細胞間隙連接蛋白43基因在斑馬魚牙齒早期發育中的表達
2015-10-13 08:15:40徐智云劉鑫黃興楊德琴
華西口腔醫學雜志 2015年4期
關鍵詞:信號
徐智云 劉鑫 黃興 楊德琴
重慶醫科大學附屬口腔醫院牙體牙髓科·口腔疾病與生物醫學重慶市重點實驗室,重慶 400017
牙齒發育是一種復雜的生理過程,主要通過上皮和間充質間的相互作用完成,期間涉及多種生長因子和信號途徑,在特定的時空調控下共同作用最終成形。細胞間隙連接蛋白43(connexin 43,cx43)直接調控細胞間的連接通訊,進行信息和能量的傳遞,進而調控細胞的新陳代謝和增殖分化等一系列生理過程[1],在脊椎動物早期的心血管發育[2]、神經系統傳導[3]、骨骼形成和礦化[4]中發揮重要作用,同時與心血管系統、泌尿生殖系統、神經系統等疾病的發生發展密切相關[5-6]。研究[7]發現,cx43基因在小鼠牙齒發育成形過程中高表達,參與牙齒發育,并且在牙齒礦化過程中發揮重要作用。cx43基因缺陷與人類常染色體顯性遺傳病眼-牙-指綜合征(oculoden-todigital dysplasia,ODDD)相關,呈現并指、釉質發育不全和(或)顱面部畸形等表型[8]。目前cx43基因在牙齒發育和礦化過程中的具體功能尚不清楚。
斑馬魚是一種公認的用于研究器官形成和發育相關分子生物學機制的理想模式生物,因其同哺乳動物高度的基因同源性而被廣泛運用于發育生物學的研究[9]。斑馬魚牙齒的發育和替換很大程度上反映了有牙脊椎動物的原始狀態[10]。本研究旨在探索cx43基因在斑馬魚牙齒發育和形成中的作用,為探究牙齒內源性再生奠定基礎。
1 材料和方法
1.1 實驗動物
野生型(wild type)斑馬魚,由西南大學分子發育實驗室購進。
1.2 主要試劑及設備
聚合酶鏈反應(polymerase chain reaction,PCR)引物由上海生工生物工程技術服務有限公司合成。限制性內切酶(NEB公司,美國),PCR puri fi cation Kit檢測試劑盒(Qiagen公司,美國),逆轉錄試劑盒(Roche公司,美國),RNA提取試劑盒(Life Technologies公司,美國),T4連接酶(Invitrogen公司,美國)。體視解剖顯微鏡(Leica公司,德國),臺式離心機(Eppendorf公司,德國)。
1.3 動物飼養
參照Wester fi eld[11]的方法于28.5 ℃水溫、14 h光照、10 h黑暗的條件下進行飼養,按雌雄比例1︰1或1︰2放入交配缸內,次日清晨收集斑馬魚受精卵并清洗,置于28.5 ℃恒溫孵育箱中培養和胚胎孵育。用于整胚原位雜交的胚胎培養液中需添加0.04%的1-苯基-2-硫脲(phenylthiourea,PTU)孵育。
1.4 cx43反義探針的制備
