自噬基因微管相關蛋白1輕鏈3、 Beclin 1在 饑餓大鼠心肌中的表達及意義
2015-05-02 06:56:18張建凱張小平褚迎欣
疑難病雜志 2015年7期
關鍵詞:模型
張建凱,張小平,褚迎欣
?
論著·基礎
自噬基因微管相關蛋白1輕鏈3、Beclin1在 饑餓大鼠心肌中的表達及意義
張建凱,張小平,褚迎欣
目的 觀察饑餓模型大鼠心肌細胞中微管相關蛋白1輕鏈3(LC3)及自噬基因Beclin1的表達變化,探討自噬機制在饑餓大鼠心肌中的作用。方法 將42只健康雄性SD大鼠隨機分為正常對照組(n=6)和饑餓模型組(n=36)。正常對照組正常飲食,饑餓模型組禁食但自由攝水,根據饑餓天數又分為1、2、3、5、7、9d等6個亞組(n=6)。各亞組在相應時間進行心臟標本提取并留存,采用免疫組化法和蛋白質印跡法檢測心肌自噬蛋白Beclin1和LC3水平變化。結果 與正常對照組相比,饑餓模型組各時間點SD大鼠心肌的自噬蛋白LC3和Beclin1陽性細胞數及蛋白表達都有不同程度的增加,差異有統計學意義(P<0.01或P<0.05),饑餓模型各亞組之間自噬蛋白LC3和Beclin1陽性細胞數及蛋白表達差異亦均有統計學意義(P<0.01或P<0.05),且在饑餓2d和7d出現2個峰值。結論 饑餓狀態可通過激活心肌自噬機制參與心肌的病理改變過程。
自噬基因;Beclin1;微管相關蛋白1輕鏈;心肌;饑餓;大鼠
自噬是指細胞在外界環境因素的影響下,對其內部受損的細胞器、錯誤折疊的蛋白質和侵入其內的病原體進行降解的生物學過程,其主要作用是清除和降解自身受損的細胞器以及多余的生物大分子,并利用降解產物提供能量和重建細胞結構,進而在維持細胞穩態、促進細胞生存方面起重要作用。在正常和輕微應激下,作為一種細胞保護機制,自噬能降解和回收細胞器組分,如長半衰期蛋白并選擇性移除受損線粒體。因為受損線粒體能釋放促凋亡因子如細胞色素C,故自噬可能阻斷凋亡[1]。誘導自噬發生的方法和手段有很多種,饑餓是有效的方法之一,本實驗采用全饑餓狀態SD大鼠模型,觀察大鼠心肌Beclin 1和微管相關蛋白輕鏈3(LC3)的表達變化,探討饑餓狀態下自噬機制對心肌病理改變的影響,報道如下。
1 材料與方法
1.1 實驗材料
1.1.1 實驗動物: 健康雄性Sprague-Dawley(SD)大鼠42只,SPF級,10周齡,體質量330~370 g,購自中國人民解放軍軍事醫學科學院[許可證號:SCXK-(軍)2009-003]。室溫控制在(23±2)℃,自然光照,適應性喂養1周適應環境后,單籠飼養1周以適應獨居環境。
1.1.2 主要試劑與儀器: 多聚甲醛(天津市永大化學試劑有限公司提供);Anti-LC3(MBL International Corporation)、Rabbit Anti-Beclin 1(北京博奧森生物技術有限公司提供);免疫組化試劑盒SP9001(北京中山金橋生物技術有限公司提供);SDS-PAGE試劑盒、HRP標記的山羊抗兔IgG、ECL試劑盒、BCA蛋白濃度測定試劑盒(碧云天生物技術公司提供); 金相研究級正立顯微鏡Olympus BX61;高速臺式離心機購自上海安亭科學儀器廠。
1.2 模型制備與取材 實驗大鼠按隨機數字表法分為正常對照組(n=6)和饑餓模型組(n=36),后者完全禁食但自由飲水,并根據饑餓時限分為饑餓1、2、3、5、7、9d6個亞組(n=6)。每只大鼠取心尖部肌肉組織用于免疫組化和蛋白質印跡法(Westernblotting)檢測自噬相關蛋白Beclin1及LC3的表達,從大鼠禁食第1天8∶00開始,按禁食1、2、3、5、7、9d的順序進行取材,對照組取材時間與饑餓模型組時間相對應。取材方法:將SD大鼠麻醉并開胸后,沿主動脈根部游離心臟,鉗夾主動脈根部,在止血鉗的遠心端剪斷主動脈及周圍結締組織,立即放入生理鹽水中放開止血鉗并攪動洗滌約5s,取出放在冰袋上切取一部分心尖組織樣本于-80℃冰箱內冰凍,以備做Westernblotting用,其余部分再用生理鹽水充分攪拌清洗2次,放在4%多聚甲醛磷酸緩沖溶液中留存,以備做免疫組化用。1.3 觀察指標與方法
