野栽滲入系水稻籽粒儲藏蛋白質(zhì)含量的QTL遺傳解析
2015-07-31 17:20:35趙琳琳李楠呂志偉等
江蘇農(nóng)業(yè)科學(xué) 2015年3期
趙琳琳 李楠 呂志偉等

摘要:以云南元江普通野生稻為供體親本、以優(yōu)良秈稻品種特青為受體親本構(gòu)建了高代回交群體(BC3)。采用全自動凱氏定氮儀測定糙米中的總氮含量,用總氮含量乘以5.95估算儲藏的蛋白質(zhì)含量,蛋白質(zhì)含量采用Map Manager QTXb17軟件進(jìn)行數(shù)量性狀基因座(quantitative trait locus,QTL)定位分析,利用QTL檢測分析2次測定的儲存蛋白,結(jié)果共發(fā)現(xiàn)14個對蛋白質(zhì)含量有較高相關(guān)性的QTL。在第1號染色體的RM272、RM243、RM23、RM5、RM212、RM306位點和第2號染色體的RM250位點附近發(fā)現(xiàn)了7個能提高蛋白質(zhì)含量的QTL,它們均來源于野生稻;另外7個QTL位于第6、7、8、10號染色體上,分別是第6號染色體的RM345、第7號染色體的RM295、RM82、RM481、RM172,第8號染色體的RM25,以及第10號染色體的RM258附近的QTL,它們來源于栽培稻親本特青的等位基因作用表現(xiàn)為提高蛋白質(zhì)含量。在檢測到的QTL中,有6個QTL在2次分析中均被檢測到,經(jīng)分析是較穩(wěn)定的低蛋白QTL,它們分別為RM243、RM23、RM5、RM295、RM82、RM258。
關(guān)鍵詞:野生稻基因滲入系;籽粒儲藏蛋白質(zhì);凱氏定氮法;QTL分析
中圖分類號: Q946.1 文獻(xiàn)標(biāo)志碼: A
文章編號:1002-1302(2015)03-0050-03
水稻是一種重要的糧食作物,世界上約50%的人口以稻米為主食,稻米蛋白質(zhì)被稱作優(yōu)質(zhì)植物蛋白[1],通過與其他谷物進(jìn)行比較可知,水稻中必需氨基酸的含量大體上要高于聯(lián)合國糧食及農(nóng)業(yè)組織(FAO)/世界衛(wèi)生組織(WHO)建議的標(biāo)準(zhǔn)水平[2],其第一限制性氨基酸賴氨酸含量高于其他谷類,是人類蛋白質(zhì)的主要來源之一。在發(fā)展中國家,約60%的蛋白來源于谷物,在未來的幾十年里,谷物仍是世界上2/3人口的主要蛋白來源[3]。因此,提高稻米蛋白質(zhì)含量已經(jīng)成為當(dāng)今水稻品質(zhì)育種的一個重要內(nèi)容;同時,隨著慢性腎功能不全患者不斷增加,培育低蛋白含量,特別是培育低可消化吸收蛋白(谷蛋白)含量水稻品種已成為功能水稻的新育種目標(biāo)。
籽粒儲存蛋白受多基因控制,屬于數(shù)量遺傳性狀,開展控制籽粒儲存蛋白含量的數(shù)量性狀基因座(quantitative trait locus,QTL)定位與克隆,不僅對水稻種質(zhì)資源創(chuàng)新和高蛋白米、低蛋白米的新品種選育有重要的理論意義,而且對當(dāng)前調(diào)整農(nóng)業(yè)結(jié)構(gòu)、保持農(nóng)業(yè)可持續(xù)發(fā)展同樣具有重要的現(xiàn)實意義。
