福建省軍團菌毒力島magA基因攜帶情況分析
2015-06-05 08:39:58李曲文鄭恩惠陳愛平楊勁松原靈王靈嵐
實驗與檢驗醫學 2015年4期
李曲文,鄭恩惠,陳愛平,楊勁松,原靈,王靈嵐
(1、福建省疾病預防控制中心,福建 福州350001;2、福建省人獸共患病研究重點實驗室,福建 福州350001)
軍團菌是一類革蘭陰性菌,軍團菌在許多環境中都很常見,目前已知的軍團菌屬有58個種70多個血清型,其中能引起人類疾病的大概有20種,致病性最強的為嗜肺軍團菌[1-3]。軍團菌的整個致病過程是其基因組上各個毒力基因及其表達產物共同作用的結果,不同軍團菌的致病力強弱與其所帶毒力因子不同有關[4,5],有些毒力因子成為了近年來的研究重點和熱點。本文利用PCR方法擴增2012-2013年30株軍團菌菌株分析毒力島magA基因攜帶情況,以初步了解我省分離株的毒力島magA基因攜帶率。
1 材料與方法
1.1 實驗用菌株 2012年-2013年福建省中央空調冷卻塔水系統軍團菌監測分離的菌株30株軍團菌菌株。其中 16株 LP1,2株 LP4,3株 LP5,5株LP7,4 株 N-LP。
1.2 試劑儀器 Taq DNA聚合酶、dNTP購自大連寶生物工程有限公司,瓊脂糖為BioAsia分裝品。Gel Doc XR+凝膠成像系統,Basic電泳儀,PCR擴增儀為G-Storm(英國GeneTech公司)。
1.3 基因引物 本研究所用的基因引物序列magA基因:F:5’-CTCTATCGCTAACGCACAAGG-3’ R:5’-CGTTGAAGTAGTTAGTGAAAG-3’,參考文獻[5],其相應的擴增片段大小為469bp,寡核苷酸由上海生工生物工程技術服務有限公司合成。
1.4 模板的制備 分純的細菌菌落,直接刮取適量于300μl的純水制備成菌懸液,100℃煮沸裂解10 min,10000r/min離心5min,取上清做為PCR檢測的DNA模板。
1.5 PCR反應條件 PCR反應均采用20μl的反應體系,各組成成分的終濃度分別為:rTaq DNA聚合酶 0.5 U,dNTP 200μmol/L, 上游引物0.4μmol/L,下游引物 0.4μmol/L,DNA 模板 3μl。
2 結果
2.1 PCR結果 30株軍團菌菌株magA基因PCR檢測電泳結果,見圖1。
2.2福建省30株軍團菌magA基因攜帶率情況見表1。
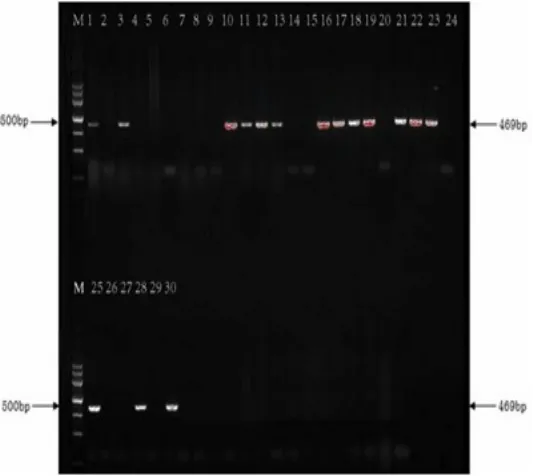
圖1 PCR電泳結果圖
3 討論
軍團菌是一種兼性細胞內致病菌,軍團菌屬的致病性主要通過細菌的胞內生命循環過程、毒力島基因座及毒力基因來完成。嗜肺軍團菌的致病性與其毒力攜帶情況密切相關,毒力基因可以增加了細菌的生存的適應性,是細菌演變及其致病性的關鍵元素之一[6-8]。毒力島作為細菌染色體上一段具有典型結構特征的基因簇,毒力島的形成是通過細菌基因組水平的轉移而改變其特性,可以增加細菌的適應性,是細菌演變及致病性的關鍵,與軍團菌多種致病菌的毒力因子的產生和細菌的進化有著密切的聯系[9-11]。嗜肺軍團菌含有一個65kb的毒力島,該毒力島共含有約70個不同的基因。該基因座包含Ⅳ型分泌系統、易變遺傳因子和毒力因子等,是嗜肺軍團菌血清型1(LP1)所特有的[12]。
本研究選用毒力島基因中間的magA(MIF associatied gene)基因作為檢測毒力島目的基因,magA基因是毒力島70個基因中的第17個,它編碼的20~24kDa的magA蛋白,據推測是表達軍團菌在細胞內生長晚期的特異性蛋白,可以作為檢測細菌在體內細胞中生長狀態的有用標記物[13]。本實驗不但檢測了嗜肺軍團菌LP1血清型毒力島攜帶情況,同時還檢測了14株非LP1型。根據實驗結果顯示16株LP1毒力島陽性10株,2株LP4均陽性,5株LP7陽性3株,4株非嗜肺軍團菌陽性1株。30株軍團菌菌株毒力島攜帶率為53.33%(16/30),說明了我省環境分離菌株具有較強的致病力,對環境有較好的適應性,可在外環境中長期存活。同時這些數據還表明毒力島并不是嗜肺軍團菌LP1所特有的,其他血清型也可攜帶。各部門應按照衛生部公共場所集中空調通風系統衛生規范[14,15],加強中央空調冷卻塔水監測,以防軍團菌病的發生。