1.3.1 一般情況觀察: 觀察2組大鼠在實驗期間活動、皮毛、體質量、排便、是否易激惹等變化。
1.3.2Beclin1和LC3表達的檢測: (1)免疫組化法陽性細胞計數。取上述多聚甲醛固定好的心尖組織常規包埋、切片、脫蠟、水化、抗原修復;PBS洗后加封閉液(正常山羊血清工作液),加一抗[RabbitAnti-Beclin1(1∶200);Anti-LC3(1∶500)]4℃過夜,加二抗(生物素標記山羊抗兔IgG),加三抗(辣根酶標記鏈霉卵白素工作液),DAB顯色,顯微鏡下操作控制顯色時間以免過度顯色,再經過蘇木精染色、分化、藍化、梯度酒精脫水、二甲苯、中性樹膠封片。所有切片,在高倍鏡下觀察細胞形態變化,染色以細胞漿內出現棕黃色顆粒為陽性細胞,隨機觀察6個高倍視野,每個視野計數100個細胞,根據其中陽性細胞的比率進行評估。(2)Westernblotting蛋白印跡法。取上述冷凍好的心尖組織,進行Westernblotting檢測,一抗選兔抗人Anti-LC3,稀釋度為1∶1 000;RabbitAnti-Beclin1,稀釋度為1∶500,二抗選辣根過氧化酶標記山羊抗兔二抗體,稀釋度為1∶2 000,內參選β-actin,稀釋度為1∶1 000。運用Image-ProPlus5.1 圖像分析軟件測定灰度值。
1.4 統計學方法 應用SPSS19.0軟件進行統計分析。計量資料以均數±標準差表示,多組間比較采用單因素方差分析,組間兩兩比較用LSD法檢驗。以P<0.05為差異有統計學意義。
2 結 果
2.1 一般情況觀察 對照組大鼠在實驗期間進食活動正常,皮毛整潔,體質量增長。饑餓模型組大鼠從饑餓2d起即出現易激惹現象,大便稍潤,黑色;饑餓5d時出現精神狀態差,皮毛蒼松并見稀少偏黃,活動減少,體溫下降;饑餓7d時消瘦單薄,少活動,但易激惹,反應仍較迅速,毛發稀黃光澤度差,尾部有掉屑,有的糞便成形,黑索狀但干癟;饑餓9d時出現靜臥少動且行動遲緩,啃食鐵籠,目光呆滯,毛發灰黃稀長,行動無力,顫抖,弓背,步履蹣跚,體溫低,尾部掉屑,觸之冰涼感,體質量下降明顯。
2.2 各組大鼠心肌LC3和Beclin1的免疫組化檢測結果 與正常對照組比較,饑餓模型組各亞組大鼠心肌組織LC3與Beclin1表達上調,差異有統計學意義(P<0.05或P<0.01)。饑餓模型組各亞組間比較,LC3與Beclin1表達差異均有統計學意義(P<0.05或P<0.01),且在饑餓模型1d亞組和2d亞組,LC3與Beclin1表達持續增高,于饑餓模型2d亞組達到峰值后表達逐漸下降,在饑餓模型7d、9d亞組,LC3與Beclin1表達再次升高。見表1,圖1~2見封3。
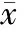
表1 各組大鼠心肌LC3和Beclin1陽性細胞數 (±s,n=6,/100個細胞)
注:與正常對照組比較,aP<0.05
2.3 各組大鼠心肌LC3和Beclin1的蛋白質印跡法檢測結果 與正常對照組比較,饑餓模型組各亞組大鼠心肌組織LC3與Beclin1表達上調,差異有統計學意義(P<0.05或P<0.01)。饑餓模型組各亞組間比較,LC3與Beclin1表達差異均有統計學意義(P<0.05或P<0.01),且在饑餓模型組1d亞組和2d亞組,LC3與Beclin1表達持續增高,于饑餓模型2d亞組達到峰值后表達逐漸下降,在饑餓模型7d、9d亞組,LC3與Beclin1表達再次升高。見表2,圖3。
3 討 論
自噬是細胞利用溶酶體降解自身受損的細胞器和大分子物質的過程。自噬一方面因具有底物水平的作用而更新長壽命蛋白[2];另一方面自噬可在饑餓等營養匱乏或疾病狀態下被激活,出現特征性的具有雙層膜結構的自噬體,自噬體通過微管系統運輸至溶酶體并融合形成自噬溶酶體,降解產物通過多種酶消化及降解釋放入胞漿而被細胞再利用[3~5],同時細胞可啟動自噬來清除受損的線粒體避免凋亡因子釋放進入胞漿,進而提高細胞對低氧的耐受力,保護細胞免于凋亡和各種毒素的侵襲。基礎水平的自噬可以維持心肌細胞的正常大小和心臟的正常結構[6],但在心臟疾病時,心肌細胞的自噬常常活化,并作為心臟應激的一種保護通路。有關研究表明,在急性心肌梗死發生后,細胞自噬對維持缺血缺氧等饑餓狀態下的氨基酸和ATP水平發揮重要作用,對維持心肌細胞穩態具有重要意義[7,8];饑餓小鼠心肌細胞中氨基酸和ATP水平在無自噬抑制劑時增高,但是給予自噬抑制劑后心肌細胞中都降低,顯示饑餓狀態時自噬在維持心功能方面起著重要作用[9]。然而,過度的自噬也會引起細胞的損