迄今為止,大量的研究主要是對水稻農(nóng)藝性狀的遺傳分析和基因定位,并且對稻米品質(zhì)性狀的遺傳分析和基因定位研究主要集中于加工品質(zhì)、外觀品質(zhì)和蒸煮及食味品質(zhì)等性狀上[4-9],而關(guān)于水稻糙米儲存蛋白定位的報道并不多見,僅限于個別報道。于永紅等利用協(xié)青早B/密陽46的重組自交系群體對水稻糙米蛋白質(zhì)含量的QTL分析表明,5個控制蛋白質(zhì)含量的QTLs(qPc-3、qPc-4、qPc-5、qPc-6、qPc-10)均被檢測到,單個QTL對群體表型變異的貢獻(xiàn)率為4.17%~19.14%;其中第6號染色體的Wx基因區(qū)域?qū)Φ鞍踪|(zhì)含量具有主效作用[10]。Li等利用亞洲栽培稻和非洲栽培稻的回交高代群體,在第8號染色體上定位到了1個QTL[11]。Yoshida等利用DH群體(由花藥單倍體誘導(dǎo)得到)定位到了分別位于第 8、9、11、12號染色體上的4個控制稻米蛋白含量的QTLs,其中第11號染色體RM206位點附近的QTL效應(yīng)較大[12]。Aluko等利用非洲野生稻與秈稻BC3F1回交構(gòu)建的DH群體,分別在第1、2、6、11號染色體上檢測到4個與蛋白質(zhì)含量相關(guān)的QTLs,其中位于第1號染色體RM226至RM297位點之間的QTL貢獻(xiàn)率較大[13]。張濤等檢測到6個QTLs (qPc-3、qPc-6、qPc-7、qPc-8-1、qPc-8-2、qPc-11)對糙米蛋白質(zhì)含量進(jìn)行控制,它們分別位于第3、6、7、8、11號染色體上,單個QTL對群體表型變異的貢獻(xiàn)率為379%~19.41%[14]。在這些QTL的區(qū)間中,第8染色體的基因區(qū)域?qū)Σ诿椎鞍踪|(zhì)含量具有主效作用。李晨等利用BC4群體共定位到了6個糙米蛋白QTLs,分別位于第1、2、8、9號染色體上,其中位于第1號染色體RM472位點附近的QTL具有主效作用[15]。
野生稻中蘊含著豐富的遺傳信息,是栽培稻的祖先,因此發(fā)掘野生稻中的有益基因不僅有利于解決當(dāng)前育種資源狹窄的現(xiàn)狀,而且是突破育種瓶頸的有效方法之一。但是,目前關(guān)于從野生稻中定位QTL的報道并不多,各個方面都待完善,因此今后需要進(jìn)一步強(qiáng)化這方面的工作。本研究通過采用凱氏定氮法對野生稻染色體片段代換系(簡稱水稻野栽滲入系)的籽粒儲存蛋白質(zhì)含量進(jìn)行分析,并對控制籽粒儲存蛋白質(zhì)含量的QTL進(jìn)行了檢測。
1 材料與方法
1.1 群體構(gòu)建
本研究所構(gòu)建的野栽滲入系由中國農(nóng)業(yè)大學(xué)孫傳清博士提供,是以元江普野為供體親本、秈稻品種特青為受體親本培育而成的。在BC3F2群體的基礎(chǔ)上,連續(xù)自交3次得到BC3F3、BC3F4群體。根據(jù)BC3F2株系表型鑒定和基因分析結(jié)果,滲入系構(gòu)建的基本材料選取了263個BC3F3、BC3F4單株,最終滲入系由106個BC3F3、BC3F4候選單株構(gòu)成。
1.2 SSR引物擴(kuò)增及數(shù)據(jù)處理