[1]朱慶義,宋亞軍,邵祝軍,等.軍團菌和軍團菌病[M].北京:科學出版社,2014:8-10.
[2]原靈,陳愛平,詹鑾峰,等.2008年福建省5設區市公共場所中央空調冷卻塔水軍團菌污染狀況調查 [J].中國人獸共患病學報,2010,26(11):1080-1082.
[3]李曲文,原靈,陳愛平,等.1株非嗜肺軍團菌的鑒定[J].實驗與檢驗醫學,2012,30(6):535-536.
[4]朱俊,盛躍穎,陳敏,等.軍團菌毒力因子致病作用的研究進展[J].上海預防醫學雜志,2009,21(11):579-580.
[5]Zhu QY,Hu ZH,Liang YM,et al.Genotyping identificationof pathogenicity island locus for Legionella pneumopgila[J].Chin J Med Technol,2007,3(1):5-12.
[6]朱慶義,胡朝暉,梁耀銘,等.嗜肺軍團菌毒力島基因聚合酶鏈反應分型鑒定[J].中華檢驗醫學雜志,2006,29(7):647-648.
[7]曾瑾,王玉炯,鄧光存.毒力島與細菌致病性[J].生物學雜志,2010,27(1):80-96.
[8]Samrakandi MM,Cirillo SL,Ridenour DA,et al.Genetic and phenotypic differences between Legionella pneumophila strains[J].J Clin Microbiol,2002,40(4):1352-1362.
[9]De Buck E,Anné J,Lammertyn E.The role of protein secretion systems in the virulence of the intracellular pathogen Legionella pneumophila[J].Microbiology,2007,153(Pt 12):3948-3953.
[10]Urwyler S,Nyfeler Y,Ragaz C,et al.Proteome analysis of Legionella vacuoles purified by magnetic immunoseparation reveals secretory and endosomal GTPases[J].Traffic,2009,10(1):76-87.
[11]Newton HJ,Ang DK,van Driel IR,et al.Molecular pathogenesis of infections caused by Legionella pneumophila[J].Clin Microbiol Rev,2010,23(2):274-298.
[12]Brassinga AK,Hiltz MF,Sisson GR,et al.A 65-kilobase pathogenicity island is unique to Philadelphia-1 strains of Legionella pneumophila[J].J Bacteriol,2003,185(15):4630-4637.
[13]Hiltz MF,Sisson GR,Brassinga AK,et al.Expression of magA in Legionella pneumophila Philadelphia-1 is developmentally regulated and a marker of formation of mature intracellular forms[J].J Bacteriol,2004,186(10):3038-3045.
[14]衛生部.公共場所集中空調通風系統衛生規范[S].2006.
[15]衛生部.中華人民共和國衛生行業標準.WS195-2001.軍團菌病診斷標準和處理原則[S].2001.