收集野生型斑馬魚受精后72 h的胚胎,用Trizol法提取總RNA,運用瓊脂糖凝膠電泳及紫外分光光度計檢測完整性及純度,逆轉錄合成cDNA,-20 ℃儲存備用。根據斑馬魚cDNA文庫中cx43基因序列設計引物,探針正向引物:5’-ACTCCCTCAAGAXXXXXGACT-3’,反向引物:5’-CTAGCGTTGGGXXXXXGCAT-3’,以cDNA為模板進行PCR擴增。連接PCR產物于PGEMT載體上,轉化到DH5α感受態菌株中進行克隆,篩選氨芐抗性陽性質粒。用EcoRⅠ酶或NotⅠ酶對重組質粒進行酶切驗證,通過測序鑒定判斷插入片段連接方向。取正確的重組質粒單一酶切位點線性化,電泳鑒定線性化徹底后回收。電泳判斷濃度,用SP6或T7 RNA聚合酶體外轉錄體系轉錄模板,同時加入地高辛標記的寡核苷酸,轉錄后用DnaseⅠ酶消化多余的DNA骨架,產物經LiCl純化后得到地高辛標記的cx43基因反義mRNA探針,1%瓊脂糖凝膠電泳鑒定RNA質量并測量濃度,-20 ℃保存。
1.5 斑馬魚全時相胚胎原位雜交
運用整胚原位雜交技術檢測斑馬魚胚胎發育早期cx43基因在牙齒發育部位的表達分布。收集受精后48 h、60 h、72 h、84 h、96 h、108 h、120 h、132 h、6 d、7 d、8 d、9 d不同發育時期的斑馬魚胚胎,4%多聚甲醛4 ℃過夜固定,甲醇梯度脫水至胚胎透明,-20 ℃、100%甲醇溶液中保存備用。第1天,1×PBST梯度孵育復水,用5 μg·mL-1蛋白酶K消化[12]并4%多聚甲醛再固定后移入68 ℃雜交爐中預雜交2~5 h,地高辛標記的cx43 RNA探針預變性處理后,加入胚胎中65 ℃雜交過夜。第2天,首先2×SSCT梯度逐步洗去多余非特異性結合探針,然后室溫下梯度過度到馬來酸緩沖液(Maleic acid buffer,MABT),用1×封閉緩沖液(1×blocking)胚胎封閉2 h。最后加入1︰2 000稀釋的anti-DIG-AP封閉液與cx43基因反義mRNA探針在4 ℃下結合過夜。第3天,室溫下用MABT清洗直至洗凈,嚴格避光,NTMT平衡液中按1︰50加入顯色液5-溴-4-氯-3-吲哚基-磷酸(5-bromo-4-chloro-3-indolyl phosphate,BCIP)/硝基四氮唑藍(nitrotetrazolium blue chloride,NBT)溶液顯色20 min,顯微鏡下觀察顯色成功(出現棕褐色雜交信號)后終止染色,記錄結果并進行分析。
1.6 茜素紅染色
取受精后9 d斑馬魚適量,放置于MS-222溶液(100 mg·L-1)中麻醉致死,用4%多聚甲醛固定2 h后50%乙醇脫水10 min;去除乙醇加入0.5%茜素紅染液染色過夜;純水漂洗后用現配的1.5%雙氧水和1%氫氧化鉀混合液漂白20 min,經0.5%KOH和甘油混合液梯度透明,保存于純甘油中[13],比較cx43原位雜交表達區域與咽齒解剖部位的關系。
2 結果
2.1 cx43反義探針的制備
通過PCR擴增獲得的特異性條帶,片段大小約1 179 bp(圖1),長度符合預期。將PCR片段連接于PGEMT載體上,對重組質粒用EcoRⅠ酶切驗證(圖2),正確的重組質粒可在EcoRⅠ酶的作用下切出3 kb骨架和1 179 bp插入片段。7個待驗證質粒中除1個外余皆可切出相應片段,為正確的重組質粒。質粒PGEMT-cx43構建成功。取正確的重組質粒,判斷片段連接方向為反向,用SalⅠ在SP6方向切開線性化重組質粒,選用T7 RNA聚合酶反轉錄獲得cx43基因反義mRNA探針。