傷或死亡。Rokeach等[10]指出,過度的細胞應激引起的非選擇性自噬是導致自噬性細胞死亡的重要原因。國外有研究者發現在壓力負荷所致的慢性心功能不全小鼠中心肌細胞自噬明顯增加并伴有心肌細胞的重構,加重心功能不全[11]。因此,自噬對細胞來說是一柄雙刃劍,其利弊取決于疾病進展的不同階段、細胞周圍環境的變化和治療干預措施的不同。
Beclin1既是調控自噬的關鍵因子之一,也是自噬重要基因之一[12],作為自噬標記物的Beclin1已成為檢測自噬活性的重要方法,其表達與自噬活性密切相關。LC3是哺乳動物細胞中酵母自噬相關基因8(ATG8)的同源物,作為被發現的第一種自噬體標記蛋白分為LC3-Ⅰ和LC3-Ⅱ兩種形式。當發生自噬時,Ⅰ型LC3經泛素樣加工修飾過程,與自噬膜表面的磷脂酰乙醇胺結合形成Ⅱ型LC3并聚集于胞內自噬體膜上。因其含量與自噬泡數量呈正比,故LC3 被認為是自噬體的標志分子[13,14]。因此,通過實驗方法檢測細胞LC3 的含量,可以判斷細胞自噬或被誘導或被抑制。本實驗通過免疫組化及蛋白印記法檢測Beclin-1和LC3陽性細胞數及蛋白含量,結果均顯示:正常對照組大鼠心肌LC3和Beclin-1的陽性細胞數及蛋白表達水平較低,而饑餓模型組大鼠心肌LC3和Beclin-1的陽性細胞數及蛋白表達水平明顯升高,差異有統計學意義(P<0.05);在饑餓模型組的第2、7天出現LC3和Beclin1陽性細胞數及蛋白表達出現2個小高峰,同時饑餓模型各亞組之間心肌Beclin1和LC3的表達差異均有統計學意義(P<0.05)。以上研究提示:隨著饑餓時限的延長,發生心肌細胞線粒體通透性及結構損傷,誘導心肌細胞啟動自噬來清除損傷的線粒體,從而自噬可能起到心肌保護作用。但持續和過強的自噬可通過降解必需蛋白質和細胞器而導致細胞溶解死亡[15]。這與饑餓模型組大鼠心肌形態學改變相一致。
綜上所述,饑餓狀態下大鼠心肌組織LC3與Beclin1表達上調,短期饑餓2dLC3與Beclin1表達達到峰值,自噬增強,長期饑餓LC3與Beclin1表達再次上調,自噬體數量增多。這種結果的出現,可能是因為自噬在不同的發病時期或者在自噬自身不同的發展階段所產生的不同的作用方式,對細胞造成不同的作用效果。初期,即在饑餓1~2d時,被饑餓誘發出的自噬是保護性的,通過清除和降解受損破壞的細胞器、變性壞死的蛋白質,維持內環境的穩定并為機體提供營養成分和能量以維持機體的生存和基礎生理功能,形成第一個基因表達的高峰;而隨著饑餓時間的延續自噬作用也在隨之增強,即在饑餓7d左右,機體內受損細胞器和變性蛋白質已被清除或降解殆盡,但是增強的自噬進程并沒有停止,而是開始降解正常的發揮基礎功能的細胞器或蛋白質,這就是過度自噬的作用,顯然具有破壞性,于是,出現了第2個小高峰,并且,因為機體細胞嚴重受損,大鼠很快趨于死亡,這和大鼠一般情況觀察的結果也是相吻合的,饑餓2d時,因為自噬的保護機制,大鼠的身體機能變化不是很大,只是出現“易激惹”現象,而“饑餓5d時出現精神狀態差,皮毛蒼松并見稀少偏黃,活動減少,體溫下降”出現了明顯的機能下降,饑餓7d時,“精神狀態差,毛發稀黃光澤度差,尾部有掉屑”,此時,自噬的破會性作用開始,因為即便是破壞性自噬,初期也會為機體提供營養和能量,所以大鼠的變化和饑餓5d相比并不太明顯,但饑餓9d時,“出現靜臥少動且行動遲緩,啃食鐵籠,目光呆滯,毛發灰黃稀長,行動無力,顫抖,弓背,步履蹣跚,體溫低,尾部掉屑,觸之冰涼感,體質量下降明顯”,很顯然,大鼠的能量已經是徹底的消耗殆盡,行將朽木了。所以短期饑餓時,自噬作用對于機體是保護性的,而在長期饑餓時,后期的自噬作用,可能也有保護性的成分,但綜合起來,應該是損傷性或是破壞性為主了。
