枯草芽孢桿菌BJ3-2 S-腺苷甲硫氨酸脫羧酶基因speD的克隆與序列分析
2015-04-23 08:58:52王亞娟韓忠安羅信旭吳擁軍
中國釀造 2015年1期
王亞娟,韓忠安,羅信旭,吳擁軍*
(貴州大學 生命科學學院,貴州 貴陽 550025)
生物胺(biogenic amines,BA)是一類主要由氨基酸脫羧或醛和酮氨基化形成的弱堿性低分子質量含氮化合物[1],是人體內的正常生理活性物質,但當體內攝入過量生物胺時會引起頭痛、心悸等一系列不良反應。研究發現,生物胺還具有潛在的致癌性,生物胺存在于食品中也會影響食品風味甚至改變其成分風味[2-3]。S-腺苷甲硫氨酸脫羧酶(S-adenosylmethionine decarboxylase,SAMDC 或AdoMet-DC)是多胺合成代謝中的3個關鍵酶之一[4-5]。其存在于細菌、真菌、古生菌和植物體內,在多胺生物合成過程中起著重要的作用,它以S-腺苷甲硫氨酸(S-adenosylmethionine,SAM)為底物催化形成脫羧S-腺苷甲硫氨酸,為精胺和亞精胺的合成提供氨丙基,是精胺和亞精胺合成的限速酶[6-7]。近年已有人類及多種植物的SAMDC 基因被分離鑒定,并證明其在多胺合成中具有重要作用[8]。食品中主要存在的生物胺包括組胺、腐胺、尸胺、酪胺、色胺、β-苯乙胺、精胺和亞精胺。
豆豉是我國傳統發酵豆制品,經微生物發酵以色香誘人、風味獨特而深受消費者的喜愛。目前已報道的貴州地區水豆豉主酵微生物是枯草芽孢桿菌(Bacillus subtilis),是細菌型豆豉生產的安全菌株[9]。據報道,與枯草芽孢桿菌同屬的納豆芽孢桿菌發酵過程中檢測到了亞精胺、精胺、腐胺、酪胺4種生物胺[10-11],但豆豉發酵過程中是否會產生過量的生物胺還不得而知。枯草芽孢桿菌在發酵過程中分泌的脫羧酶能夠將氨基酸經過脫羧反應生成相應的胺類物質[10]。在前期的研究中,實驗室已從貴州地區豆豉樣品中分離獲得了可作為生產應用的菌株BJ3-2[12],以Bacillus subtilisBJ3-2基因組為模板,克隆S-腺苷甲硫氨酸脫羧酶(SAMDC)基因(speD),并與已報道的枯草芽孢桿菌的speD序列進行同源性比對分析,推測SAMDC活性結構域和酶活性位點,研究為SAMDC活性影響因素的進一步研究和豆豉生產中的安全評價奠定了基礎。
1 材料與方法
1.1 材料與試劑
枯草芽孢桿菌(B.subtilis)BJ3-2:實驗室保藏菌株;E.coliDH5α感受態細胞:本實驗室經氯化鈣法制備;質粒pGEM-T載體:美國Promega公司。
T4 DNA連接酶、Taq(聚合)酶、DNA Marker、dNTP、10×PCR Buffer等:大連TaKaRa公司;基因組提取試劑盒:北京Promega生物技術有限公司;質粒提取試劑盒、膠回收試劑盒:美國OMEGA生物技術公司;其他試劑均為國產分析純。
1.2 儀器與設備