根據(jù)Temnykh等發(fā)表的水稻SSR序列合成引物,用在親本間有多態(tài)性的112個SSR標(biāo)記,調(diào)查上述野栽滲入系群體106個系的基因型[16]。在基因型分析中,“B”表示與親本特青帶型相同,“A”表示與野生稻親本帶型相同,“H”則表示具雜合帶型,“-”表示帶型模糊或由于某種原因缺失。采用MAPMAKER/EXP3.0軟件并參考Temnykh等發(fā)表的水稻SSR連鎖圖譜為框架,構(gòu)建連鎖圖譜,可見112個SSR標(biāo)記分布于12條染色體上,各染色體上最多的有13個標(biāo)記,最少的有7個標(biāo)記,平均每條染色體有9.3個標(biāo)記,標(biāo)記平均間距17.8 cM,基本能滿足QTL定位需要[16]。
1.3 糙米儲藏蛋白質(zhì)含量的測定及QTL分析
選用2006年于北京采收的種子,挑選表型一致(如大小、顏色)的水稻籽粒,剝?nèi)シf殼后于干燥箱(105 ℃)中干燥5 h,然后以8粒種子為1組稱質(zhì)量,每個家系重復(fù)5次。采用意大利產(chǎn)全自動凱氏定氮儀(UDK 142)測定糙米中的總氮含量,用總氮含量乘以5.95估算儲藏的蛋白質(zhì)含量[17-18]。為了確保測定結(jié)果的準(zhǔn)確性,同時選用2007年采收于北京的種子進(jìn)行同樣的測定,重復(fù)3次。
采用Map Manager QTXb17軟件,取P<0.05作為判斷QTL存在的閾值,并用單位點分析法對籽粒儲藏蛋白質(zhì)含量進(jìn)行QTL定位與遺傳分析[19]。
2 結(jié)果與分析
2.1 籽粒儲藏蛋白質(zhì)含量的分布
圖1是不同家系的蛋白質(zhì)含量分布情況。可見滲入系群體的蛋白質(zhì)含量趨于正態(tài)分布,符合數(shù)量性狀的特征,適于QTL定位分析。
2.2 QTL解析
將滲入系群體的籽粒蛋白質(zhì)含量數(shù)據(jù)采用Map Manager QTXb17軟件進(jìn)行QTL定位分析,共檢測到11個與蛋白質(zhì)含量相關(guān)的QTL(表1)。其中位于第1號染色體RM272、RM243、RM23、RM5、RM306位點附近以及位于第2號染色體RM250位點附近的計6個QTL,貢獻(xiàn)率分別為5%、8%、6%、7%、5%、6%,加性效應(yīng)分別為-0.61%、-0.91%、-0.56%、-0.57%、-0.62%、-0.69%,來源于野生稻的等位基因能夠提高糙米籽粒儲藏蛋白質(zhì)含量。另外5個QTL位于第6、7、10號染色體上,分別是第6號染色體RM345位點,第7染色體RM295、RM481、RM82位點,以及第10染色體RM258位點附近的QTL,貢獻(xiàn)率分別為5%、9%、6%、7%、5%,加性效應(yīng)值分別為084%、0.77%、0.58%、0.59%、047%,來源于特青的等位基因表現(xiàn)為提高糙米籽粒儲藏蛋白質(zhì)含量。
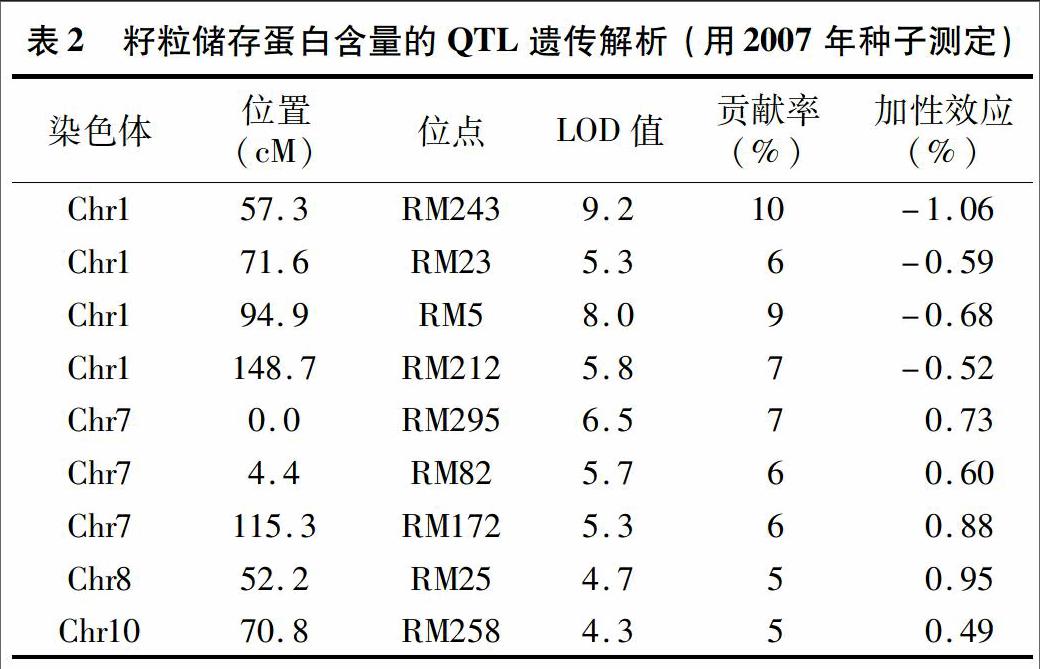
2007年批次種子的重復(fù)試驗結(jié)果見表2,共檢測到9個與蛋白質(zhì)含量相關(guān)的QTL。分別位于第1、7、8、10號染色體上,貢獻(xiàn)率為5%~10%。與表1相比可知,在第2、6號染色體上沒有檢測到QTL,而在第8號染色體上檢測到的來源于特青的、提高蛋白質(zhì)含量的QTL在第1次的結(jié)果中并沒有檢測到。
2次結(jié)果均檢測到的QTL位點見表3,可見有位于第1號染色體的RM243、RM23、RM5位點附近的3個QTL,表現(xiàn)為野生稻等位基因提高糙米籽粒儲藏蛋白質(zhì)含量;還有位于第7號染色體RM295、RM82位點和第10號染色體RM258位點附近的3個QTL,表現(xiàn)為特青等位基因提高糙米籽粒儲藏蛋白質(zhì)含量。
