骨形態發生蛋白受體2在維甲酸誘導腭裂小鼠胚胎腭突中的表達
2015-10-13 08:15:46陳沐劉學余東升王成王偉財黃洪章
華西口腔醫學雜志 2015年4期
關鍵詞:小鼠
陳沐 劉學 余東升 王成 王偉財 黃洪章
1.廣東醫學院附屬南山醫院口腔科,深圳 518052;2.中山大學光華口腔醫學院·附屬口腔醫院口腔頜面外科,中山大學口腔醫學研究所,廣州 510055
先天性唇腭裂是一類常見的口腔頜面部缺陷,占所有先天性畸形的65%左右。目前我國非綜合征性唇腭裂的發病率為0.762‰,其中非綜合征性腭裂的發病率為0.2‰[1];中國出生缺陷監測網數據顯示,腭裂年度發病率呈上升趨勢[2];因此腭裂是目前出生缺陷研究中的焦點問題之一[3-4]。已有研究[5]表明,妊娠期攝入致畸物可能導致新生兒腭裂的發生。在致畸物的研究中,維生素A族與唇腭裂發病的關系一直受到唇腭裂病因研究學者的高度關注[6]。全反式維甲酸(all-trans retinoic acid,atRA)是維生素A在體內的代謝中間產物及發揮功能的主要形式,參與調節早期胚胎細胞的定向分化,在胚胎發生、發育中起重要作用[7]。在哺乳動物胚胎發育過程中,過多攝入維甲酸可導致神經管、心臟和顏面部等多種畸形的發生,形成唇腭裂、面中部發育不良、顱中縫早閉等一系列的頜面部骨骼發育畸形,但其作用機制目前仍未十分明確[8-9]。
有學者[10-11]認為,上頜骨的生長發育障礙是腭裂畸形本身內在的發育缺陷所致。根據國內外的研究[12-15]可以發現,腭裂可因胚胎發育時期的胚胎細胞外基質及細胞的分化、移動、增殖受到干擾而引起;一些致畸劑通過阻斷某些基因的表達或干擾某些物質(如表皮生長因子受體、轉化生長因子β3等)的合成而影響胚胎細胞的分化移動。當裂隙形成后,這些內源性因素就成為影響上頜骨發育的主要原因。研究[16]指出,骨形態發生蛋白(bone morphogenetic protein,BMP)信號通路在唇及腭部的發育中起著非常重要的調節作用,然而BMP信號通路在atRA誘導過程中的表達模式及其對小鼠腭突生長發育影響的調控作用目前尚不清楚。本實驗采用孕鼠妊娠第10天時atRA灌胃的方法,用免疫組織化學及逆轉錄聚合酶鏈式反應(reverse transcription-polymerase chain reaction,RT-PCR)技術檢測BMP信號通路關鍵因子骨形態發生蛋白受體(bone morphogenetic protein receptor,BMPR)2的時空表達,為后期研究維甲酸對腭部間充質細胞成骨分化進程的影響建立理論及實驗基礎。
1 材料和方法
1.1 實驗材料
無特定病原體(spece fi c pathogen free,SPF)級6~10周齡未生育C57BL/6N小鼠(購自南方醫科大學實驗動物中心);atRA(Sigma-Aldrich公司,美國,批號R2625);BMPR2抗體(Santa Cruz公司,美國);SP免疫組織化學試劑盒、DAB顯色試劑盒(北京中杉金橋生物技術有限公司)。
1.2 實驗方法
將C57BL/6N小鼠于上午8時按雌雄比例1∶1合籠,2 h后檢查陰栓,查見陰栓日定為妊娠(gestation day,GD)0 d(GD0)。將atRA溶解于玉米油中配制成10 g·L-1溶液,以純玉米油作為對照。在GD10隨機將妊娠母鼠分為兩組,實驗組按照100 mg·kg-1劑量給予atRA灌胃1次,對照組給予相同劑量的玉米油灌胃。
1.3 蘇木精-伊紅(haematoxylin-eosin,HE)染色