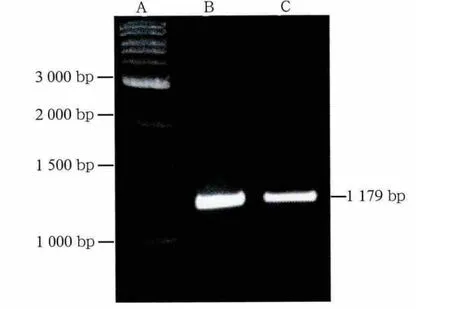
圖1 PCR結果Fig 1 PCR results
2.2 茜素紅染色結果
茜素紅染色結果表明,受精后9 d時cx43原位雜交表達部位與咽齒解剖部位基本重疊一致(圖3),說明cx43基因表達于發育中的咽齒,而對已經礦化完全的咽齒不發揮作用。
2.3 斑馬魚全時相胚胎原位雜交結果
原位雜交結果表明,cx43基因在斑馬魚受精后各個時期咽弓牙齒相應部位均可見到強棕褐色陽性信號,除此以外中樞神經系統、頭部、眼睛以及胰腺等部位也可檢測到陽性信號(圖4)。
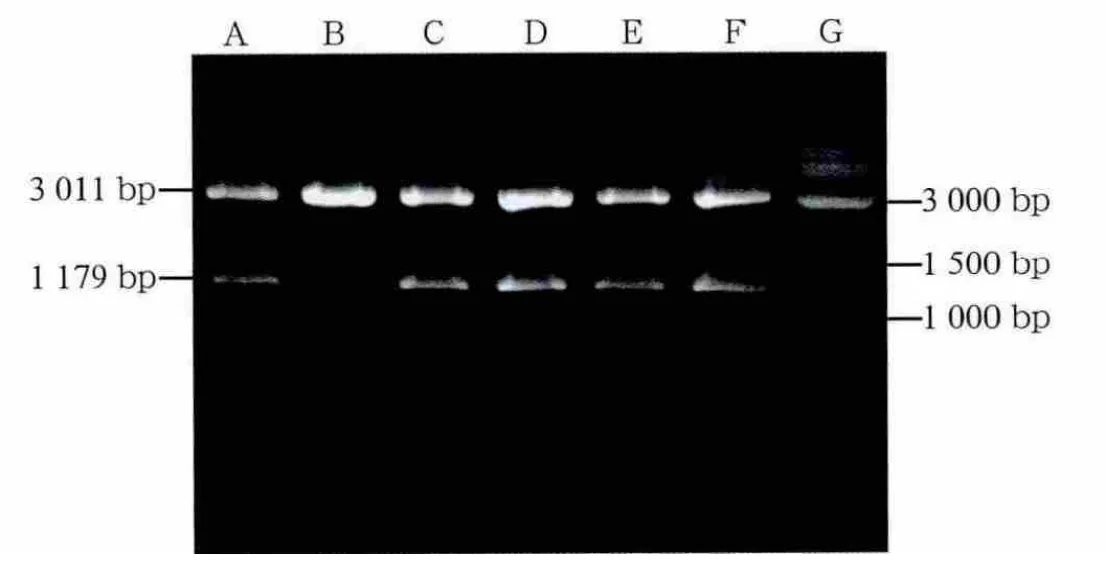
圖2 重組質粒EcoRⅠ酶切驗證Fig 2 Restriction enzyme digestion of recombination plasmid with EcoRⅠ
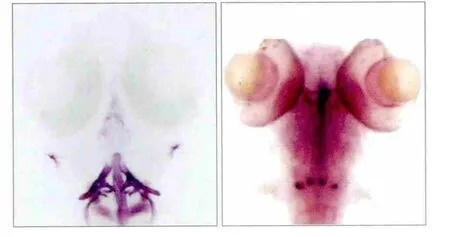
圖3 cx43基因表達部位與咽齒解剖部位 茜素紅染色 × 50Fig 3 cx43 gene expression and anatomical of pharyngeal teeth ali-zarin red stain × 50
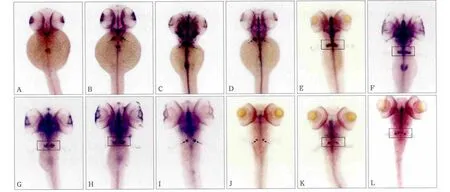
圖4 cx43基因48 h~9 d的斑馬魚胚胎 原位雜交 × 50Fig 4 cx43 gene from 48 h to 9 d embryo of Zebra fi sh in situ hybridization × 50
在斑馬魚牙齒發育起始點受精后48 h,cx43少量表達于眼睛、前腎以及神經中樞(圖4A);60 h時檢測到cx43在牙齒相應部位開始表達,同時神經中樞表達增強,背側觀棕褐色信號以中線為軸左右對稱,位于頭部稍下最后咽弓處(圖4B),左側位觀表達位于心臟斜上方,部位與咽齒的解剖部位相重疊。72 h時可見cx43信號表達量較之前明顯上升,背側觀信號增強明顯(圖4C);84 h時出現回落,表達量逐漸降低(圖4D);96 h時開始一側由一整個信號點逐漸分為2個,部分兩信號間區分不明顯(圖4E);108 h單側2個等大信號較前一時相更為獨立清晰(圖4F);120 h時單側信號點由2個向3個轉換(圖4G);132 h時呈現單側3個清晰的信號點且左右表型不對稱(圖4H);6 d時表達量明顯減弱(圖4I);7 d時維持一個較弱的表達(圖4J);8 d時表達略增強,呈中間大兩邊小3個點(圖4K)或一大一小2個點;9 d時單側出現兩個等大的弱信號(圖4L)。
3 討論
目前國內外學者多以小鼠、小型豬等作為研究牙發育的首選,然而這些模式生物由于自身的局限性,對于基因表達的動態追蹤定位、信號間相互傳導的級聯網絡分析等相關研究存在極大困難,不能夠準確獲得基因表達時間、空間與具體發育過程之間的相互聯系。斑馬魚雖為原始脊椎動物,但與人類具有高度的基因同源性和相似的分子機制,具有研究的前瞻價值[14]。斑馬魚的優勢除了體積小、易于飼養與繁殖、成熟周期短,能夠快速獲得大量個體以外,同時具有體外發育、早期胚體透明等特點,借助一些特殊的實驗技術和實驗手段,易于追蹤觀察,獲取直觀信息,為研究分子機制提供了可操作性。這是本研究選用斑馬魚為模式生物的原因。
