苯磺隆誘導甘藍型油菜中雙2號雄性不育的花藥細胞學觀察
2015-10-20 14:17:53梁魁景等
江蘇農業科學 2015年9期
梁魁景等
摘要:通過花藥形態和細胞學觀察,以測定磺酰脲類除草劑苯磺隆對甘藍型油菜中雙2號育性的影響。結果表明,苯磺隆誘導處理的花藥在形態上發生明顯改變,花藥皺縮、干癟缺水、顏色淡黃、纖維絲薄短、緊連蜜腺管底端、不形成花粉粒等;絨氈層的破壞是導致甘藍型油菜雄性不育的主要原因。
關鍵詞:苯磺隆;油菜;花藥;細胞學觀察
中圖分類號:S565.401文獻標志碼: A文章編號:1002-1302(2015)09-0128-02
油菜是人們日常生活所需食用油和調味料的重要資源,還可作為一種蔬菜。利用雜種優勢是提升農作物產量和價值的重要技術[1-2],而油菜在產量和一些農藝性狀方面具有顯著的雜種優勢。阻止母本自我受精雖然降低了農作物種子的純度,但對雜種的獲得具有重要的意義。開發利用雄性不育親本是確保母本異型雜交和大規模獲得F1代雜交種子的重要途徑。張寶娟等研究表明,通過葉面噴施15~20 mL磺酰脲類除草劑苯磺隆(TM)2次,能誘導甘藍型油菜中雙9號完全不育,且高效穩定,是一種較好的油菜化學殺雄劑[3]。本研究通過苯磺隆誘導油菜品種中雙2號,對其花藥進行形態和細胞學觀察,為揭示化學藥物誘導油菜雄性不育的機制提供形態和細胞學依據。
1材料與方法
1.1材料
適合于河北省種植的冬性甘藍型油菜品種中雙2號,由中國農業科學院油料作物研究所提供;化學殺雄劑75%苯磺隆水分散粒劑,由江蘇江南農化有限公司生產。
1.2處理方法
2013年9月初,將油菜定植在大田,株行距為0.1 m×0.5 m;11月中旬,將油菜從大田移植到溫室,溫室條件:溫度為16~23 ℃,濕度為65~75%,光照16 h左右;在油菜抽薹高度為15~20 cm、最大花蕾長度為1~2 mm時,按苯磺隆最佳用藥量15~20 mL/單株,采用小型手動噴霧器均勻噴施于油菜葉片表面,以噴清水為對照。
1.3細胞學觀察
采用石蠟切片法對花藥進行細胞學觀察。石蠟切片法流程采用李正理的方法[4],將油菜穩定的可育和不育性花序在雙蒸水 ∶無水乙醇 ∶醋酸為6 ∶3 ∶1的改良卡諾固定液中固定24 h;分別用30%、50%、70%、80%、95%、100%乙醇從低濃度到高濃度進行梯度脫水;用二甲苯進行透明,浸56~58 ℃石蠟,包埋;對石蠟塊進行固著和整修,用Leica EM UC7超薄切片機進行切片,切片厚度為1 μm;明膠黏貼劑黏片,37 ℃烘干超過24 h;蘇木精染色,阿拉伯樹膠封片,Olympus BX51顯微鏡觀察,獲得70 nm超微結構,用JEM-1230型透射電子顯微鏡進行拍照。
2結果與分析
2.1形態觀察
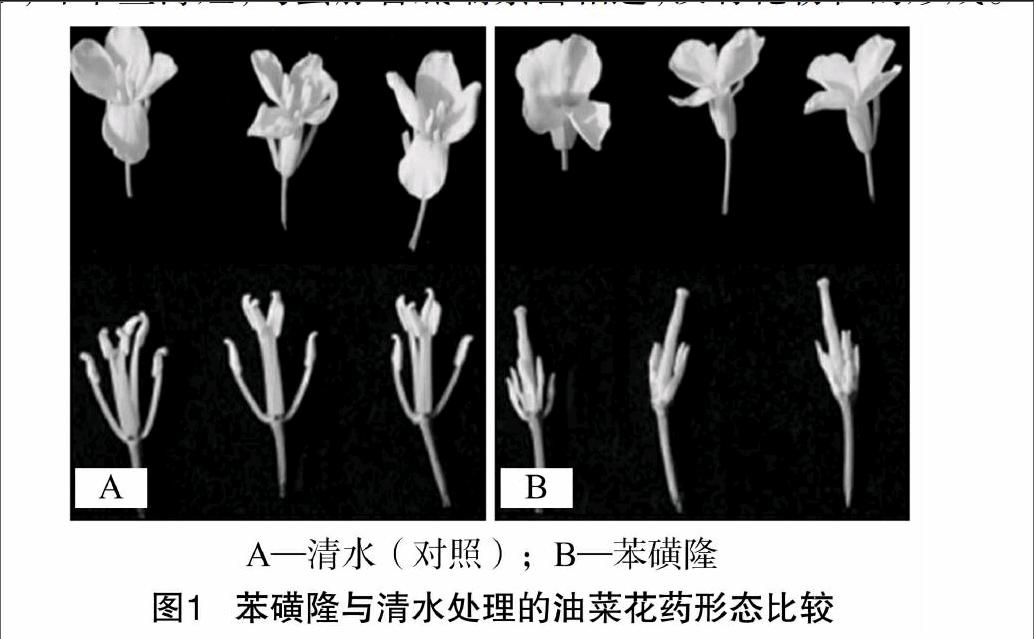
由圖1可見,正常發育的油菜花藥(噴清水)生長良好,長而豐滿,花粉粒數量眾多;苯磺隆誘導處理的油菜,雌蕊顯著長于雄蕊,花藥發育較差,呈淡黃色,皺縮,呈干燥的三角形,纖維絲薄短,與蜜腺管底端緊密相連,沒有花粉粒的形成。