兩組動物分別在GD15、GD17時隨機選取3只,以頸椎脫位法處死,剖腹取出所有胚胎。每只孕鼠的胚胎仔鼠中隨機選取5只,在體視顯微鏡下用顯微外科器械從口裂至耳屏前水平切開,以顯微鑷去除下頜、舌體及后腦組織,充分暴露胎鼠腭突。取材組織固定、包埋并進行HE染色。
1.4 免疫組織化學染色
免疫組織化學染色方法和操作步驟按試劑盒說明書進行。切片脫蠟至水,封閉通透液浸潤,檸檬酸修復抗原,5%血清封閉非特異性蛋白,一抗(1∶100)濕盒孵育4 ℃過夜;次日復溫,PBS洗后加入稀釋的二抗烤片;PBS洗后滴加DAB溶液顯色,蘇木素染液復染,鹽酸乙醇藍化,中性樹膠封固,光學顯微鏡下觀察。在高倍鏡下隨機選取小鼠胚胎腭突部5個視野觀察100個組織細胞,計算陽性細胞所占的百分比為陽性指數,取其均值計為BMPR2的表達強度。
1.5 RT-PCR檢測
在GD15和GD17時按照步驟1.3的方法取兩組孕鼠的胚胎,分離胚胎腭突并取材,冷PBS清洗,取50~100 mg組織塊,用液氮在研缽中研磨成粉末。參照Trizol試劑說明書提取胚胎腭突內總RNA。按照Thermo ScriptRTSystem試劑盒說明書要求進行RTPCR反應,引物序列如表1所示。RT-PCR產物進行瓊脂糖凝膠電泳,結果采用凝膠成像系統照相,測定每一條帶的灰度值。目的基因mRNA的表達以目的基因灰度值/內參β-actin灰度值表示。將目的基因mNRA表達比值進行統計學分析。
1.6 統計學分析
所得數據輸入SPSS 19.0統計學軟件進行統計學分析,對樣本檢測數據進行組間方差分析,檢驗水準為雙側α=0.05。

表1 RT-PCR引物Tab 1 Primers of RT-PCR
2 結果
HE染色觀察胚胎腭突的發育情況見圖1,免疫組織化學檢測胚胎腭部BMPR2的表達情況見圖2。通過HE染色可見,GD15時,對照組兩側腭突在中線處已經接觸融合,腭中縫上皮消失,兩側胚胎腭間充質組織相互交雜,間充質細胞核呈梭形,向腭突中心部位遷移生長,中線處有大量細胞聚集,已經出現細胞聚集區(圖1A);實驗組兩側腭突不能在中線處接觸,間充質組織無法相互貫通,無明顯細胞聚集區(圖1B)。對照組間充質細胞分布出現區域差異,腭突生長迅速的部位即腭突突部細胞密集,根部則細胞稀疏;實驗組細胞分布呈現與對照組相似的規律,但實驗組腭突突部的細胞密集程度較對照組差。
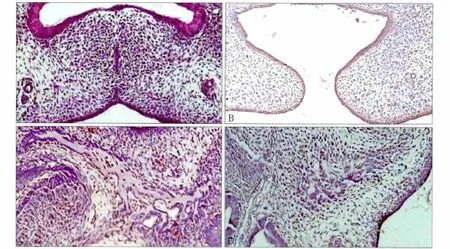
圖1 小鼠腭部發育的病理學觀察 HE × 100Fig 1 The morphology observation of mice embryonic palates HE × 100
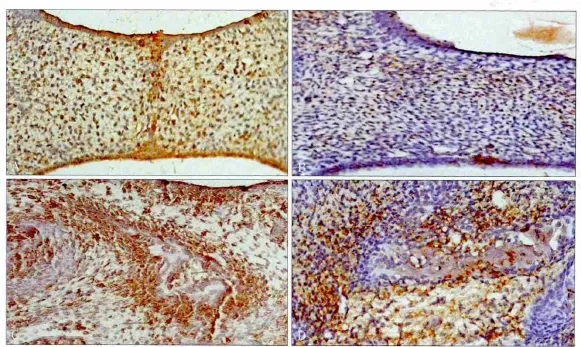
圖2 小鼠腭部BMPR2的免疫組織化學染色 SP × 100Fig 2 Immunohistochemical stain of BMPR2 of mouse embryonic palates SP × 100
GD17時,對照組雙側腭突完全融合,間充質細胞凝集區進一步擴大,腭胚突間充質細胞開始向成骨細胞分化,呈圓形,核仁大,出現多核細胞,細胞質嗜蘇木精染色,在中心區出現骨組織,骨化中心開始形成(圖1C);實驗組腭突沒有在中線處融合,形成體積較小的畸形腭突,兩側腭突間充質內可見成骨中心區,但其范圍明顯小于對照組(圖1D)。
免疫組織化學染色可見,GD15時,BMPR2在對照組胚胎腭部有高水平表達,在腭突上皮細胞和間充質細胞中都有陽性反應顆粒,陽性指數為75%~100%;實驗組BMPR2陽性指數低于25%,表達強度明顯低于對照組,主要分布在腭突間充質細胞的細胞質中(圖2A、B)。GD17時,對照組BMPR2染色強度較GD15時期變化不大,陽性指數達到50%~75%,陽性顆粒分布區域較廣,在細胞凝集區成骨中心周圍更為明顯;而實驗組的陽性指數約為25%,陽性染色主要集中在細胞凝集區,非凝集區的細胞陽性顆粒少(圖2C、D)。
