory Press, 2002:484-485.
[17] 李云南. 鄉(xiāng)城縣藏豬資源的保護及建議[J]. 四川畜牧獸醫(yī),2009(11):8-9.
[18] Jia SG, Chen H, Zhang GX, et al. Genetic variation of mitochondrial D-loop region and evolution analysis in some Chinese cattle breeds[J]. Genet Genomics, 2007, 34:510-518.
[19] Bhuiyan MS, Chen S, Faruque S, et al. Genetic diversity and maternal origin of Bangladeshi chicken[J]. Molecular Biology Reports, 2013, 40(6):4123-4128.
[20] 趙生國. 亞洲部分雞種系統(tǒng)發(fā)育研究及遺傳資源優(yōu)先保護順序評估[D]. 蘭州:甘肅農業(yè)大學, 2009:1-117.
[21] Chen SY, Su YH, Wu SF, et al. Mitochondrial diversity and phylogeographic structure of Chines e domestic goats[J]. Molecular Phylogenetics and Evolution, 2005, 37(3):804-814.
[22] 趙倩君. 中國部分綿羊的起源、遺傳多樣性及保護研究[D].北京:中國農業(yè)科學院, 2007:1-101.
[23] 賴松家. 中國三個牛種遺傳多樣性和分子系統(tǒng)進化研究[D].雅安:四川農業(yè)大學, 2004:1-119.
[24] 雷初朝, 陳宏, 楊公社. 中國部分黃牛品種mtDNA遺傳多態(tài)性研究[J]. 遺傳學報, 2004, 31(1):55-62.
[25] Larson G,Dobney K,Albarella U,et al. Worldwide phylogeography of wild boar reveals multiple centers of pig domestication[J]. Science,2005, 307(5715):1618-1621.
[26] 姚永芳, 徐懷亮, 楊曉軍. 野豬遺傳學研究進展[J]. 經濟動物報, 2007, 11(2):112-115.
[27] Nei M, Tajima F. DNA polymorphism detectable by restruction endonnucleases[J]. Genetics, 1981, 97:145-163.
[28] Neigel JE, Avise JC. Application of a random walk model to geographic distributions of animinal mitochondrial DNA variation[J]. Genetics, 1993, 135(4):1209-1220.
[29] Yang SI, Wang ZG, Liu B, et al. Genetic vari—ation and relationships of eighteen Chinese indigenous pig breeds[J]. Genet Sel Evol, 2003, 35(6):657-667.
[30] 陳國宏, 季從亮, 王敏強, Steffen Weigend. 12個中國地方雞種群體遺傳結構及遺傳多樣性分析[J]. 畜牧獸醫(yī)學報, 2006,37(2):105-111.
[31] 劉益平, 邢光東, 陳仕毅, 等. 蘇種豬和太湖豬的線粒體D環(huán)部分序列比較分析[J]. 江蘇農業(yè)學報, 2007, 23(3):200-203.
[32] Ai H,Huang L,Ren J. Genetic diversity, linkage disequilibrium and selection signatures in Chinese and Western pigs revealed by genome-wide SNP markers[J]. PLoS One, 2013, 8(2):e56001.
[33] FAO. The Global Stratery for the management of farm animal genetic[R]. Rome, Italy, 2007.
[34] Giuffra E, Kijas JMH, Amarger V, et al. The origin of the domestic pig:independent domestication and subsequent introgression[J]. Genetics, 2000, 154:1785-1791.
[35] Watanabe T, HayashiI Y, Kimura J, et al. Pig mitochondrial DNA:polymorphism, restriction map orientation, and sequence data[J]. Biochem Genet, 1986, 24:385-396.
[36] Okumura N, Ishiguro N, Nakano M, et al. Geographic population structure and sequence divergence in the mitochondrial DNA control region of the Japanese wild boar(Sus scrofa leucomystax),with reference to those of domestic pigs[J]. Biochem Genet,1996, 34:179-189.
[37] Wu GS, Yao YG, Qu KX, et al. Population phylogenomic analysis of mitochondrial DNA in wild boars and domestic pigs revealed multiple domestication events in East Asia[J]. Genome Biol,2007, 8:R245.
[38] 劉麗, 趙生國, 蔡原, 等. 早勝牛及其雜交群體遺傳多樣性研究[J]. 農業(yè)生物技術學報, 2014, 22(3):317-325.
[39] Li JY, Luo Z. Research on the habits and characteristics of Tibet pigs on Tibet Plateau[J]. Ecol Domestic Anim, 1993, 14(1):18-21.
(責任編輯 馬鑫)
The Study of Genetic Diversity in Tibetan Pig of Tibet
Guo Yongbo Cai Yuan
(Faculty of Animal Science & Technology,Gansu Agriculture University,Lanzhou730070)
mtDNA D-loop was selected as a marker to determine genetic diversity of the 4 Tibetan pig populations(Nyingchi, Shannan,Chamdo and Shigatse)in Tibet. The results showed A + T content(62.90%)was significantly higher than the G + C content(37.1%),indicating that Tibetan pigs’ mtDNA D-loop was rich in A and T; there was the presence of a base bias. A total of 20 variable sites and 26 haplotypes were identified in the 435 bp nucleotide sequence of mtDNA D-loop, haplotype diversity(Hd)was 0.705 ± 0.021, the average number of nucleotide differences(k)was 1.231, and nucleotide diversity(Pi)was 0.00283. Among them, Hd, k and Pi were the highest in Chamdo Tibetan pig population, and lowest in Shigatse. In addition, Hap1 and Hap3 were 4 shared haplotypes, revealing that there were 2 common maternal ancestor haplotypes in 4 Tibetan pig populations.
Tibetan pig;mtDNA D-loop;haplotype;genetic diversity
10.13560/j.cnki.biotech.bull.1985.2015.05.034
2014-09-10
國家自然科學基金(青年)項目(31101682)
郭永博,男,碩士研究生,研究方向:動物遺傳理論與應用;E-mail:guoyongbo016@163.com
蔡原,女,博士,副教授,研究方向:動物遺傳育種與繁殖;E-mail:caiyuan@gsau.edu.cn
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