2.2細胞學觀察
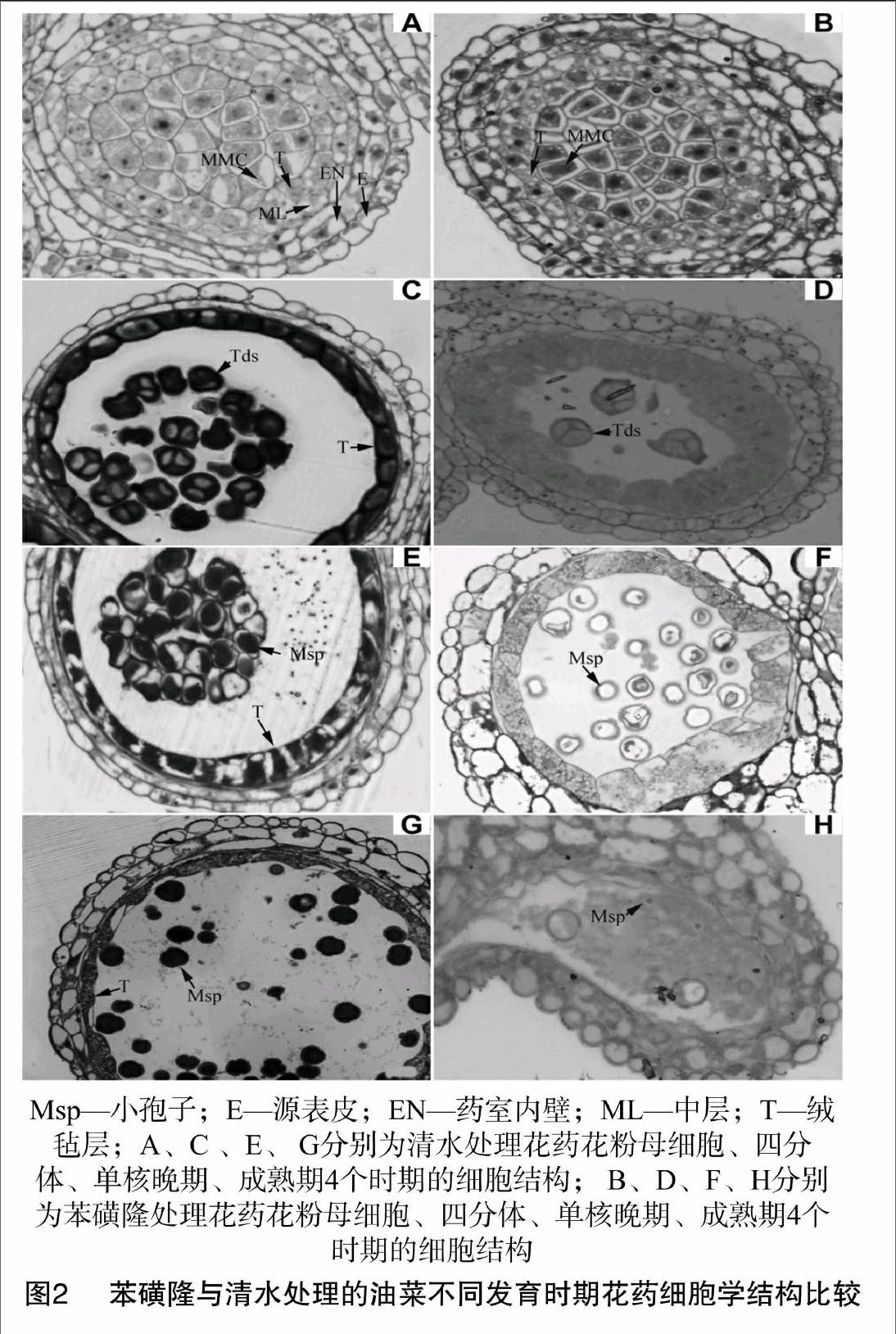
在花粉母細胞、四分體、單核細胞、成熟期4個發育時期,對中雙2號油菜花藥的細胞結構進行觀察。由圖2可見,在花粉母細胞時期,噴清水處理的油菜花藥,細胞結構包括1層源表皮、藥室內壁、中層及絨氈層,細胞質的染色較均勻(圖2-A);苯磺隆誘導藥物處理的油菜花藥,細胞源表皮、藥室內壁、中層細胞沒有發生明顯變化,花粉母細胞的數量和形態也未發生改變,絨氈層雖然保持完整性,但有大量藍色物質聚集在絨氈層細胞中央,周圍形成空泡結構(圖2-B),與清水處理的花藥有明顯差別;在四分體時期,苯磺隆處理的油菜花藥絨氈層細胞遭到不同程度破壞,與對照(圖2-C)相比,其花藥雖然包括有完整的絨氈層,但細胞質聚合成塊,周圍形成空泡狀,細胞數量和排布密度都有很大程度降低(圖2-D);在單核期,清水處理的花藥小孢子出現空泡化,絨氈層也發生退化,這是單核晚期的表現(圖2-E),而苯磺隆誘導處理的花藥,其細胞絨氈層發生異常,大小不一,退化延遲,且無細胞核,小孢子細胞發生畸變而無活性(圖2-F);在成熟期,清水處理的花藥中已經形成許多去空泡化、可育的小孢子,絨氈層還存在,但已經發生退化(圖2-G),而苯磺隆處理的花藥絨氈層已經消失不見,小孢子數量很少,即使形成小孢子,也呈空泡化狀,呈現敗育性狀(圖2-H)。
3結論
苯磺隆屬于乙酰乳酸合酶抑制劑,是一類非常重要的磺酰脲類除草劑[5],目前對其雜草清除的生化和分子機制已經進行了深入研究[6]。苯磺隆還是一種可高效誘導甘藍型油菜完全雄性不育的化學殺雄劑[3,7-8],但當前對其誘導植物雄性不育的細胞學機制還不是十分明確。
通過光學顯微鏡和透射電子顯微鏡觀察分析苯磺隆誘導處理的油菜花藥細胞學形態發現,在花粉母細胞、四分體、單核細胞、成熟期4個發育時期,都能觀察到花藥細胞有缺陷的絨氈層,花粉母細胞時期絨氈層細胞的細胞質濃縮聚集,形成空泡狀結構,絨氈層細胞在單核期退化延遲。除此之外,在苯磺隆誘導處理的花藥中,還觀察到一些畸變的小孢子,最終導致成熟期花粉的敗育,與Cheng等研究結論[9-10]較為相似。這可能是因為苯磺隆誘導處理油菜花藥,使各個時期絨氈層的細胞發育異常,包括提早分裂、絨氈層與中間層剝離、延遲降解等,不能提供正常花粉發育所需的成分,形成敗育的小孢子,從而誘導油菜雄性不育,這種化學藥物誘導的油菜雄性不育,其花藥細胞學特性不同于受特定育性相關基因調控的細胞質雄性不育和細胞核雄性不育[11-12]。
參考文獻:
[1]李培武,丁小霞,張文,等. 甘藍型油菜葉片硫甙含量的雜種優勢研究[C]//湖北省植物生理學會第十五次學術研討會論文集,2007:63-68.
[2]傅廷棟. 雜交油菜的育種與利用[M]. 武漢:湖北科學技術出版社,1995.
[3]張寶娟,趙惠賢,胡勝武. 苯磺隆對甘藍型油菜中雙9號的殺雄效果[J]. 中國油料作物學報,2010,32(4):467-471.
[4]李正理. 植物制片技術[M]. 2版.北京:科學出版社,1987:90-92.