斑馬魚缺乏口齒但存在發育礦化良好的咽齒,咽齒局限于第五鰓弓處,其牙為端生牙、同形牙、多牙列。咽齒無論在基本組成結構還是發育過程上都與哺乳動物具有極強的相似性[15]。礦化程度和Ca、P比例亦與哺乳動物具有一定程度的相似性[16]。咽齒的特異性在于存在部位特殊和缺乏牙根[17],但這不影響早期牙胚發育過程的研究。斑馬魚是研究人類牙齒形態特征、組織結構和生物礦化過程的理想模式動物。斑馬魚牙齒分為腹(V)、中(MD)、背(D)側3組,最早發育的牙齒4V1在胚胎受精后48 h即開始形成,在4 d時完全礦化及附著于牙槽骨上[18]。3V1和5V1在受精后56 h時同時出現在4V1兩側,并在6 d時發育完善及附著于牙槽骨上[20]。最早替換的牙齒4V2形成于受精后80 h,3V2、5V2隨后出現于144 h。4V2、3V2和5V2在受精后12 d完全發育及附著于牙槽骨上[19]。
本研究整胚原位雜交結果表明,cx43在斑馬魚牙齒發育過程中發揮作用。初代牙齒期:雖然4V1在受精后48 h開始形成,但cx43無表達,此時牙處于鐘狀期尚未礦化。60 h時4V1進入晚期細胞分化期,cx43基因參與其中并開始表達于牙齒發育部位。72 h時3V1和5V1由早期細胞向晚期細胞分化,cx43表達量上升迅速,84 h時可見cx43表達量有所減低。由此可推測cx43在細胞分化晚期至附著初期發揮作用,即在礦化階段發揮功能。3V1、5V1從72 h到120 h一直處于晚期細胞分化階段,從96 h開始隨著3V1、5V1發育的進行,單側逐漸出現兩個高表達點(3V1,5V1),兩個表達信號之間起初較為接近,至108 h時逐漸獨立清晰,表達信號左右對稱等大。發育替換期:最早的替換牙齒4V2形成于80 h,而3V2、5V2形成于6 d。既往的研究發現,斑馬魚替換牙齒與初代牙齒發育分子信號機制存在差異,發育時間有所延長。初代牙齒一般2~4 d即可完成發育,而替換的牙齒發育時間多延長到8 d左右[20]。因此可以推論,替換的牙齒發育礦化的時間也相應推后。從cx43在相應部位的表達檢測可以發現,120 h時4V2開始礦化,此時兩個信號點(3V1、5V1)向3個信號點(4V2、3V1、5V1)轉化,132 h時基本兩側呈現3個信號點(4V2、3V1、5V1)且兩邊稍大,中間略小。6 d時3V1、5V1礦化完成,cx43相應部位表達結束,原位雜交結果符合預期。6、7 d時cx43表達呈單側單一信號(4V2),8 d時4V2、3V2及5V2同時處于細胞分化礦化期,呈現中間大兩邊小3個信號點(4V2、3V2、5V2)。9 d時4V2礦化完成,3V2、5V2繼續直至12 d完成附著。cx43表達模式同牙齒礦化階段吻合,且主要表達于礦化晚期。這可初步認為cx43在牙齒細胞分化晚期發揮作用,參與礦化。此外從cx43信號的表達量上,亦可以看出初代牙齒與替換牙齒之間存在差異。這種差異可能與干細胞在初代牙齒形成過程中的集聚現象有關。