表2 各組大鼠心肌LC3和Beclin1的 Western blot檢測結果 (±s,n=6)
注:與對照組比較,aP<0.05
1KimI,Rodriguez-EnriquezS,LemastersJJ.Selectivedegradationofmitochondriabymitophagy[J].ArchBiochemBiophys,2007,462(2):245-253.
2LevineB,KlionskyDJ.Developmentbyself-digestion:molecularmechanismsandbiologicalfunctionsofautophagy[J].DevCell,2004,6(4):463-477.
3MizushimaN,LevineB,CuervoAM,etal.Autophagyfightsdiseasethroughcellularself-digestion[J].Nature,2008,451(7182):1069-1075.
4RavikumarB,FutterM,JahreissL,etal.Mammalianmacro-autophagyataglance[J].JCellSci, 2009,L22(Ptll): 1707-1711.
5HeCC,KlionskyDJ.Regulationmechanismsandsignalingpathwaysofautophagy[J].AnnuRevGenet,2009,43:67-93.
6NakaiA,YamaguchiO,TakedaT,etal.Theroleofautophagyincardiomyocytesinthebasalstateandinresponsetohemodynamicstress[J].NatMed,2007,13(5):619-624.
7KanamoriH,TakemuraG,GotoK,etal.Autophagylimitsacutemyocardialinfarctioninducedbypermanentcoronaryarteryocclusion[J].AmJPhysiolHeartCircPhysiol,2011,300(6):H2261-H2271.
8KanamoriH,TakemuraG,GotoK,etal.Theroleofautophagyemerginginpostinfarctioncardiacremodelling[J].CardiovascRes,2011,91(2):330-339.
9KanamoriH,TakemuraG,MaruyamaR,etal.Functionalsignificanceandmorphologicalcharacterizationofstarvation-inducedautophagyintheadultheart[J].AmJPathol,2009,174(5):1705-1714.
10RokeachLA,JbelM,DuludeD.Anotherfaceofcelldeath[J].CellCycle,2014,13(2):181-182.
11ZhuH,TannousP,JohnstoneJL,etal.Cardiacautophagyisamaladaptiveresponsetohemodynamicstress[J].JClinInvest,2007,117(7):1782-1793.
12 劉斌,孫靜,張晉霞,等.自噬及自噬相關蛋白在帕金森病模型大鼠黑質紋狀體中的表達及意義[J].中國神經免疫學和神經病學雜志,2014,21(3):187-191.
13KabeyaY,MizushimaN,UenoT,etal.LC3,amammalianhomologueofyeastApg8p,islocalizedinautophagosomemembranesafterprocessing[J].EMBOJ,2000,19(21):5720-5728.