RT-PCR檢測胚胎腭部Bmpr2 mRNA的表達情況見圖3。在兩個時間點對照組和實驗組均可檢測到Bmpr2 mRNA的表達,也可見到β-actin內參照的表達(圖3上)。分析Bmpr2 mRNA的相對表達量,對照組GD15時的表達水平明顯高于實驗組(P<0.05),GD17時的表達水平與GD15時差異不大,但仍明顯高于實驗組(P<0.05)(圖3下),提示atRA可能下調了腭突Bmpr2 mRNA的表達水平。
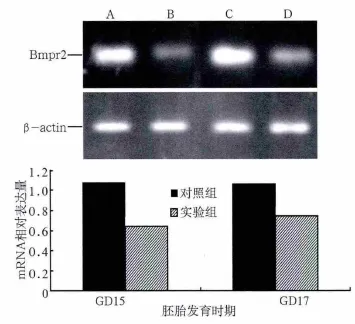
圖3 小鼠腭部Bmpr2 mRNA表達的RT-PCR測定Fig 3 Bmpr2 mRNA expression of mouse embryonic palate by RTPCR
3 討論
胚胎腭的發生發育期是致畸藥物作用的敏感時期[17]。小鼠胚胎7~12 d為腭器官發生期,胚胎12~15 d為腭發育形成期[18]。本研究在孕鼠GD10時使用atRA灌胃,實驗組腭突發育明顯滯后,雙側腭突未能及時接觸融合。有研究[19-20]認為,維甲酸的作用使腭突間充質細胞增殖受到抑制,分布異常,雙側腭突無法正常接觸融合導致腭裂畸形。本課題組在前期實驗[21-23]中發現,在小鼠GD10時采用100 mg·kg-1的atRA灌胃,誘導腭裂發生的比例為67%~100%,進一步研究發現,atRA誘導成功的絕大部分標本的腭突呈水平生長,但兩側腭突不能在中線處接觸并及時融合,間充質組織無法相互貫通。本實驗觀察到atRA誘導后胎鼠腭突間充質組織的細胞凝集區形成相對滯后且變小,提示實驗組小鼠胚胎腭部發育后期的成骨情況可能不如對照組,有可能導致腭部骨骼畸形的發生。有研究[24-25]發現,atRA能使細胞凝集區的細胞維持在低分化的間充質細胞狀態,阻止其向軟骨細胞分化,這種抑制軟骨分化的作用可能和atRA下調了轉化生長因子-β/Smad信號通路有關。Lu等[26]通過原位雜交實驗發現,在正常腭胚發育階段,BMP2、4、5在上皮和間充質都有一定量的表達,其后隨著腭部發育逐漸消退,但在細胞凝集區內依然有強表達;而atRA誘導的腭裂鼠模型中,Bmp2、4、5 mRNA表達水平較正常低,且細胞凝集區的出現時間延遲,腭突的形態萎縮變小,不能接觸融合而導致腭裂。Bandyopadhyay等[27]通過免疫組織化學染色進行半定量分析,結果發現,在腭突發育期間BMP2、4、7的表達量持續上升,在凝聚區尤為明顯,較散在間質區高出2~4倍,但維甲酸誘導的腭裂鼠中,BMP蛋白的表達量明顯減少。
BMP信號主要調控胚胎發育和組織穩態等重要進程,在骨骼發育中起關鍵作用[27]。有研究[26,28]指出,atRA使腭突間充質細胞區的Bmp2、4、5、7 mRNA明顯下降,提示妊娠期孕鼠維甲酸灌胃可能影響腭突區BMP信號的表達。本課題組在前期研究[21]中也發現,BMPR1b的表達與腭突間充質組織的生長發育呈正相關,在GD10灌胃維甲酸后,GD13—GD17胎鼠腭間充質中Bmpr1b mRNA以及蛋白的表達水平均下調。BMPR2是BMP的細胞膜受體,是BMPR1的激活劑,二者結合在一起形成受體復合物,激活下游的信號傳導途徑,如Smad蛋白等,從而調控基因轉錄,維持正常細胞功能活動[29]。小鼠神經嵴缺乏BMP信號將產生帶有多種顱頜面部骨骼缺陷的表型,在神經嵴細胞中敲除BMPR1a會誘使小鼠發生腭裂、下頜萎縮等顱面部畸形[30]。BMPR1a突變還將導致小鼠胚胎流產,同時流產胚胎無法形成中胚層組織[31]。