3 結(jié)論與討論
本研究以元江普野為供體親本、秈稻品種特青為受體親本構(gòu)建的野栽滲入系群體的種子為材料,在全自動凱氏定氮儀中消化并測定剝掉穎殼后糙米的含氮量,用含氮量乘以595作為糙米蛋白質(zhì)含量的計算指標(biāo)。用Map Manager QTXb17軟件,取P<0.05作為判斷QTL存在的閾值,并用單位點分析法,對籽粒儲藏蛋白質(zhì)含量進(jìn)行QTL定位與遺傳分析,2次共定位到14個QTL位點。其中來自野生稻親本的高蛋白QTL有7個,分別分布于第1、2號染色體上,第1號染色體上有6個,第2號染色體上有1個。來源于栽培稻親本特青的高蛋白QTL也有7個,分別分布于第6、7、8、10號染色體上,第7號染色體上有4個,其余染色體上各有1個。
通過與以前的研究結(jié)果相比較,我們發(fā)現(xiàn)有5 個QTL位點與前人所報道的QTL位點在同一區(qū)域或者在相臨區(qū)域。位于第1號染色體上RM23位點附近的QTL與李晨等報道的qCP1-1[15]在同一區(qū)域。位于第1號染色體上RM212位點附近的QTL,以及位于第2號染色體上RM250位點附近的QTL分別與Aluko等報道的Pro1、Pro2[13]在同一區(qū)域。位于第7號染色體RM172位點附近的QTL與吳長明等報道的Pr3[20]處于相臨區(qū)域。位于第8號染色體RM25位點附件的QTL與張濤等報道的qPc-8-2[14]在相近區(qū)域,經(jīng)分析是同一位點。
此外,經(jīng)過分析有9個QTL位點與以前報道的位置不一樣,是檢測到的新QTL;這9個QTL位點分別位于第1號染色體RM272、RM243、RM5、RM306位點附近,第6號染色體RM345位點附近,第7號染色體RM295、RM481、RM82位點附近,以及位于第10號染色體RM258位點附近。
在本研究中,沒有檢測到主效QTL,檢測到的QTL貢獻(xiàn)率最大的為10%,這一結(jié)論與Li等的研究結(jié)果[11,21-22]類似,而與于永紅等的研究結(jié)果[10,12-15,23]有所差異,認(rèn)為這種差異可能是由于所用的材料不同造成的。
本研究共從野生稻中檢測到7個控制籽粒儲存蛋白質(zhì)的QTL,其中在第1號染色體上有6個,在第2號染色體上有1個,這為今后進(jìn)一步研究分析和利用這些QTL打下了基礎(chǔ)。
參考文獻(xiàn):
[1]周麗慧,劉巧泉,張昌泉,等. 水稻種子蛋白質(zhì)含量及組分在品種間的變異與分布[J]. 作物學(xué)報,2009,35(5):884-891.
[2]王治平. 大米品質(zhì)與主食米飯食用品質(zhì)相關(guān)性的研究[D]. 長沙:湖南農(nóng)業(yè)大學(xué),2000:2-4.
[3]姚惠源. 稻米深加工[M]. 北京:化學(xué)工業(yè)出版社,2004.
[4]Tan Y F,Li J X,Yu S B,et al. The three important traits for cooking and eating quality of rice grains are controlled by a single locus in an elite rice hybrid,Shanyou 63[J]. Theoretical and Applied Genetics,1999,99(3/4):642-648.