關于cx43在牙齒發育中的作用,最早Pinero等[21]用受精后1 d的小鼠進行免疫組織化學染色方法發現cx43在新生鼠的口腔成釉上皮層、成牙本質細胞層及牙槽骨處均有表達,且表達廣泛存在于內釉上皮、中間層和成牙本質細胞區域。這說明cx43與牙的形成相關。Jo?o等[7]運用激光共聚焦技術發現cx43基因參與成釉細胞和成牙本質細胞的早期分化。Toth等[22]運用ODDD小鼠模型證明了cx43表達水平的降低是成釉細胞發育失常、釉質發育不全的主要原因。這說明cx43同時參與了釉質的形成。本實驗運用斑馬魚整胚原位雜交技術對基因表達部位進行連續動態檢測,證實了cx43參與牙齒的礦化過程,并且主要作用于礦化晚期階段。
牙的發育和形成過程涉及多種基因共同參與形成復雜的網絡調節系統,根據原位雜交數據可發現,cx43在斑馬魚牙齒發生部位和礦化階段高表達,說明cx43參與牙齒發育礦化的過程,然而cx43基因調控牙齒發育的機制及與牙本質細胞礦化相關性的研究尚屬探索階段,因此本實驗運用斑馬魚為模式生物,通過原位雜交技術探究cx43基因在斑馬魚早期胚胎發育過程中的表達,結果表明cx43基因參與了牙齒的發育和分化,且主要作用于礦化晚期階段。
[1]Bloor DJ, Wilson Y, Kibschull M, et al. Expression of connexins in human preimplantation embryos in vitro[J]. Reprod Biol Endocrinol, 2004, 2:25.
[2]Song YN, Zhang H, Zhao JY, et al. Connexin 43, a new therapeutic target for cardiovascular diseases[J]. Pharmazie,2009, 64(5):291-295.
[3]Salmina AB, Morgun AV, Kuvacheva NV, et al. Establishment of neurogenic microenvironment in the neurovascular unit: the connexin 43 story[J]. Rev Neurosci, 2014, 25(1):97-111.
[4]Plotkin LI, Bellido T. Beyond gap junctions: connexin43 and bone cell signaling[J]. Bone, 2013, 52(1):157-166.
[5]Nishii K, Shibata Y, Kobayashi Y. Connexin mutant embryonic stem cells and human diseases[J]. World J Stem Cells, 2014, 6(5):571-578.
[6]林山力, 文歡, 鄧紅. 連接子蛋白43在腫瘤中的作用及其機制的研究進展[J]. 中華病理學雜志, 2014, 43(1):62-64.