進一步研究[32]指出,BMPR1b對骨骼形成也非常重要,在顱面部的骨骼發育區和成熟區都有高度表達,轉基因鼠實驗證明缺乏BMPR1b的小鼠將發生顱骨骨化延遲。近年來,在BMPR2作用機制及其與遺傳疾病、骨的形成與再生、哺乳動物的發育等關系的研究中取得了一些重要的進展。在哺乳動物胚胎發育早期,BMPR2調節多個信號通路的功能穩定,維持妊娠過程正常[33]。Bmpr2基因突變后,BMPR2蛋白表達下降,導致BMP信號通路功能受損[34]。盡管已經取得了一些進展,但目前對腭部BMPR2的相關研究仍較少,尚不明確它在維甲酸誘導腭突中的表達情況。本課題組在前期實驗[21]中觀察到,胎鼠腭間充質細胞在GD15—GD17出現較為明顯的細胞凝集區并逐步分化為成骨細胞,在中心區形成骨組織,在此基礎上,本實驗選擇GD15和GD17作為時間點,通過免疫組織化學和RT-PCR方法檢測BMPR2的表達是否受到atRA的影響,為下一步研究維甲酸對腭部成骨分化的影響提供基礎。
本實驗中,免疫組織化學結果顯示,正常腭部GD15的BMPR2處于高水平表達狀態,但在atRA誘導的腭突中,BMPR2的表達量下調。這種情況在GD17小鼠腭部進入分化期時依然持續。RT-PCR結果也證實Bmpr2 mRNA在實驗組的表達量明顯下調,提示atRA處理將下調腭突Bmpr2的表達量。Beppu等[35]通過基因打靶的方式培育了Bmpr2突變小鼠,發現在小鼠的早期發育中,Bmpr2對傳遞BMP信號至關重要,是外胚層分化和中胚層誘導所必需的關鍵因子。通過牙周組織再生研究[36]發現,白細胞介素-1β和生物機械力會通過下調BMPR2來抵消釉基質蛋白衍生物(enamel matrix derivative,EMD)誘導BMP2合成的作用。BMPR2無論在膜內成骨還是軟骨內成骨中都呈現出活躍表達,BMP及其受體系統能誘導軟骨細胞及成骨細胞的分化,破骨細胞的再吸收,對骨的形成(包括異位成骨)與再生起重要作用。在BMP2促進小鼠異位成骨過程中,未分化間充質細胞的BMPR2表達水平在早期即顯著上調[37]。在早期研究[26]中已發現維甲酸能導致腭突間充質細胞凝集區的發生延遲并減弱未分化細胞向成骨方向分化,因此筆者推測atRA通過下調BMPR2表達水平影響BMP及其受體信號通路,從而使細胞凝集區發生延遲,腭突發育不良,并影響骨化中心的形成,甚至會在腭部發育后期產生明顯的骨骼畸形。
BMP信號通路參與了體內眾多重要的生命活動過程,尤其是間充質細胞的成骨分化和骨骼形成,BMPR2作為該通路中的關鍵受體蛋白,其結構與含量的變化將會不同程度地影響該通路的正常運行。本實驗通過建立atRA誘導的小鼠腭裂模型來探索在腭部的BMP信號通路相關蛋白表達情況,發現在atRA誘導下,腭突發育滯后,無法融合,細胞凝集區出現延遲且變小,同時Bmpr2 mRNA表達水平下調,BMPR2陽性率降低,提示維甲酸可能通過胚胎期腭突的BMP及其受體信號通路來影響腭部發育,尤其是腭部間充質細胞的成骨分化進程,對于闡明其正常的生理功能和異常的病理機制具有重要意義。目前維甲酸影響胚胎頜面部BMP信號造成骨骼異常的研究尚處于起步階段,其BMP信號和成骨分化進程的變化模式還有待深入研究。
[1]Dai L, Zhu J, Mao M, et al. Time trends in oral clefts in Chinese newborns: data from the Chinese National Birth Defects Monitoring Network[J]. Birth Defects Res Part A Clin Mol Teratol, 2010, 88(1):41-47.