[5]Zhou Q Y,Liu W P,Zhang Y S,et al. Action mechanisms of acetolactate synthase-inhibiting herbicides[J]. Pesticide Biochemistry and Physiology,2007,89(2):89-96.
[6]Das M,Reichman J R,Haberer G,et al. A composite transcriptional signature differentiates responses towards closely related herbicides in Arabidopsis thaliana and Brassica napus[J]. Plant Molecular Biology,2010,72(4/5):545-556.
[7]Yu C,Hu S,He P,et al. Inducing male sterility in Brassica napus L. by a sulphonylurea herbicide,tribenuron-methyl[J]. Plant Breeding,2006,125(1):61-64.
[8]胡海兵. 化學除草劑誘導甘藍型油菜雄性不育效果與生理機制研究[D]. 雅安:四川農業大學,2012:14-27.
[9]Cheng Y,Wang Q,Li Z,et al. Cytological and comparative proteomic analyses on male sterility in Brassica napus L. induced by the chemical hybridization agent monosulphuron ester sodium[J]. PLoS One,2013,8(11):e80191.
[10]井苗,董振生,嚴自斌,等. BHL等4種藥物對油菜殺雄效果的研究[J]. 西北農業學報,2008,17(3):165-170.
[11]González-Melendi P,Uyttewaal M,Morcillo C N,et al. A light and electron microscopy analysis of the events leading to male sterility in Ogu-INRA CMS of rapeseed (Brassica napus)[J]. Journal of Experimental Botany,2008,59(4):827-838.
[12]Xiao Z D,Xin X Y,Chen H Y,et al. Cytological investigation of anther development in DGMS line Shaan-GMS in Brassica napus L.[J]. Czech Journal of Genetics and Plant Breeding,2013,49(1):16-23.