14MizushimaN.Methodsformonitoringautophagy[J].IntJBiochemCellBiol,2004,36(12):2491-2502.
15TermanA,BrunkUT.Autophagyincardiacmyocytehomeostasis,aging,andpathology[J].CardiovascRes,2005,68(3):355-365.
《疑難病雜志》可直接使用的英文縮略語
T(體溫)R(呼吸)HR(心率)BP(血壓)
SBP(收縮壓)DBP(舒張壓)ECG(心電圖)EEG(腦電圖)
Hb(血紅蛋白)RBC(紅細胞)WBC(白細胞)N(中性粒細胞)
L(淋巴細胞)M(單核細胞)E(嗜酸粒細胞)PLT(血小板)
ESR(紅細胞沉降率)HCT(紅細胞壓積)TG(三酰甘油)TC(總膽固醇)
CH(膽固醇)LDL-C(低密度脂蛋白膽固醇)HDL-C(高密度脂蛋白膽固醇)SCr(血肌酐)
UA尿酸NO(一氧化氮)BUN(血尿素氮)T3(三碘甲狀腺原氨酸)
T4(甲狀腺素)ALT(丙氨酸氨基轉移酶)AST(天冬氨酸氨基轉移酶)APTT(活化部分凝血活酶時間)
PaO2(動脈血氧分壓)PaCO2(動脈血二氧化碳分壓)SaO2動脈血氧飽和度IU或U(酶活力的國際單位)
The expression and significance of autophagy gene LC3, Beclin 1 in cardiac muscle of rat model with starvation
ZHANGJiankai*,ZHANGXiaoping,CHUYingxin.*DepartmentofAnesthesiology,NorthChinaUniversityofScienceandTechnologyAffiliatedHospital,Tangshan063000,China
ZHANGXiaoping,E-mail:xpzh55@sohu.com
Objective To observe the expression of microtubule associated protein 1 (LC3) and autophagy gene Beclin 1 in rat cardiac myocytes, and to investigate the effect of autophagy on the myocardium of starved rats. Methods 42 healthy male SD rats were randomly divided into normal control group (n=6)andstarvationmodelgroup(n=36).Normalcontrolgroupwithnormaldiet,starvationmodelgroupwereonlywithfreewaterintake,accordingtothenumberofdaysofthestarvation,theyweredividedinto1d, 2d, 3d, 5d, 7d, 9dsubgroups(n=6).Thecardiacspecimenswereextractedandretained,andthelevelsofautophagyproteinLC3andBeclin1weredetectedbyimmunohistochemistryandWesternblotting.Results Compared to normal control group, starvation model group’s SD rat cardiac myocyte autophagy protein LC3 and Beclin 1 positive cell number and protein expression increased in different degree, and the differences were statistically significant (P<0.01orP<0.05),andtherewerestatisticallysignificantdifferencesofautophagyproteinLC3andBeclin1positivecellnumberandproteinexpressionbetweendifferentsubgroupofstarvationmodelgroup(P<0.01orP<0.05),andthereweretwopeaksat2dand7d.Conclusion Starvation involved in the mechanism of myocardial pathological changes of myocardial process by activation of autophagy.
Autophagy gene; Beclin 1; Microtubule associated protein 1 light chain; Myocardium; Starvation; Rats
063000 唐山,華北理工大學附屬醫院麻醉科研究生[(張建凱(現在唐山市第二醫院工作)、褚迎欣],麻醉科(張小平)
張小平,E-mail:xpzh55@sohu.com
2015-03-19)
【DOI】 10.3969 /j.issn.1671-6450.2015.07.018
猜你喜歡
童話王國·奇妙邏輯推理(2024年5期)2024-06-19 16:03:38
網絡安全與數據管理(2022年1期)2022-08-29 03:15:20
導航定位學報(2022年4期)2022-08-15 08:27:00
中學生數理化·中考版(2022年8期)2022-06-14 06:55:24
新世紀智能(數學備考)(2021年9期)2021-11-24 01:14:36
成都醫學院學報(2021年2期)2021-07-19 08:35:14
新世紀智能(數學備考)(2020年9期)2021-01-04 00:25:14
中學生數理化·七年級數學人教版(2020年10期)2020-11-26 08:24:50
數學物理學報(2020年2期)2020-06-02 11:29:24
光學精密工程(2016年6期)2016-11-07 09:07:19