[2]代禮, 繆蕾, 周光萱, 等. 1996~2000年中國圍產兒腭裂畸形發生狀況分析[J]. 華西口腔醫學雜志, 2004, 22(1):35-37.
[3]Leslie EJ, Marazita ML. Genetics of cleft lip and cleft palate[J]. Am J Med Genet C Semin Med Genet, 2013, 163C(4):246-258.
[4]Abbott BD. The etiology of cleft palate: a 50-year search for mechanistic and molecular understanding[J]. Birth Defects Res B Dev Reprod Toxicol, 2010, 89(4):266-274.
[5]Ackermans MM, Zhou H, Carels CE, et al. Vitamin A and clefting: putative biological mechanisms[J]. Nutr Rev, 2011,69(10):613-624.
[6]李精韜, 石冰. 維生素A族與唇腭裂發生關系的研究進展[J]. 國際口腔醫學雜志, 2012, 39(3):346-348.
[7]Ross SA, McCaffery PJ, Drager UC, et al. Retinoids in embryonal development[J]. Physiol Rev, 2000, 80(3):1021-1054.
[8]汪淼, 黃洪章, 侯勁松. 小鼠體外胚胎腭器官培養模型的建立及全反式維甲酸致畸機制的研究[J/CD]. 中華口腔醫學研究雜志: 電子版, 2010, 4(3):215-223.
[9]Gutierrez-Mazariegos J, Schubert M, Laudet V. Evolution of retinoic acid receptors and retinoic acid signaling[J]. Subcell Biochem, 2014, 70:55-73.
[10]Diah E, Lo LJ, Huang CS, et al. Maxillary growth of adult patients with unoperated cleft: answers to the debates[J]. J Plast Reconstr Aesthet Surg, 2007, 60(4):407-413.
[11]Iwasaki H, Kudo M, Yamamoto Y. Does congenital cleft palate intrinsically in fl uence craniofacial morphology? Craniofacial features in unoperated submucous cleft palate children in prepuberty[J]. J Oral Maxillofac Surg, 2009,67(3):477-484.
[12]Young DL, Schneider RA, Hu D, et al. Genetic and terato-genic approaches to craniofacial development[J]. Crit Rev Oral Biol Med, 2000, 11(3):304-317.
[13]汪淼, 黃洪章. 側腭突生長發育和腭裂的分子調節機制[J/CD]. 中華口腔醫學研究雜志: 電子版, 2009, 3(6):667-674.
[14]黃洪章, 呂寶輝, 陳亦陽, 等. 維甲酸誘導小鼠腭裂發病機制的實驗研究[J]. 中華口腔醫學雜志, 2003, 38(3):185-187.
[15]沈璐, 叢蔚, 王如, 等. 維甲酸誘導腭裂相關wnt和成纖維細胞生長因子配體表達的動態變化[J]. 華西口腔醫學雜志, 2011, 29(1):62-65.
[16]Parada C, Chai Y. Roles of BMP signaling pathway in lip and palate development[J]. Front Oral Biol, 2012, 16:60-70.
[17]Levi B, Brugman S, Wong VW, et al. Palatogenesis: engineering, pathways and pathologies[J]. Organogenesis, 2011,7(4):242-254.
[18]Abbott BD, Harris MW, Birnbaum LS. Etiology of retinoic acid-induced cleft palate varies with the embryonic stage[J].Teratology, 1989, 40(6):533-553.