[7]Jo?o SM, Arana-Chavez VE. Expression of connexin 43 and ZO-1 in differentiating ameloblasts and odontoblasts from rat molar tooth germs[J]. Histochem Cell Biol, 2003,119(1):21-26.
[8]Laird DW. Syndromic and non-syndromic disease-linked Cx43 mutations[J]. FEBS Lett, 2014, 588(8):1339-1348.
[9]Verstraeten B, van Hengel J, Sanders E, et al. N-cadherin is required for cytodifferentiation during zebra fi sh odontogenesis[J]. J Dent Res, 2013, 92(4):365-370.
[10]Stock DW. Zebra fi sh dentition in comparative context[J].J Exp Zool B Mol Dev Evol, 2007, 308(5):523-549.
[11]Wester fi eld M. The zebra fi sh book. A guide for the laboratory use of zebra fi sh (Danio Rerio)[M]. 5th ed. Eugene:University of Oregon Press, 2007:231-236.
[12]Verstraeten B, Sanders E, Huysseune A. Whole mount immunohistochemistry and in situ hybridization of larval and adult zebra fi sh dental tissues[J]. Methods Mol Biol, 2012,887:179-191.
[13]Walker MB, Kimmel CB. A two-color acid-free cartilage and bone stain for zebra fi sh larvae[J]. Biotech Histochem,2007, 82(1):23-28.
[14]Tsang M. Zebra fi sh: a tool for chemical screens[J]. Birth Defects Res C Embryo Today, 2010, 90(3):185-192.
[15]Jernvall J, Thesleff I. Tooth shape formation and tooth renewal: evolving with the same signals[J]. Development,2012, 139(19):3487-3497.
[16]Arnold WH, Naumova KI, Naumova EA, et al. Comparative qualitative and quantitative assessment of biomineralization of tooth development in man and zebra fi sh (Danio rerio)[J].Anat Rec: Hoboken, 2008, 291(5):571-576.
[17]Huysseune A, Thesleff I. Continuous tooth replacement: the possible involvement of epithelial stem cells[J]. Bioessays,2004, 26(6):665-671.
[18]Jackman WR, Stock DW. Transgenic analysis of Dlx regulation in fi sh tooth development reveals evolutionary retention of enhancer function despite organ loss[J]. Proc Natl Acad Sci USA, 2006, 103(51):19390-19395.
[19]Borday-Birraux V, Van der Heyden C, Debiais-Thibaud M, et al. Expression of Dlx genes during the development of the zebra fi sh pharyngeal dentition: evolutionary implications[J]. Evol Dev, 2006, 8(2):130-141.
[20]Van der Heyden C, Huysseune A. Dynamics of tooth formation and replacement in the zebra fi sh (Danio rerio)(Teleostei, Cyprinidae)[J]. Dev Dyn, 2000, 219(4):486-496.
[21]Pinero GJ, Parker S, Rundus V, et al. Immunolocalization of connexin 43 in the tooth germ of the neonatal rat[J]. Histochem J, 1994, 26(10):765-770.
[22]Toth K, Shao Q, Lorentz R, et al. Decreased levels of Cx43 gap junctions result in ameloblast dysregulation and enamel hypoplasia in Gja1Jrt/+ mice[J]. J Cell Physiol, 2010, 223(3):601-609.
猜你喜歡
鴨綠江(2021年35期)2021-04-19 12:24:18
考試與評價·高一版(2020年6期)2020-11-02 02:45:24
媽媽寶寶(2019年10期)2019-10-26 02:45:34
中國生殖健康(2019年3期)2019-02-01 06:12:26
鐵道通信信號(2018年11期)2019-01-19 01:15:08
電子制作(2018年11期)2018-08-04 03:25:42
鐵道通信信號(2018年2期)2018-04-18 12:18:10
鐵道通信信號(2016年11期)2016-06-01 12:11:32
鑿巖機械氣動工具(2016年3期)2016-03-01 04:00:25
中國病理生理雜志(2015年8期)2015-12-21 12:38:06