[19]黃洪章, 陳亦陽. 細胞凋亡與腭裂發生關系的實驗研究[J]. 中山醫科大學學報, 2000, 21(4):245-247.
[20]王如, 劉彬, 王博, 等. 腭發育不同時期維甲酸對腭突細胞增殖和凋亡的影響[J]. 華西口腔醫學雜志, 2008, 26(5):546-549.
[21]陳沐, 侯勁松, 王成, 等. 全反式維甲酸對C57小鼠胚胎腭突Bmpr-1b表達的影響[J/CD]. 中華口腔醫學研究雜志:電子版, 2012, 6(6):489-496.
[22]Chen M, Huang HZ, Zeng DL, et al. Cephalometric analysis of craniofacial malformations in newborn mice with cleft palate induced by retinoic acid[J]. Cleft Palate Craniofac J,2011, 48(2):197-204.
[23]呂寶輝, 黃洪章. RA誘導小鼠腭裂動物模型的實驗研究[J]. 口腔醫學研究, 2002, 18(4):248-250.
[24]Yu Z, Xing Y. All-trans retinoic acid inhibited chondrogenesis of mouse embryonic palate mesenchymal cells by downregulation of TGF-beta/Smad signaling[J]. Biochem Biophys Res Commun, 2006, 340(3):929-934.
[25]汪淼, 侯勁松, 黃洪章. RNA干擾原代培養胚胎腭間充質細胞Smad2/3基因對atRA阻滯細胞周期作用的影響[J].中國口腔頜面外科雜志, 2010, 8(4):352-358.
[26]Lu H, Jin Y, Tipoe GL. Alteration in the expression of bone morphogenetic protein-2,3,4,5 mRNA during pathogenesis of cleft palate in BALB/c mice[J]. Arch Oral Biol, 2000, 45(2):133-140.
[27]Bandyopadhyay A, Yadav PS, Prashar P. BMP signaling in development and diseases: a pharmacological perspective[J]. Biochem Pharmacol, 2013, 85(7):857-864.
[28]Guo L, Zhao YY, Zhang SL, et al. Retinoic acid downregulates bone morphogenetic protein 7 expression in rat with cleft palate[J]. Chin Med Sci J, 2008, 23(1):28-31.
[29]Chen G, Deng C, Li YP. TGF-β and BMP signaling in osteoblast differentiation and bone formation[J]. Int J Biol Sci,2012, 8(2):272-288.
[30]Dudas M, Sridurongrit S, Nagy A, et al. Craniofacial defects in mice lacking BMP typeⅠreceptor Alk2 in neural crest cells[J]. Mech Dev, 2004, 121(2):173-182.
[31]Mishina Y, Suzuki A, Ueno N, et al. Bmpr encodes a typeⅠbone morphogenetic protein receptor that is essential for gastrulation during mouse embryogenesis[J]. Genes Dev,1995, 9(24):3027-3037.
[32]Zhao M, Harris SE, Horn D, et al. Bone morphogenetic protein receptor signaling is necessary for normal murine postnatal bone formation[J]. J Cell Biol, 2002, 157(6):1049-1060.
[33]Nagashima T, Li Q, Clementi C, et al. BMPR2 is required for postimplantation uterine function and pregnancy maintenance[J]. J Clin Invest, 2013, 123(6):2539-2550.
[34]Majka S, Hagen M, Blackwell T, et al. Physiologic and molecular consequences of endothelial Bmpr2 mutation[J]. Respir Res, 2011, 12:84.
[35]Beppu H, Kawabata M, Hamamoto T, et al. BMP typeⅡreceptor is required for gastrulation and early development of mouse embryos[J]. Dev Biol, 2000, 221(1):249-258.
[36]Nokhbehsaim M, Deschner B, Winter J, et al. Interactions of regenerative, inflammatory and biomechanical signals on bone morphogenetic protein-2 in periodontal ligament cells[J]. J Periodont Res, 2011, 46(3):374-381.
[37]Nakamura Y, Wakitani S, Nakayama J, et al. Temporal and spatial expression profiles of BMP receptors and noggin during BMP-2-induced ectopic bone formation[J]. J Bone Miner Res, 2003, 18(10):1854-1862.
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