MyCycler PCR儀、PowerPac HC電泳儀、Gel Doc XR型UniversalHood凝膠成像系統:美國Bio-Rad公司;DHP-9162型電熱恒溫培養箱:上海一恒科技有限公司;SKY-2102C型恒溫搖床:上海蘇坤實業有限公司;SW-CJ-IFD型標準型凈化工作臺:蘇州凈化設備工程有限公司。
1.3 方法
1.3.1 引物設計與合成
查詢GenBank獲得B.subtilis168菌株的speD序列,應用Primer 5.0軟件設計speD特異性克隆引物。結果見表1。
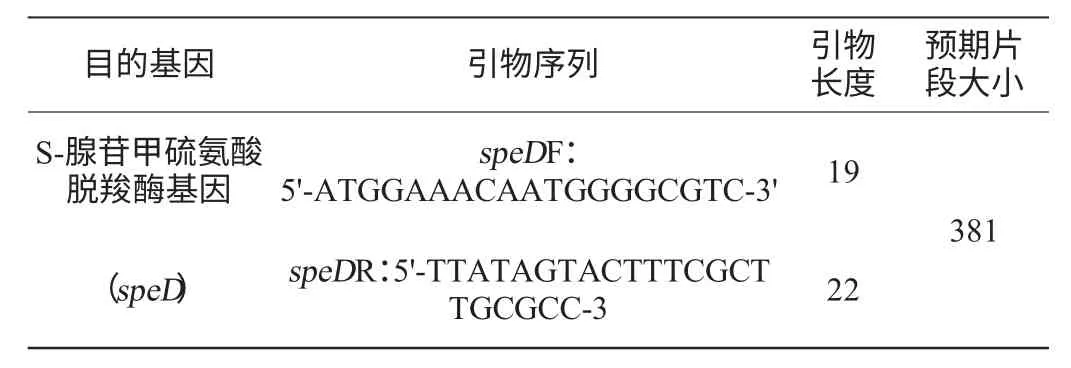
表1 擴增目的基因的名稱及引物序列Table 1 Name of target gene amplified and its primer sequence
1.3.2B.subtilisBJ3-2基因組提取、質粒提取
依照試劑盒說明書進行;片段回收、連接、轉化參照分子克隆實驗指南第三版。
1.3.3 PCR擴增
以B.subtilisBJ3-2的gDNA為模板,引物speDF/speDR進行PCR擴增,反應體系:模板1 μL、dNTP 1.6 μL、10×PCR Buffer 2 μL、上下游引物各0.5 μL、Taq酶0.1 μL、ddH2O 14.3 μL,總體系為20 μL。
1.3.4 重組子鑒定
經菌落PCR和質粒PCR驗證,重組質粒送北京諾賽基因組研究中心有限公司測序。
1921年中國共產黨成立后就立即研究在各地建立和發展社會主義青年團,并派出許多黨員去加強對各地團的早期組織的領導工作。1922年5月,中國社會主義青年團第一次全國代表大會開幕。大會通過的團的綱領確定了中國社會主義青年團為中國青年無產階級的組織,是為解放無產階級而奮斗的組織。此后十幾年間,共青團組織在黨的領導下,帶領各地的團員青年相繼投身工人運動、改組國民黨、北伐戰爭、反對國民黨統治的斗爭中去。抗日戰爭爆發后,為了建立抗日民族統一戰線,更廣泛地團結各界青年投入抗日救亡的斗爭,共產主義青年團響應黨的號召,主動改造變身為各種抗日救國的青年團體,為抗戰的勝利做出了突出貢獻。
1.3.5speD序列分析
用DNAStar和MEGA5生物學軟件對speD的核苷酸序列及推導的氨基酸序列進行編輯,與GenBank中已發表的其他speD序列進行同源性分析并繪制SAMDC遺傳進化樹[14]。使用Clustalw2和GeneDoc生物學軟件進行多序列比對分析。
2 結果與分析
2.1 B.subtilis BJ3-2g DNA的提取
提取B.subtilisBJ3-2基因組,樣品經瓊脂糖凝膠電泳檢測,結果見圖1。圖1顯示所提取的B.subtilisBJ3-2 gDNA較完整,無顯著降解,可用于目的基因的PCR擴增。
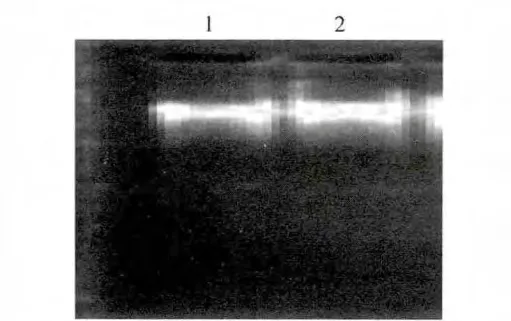
圖1 枯草芽孢桿菌BJ3-2基因組Fig.1 Genome of Bacillus subtilis BJ3-2
2.2 speD的PCR擴增
將克隆產物經瓊脂糖凝膠電泳檢測,結果見圖2。由圖2可知,目的條帶約與預期片段(381 bp)相符。
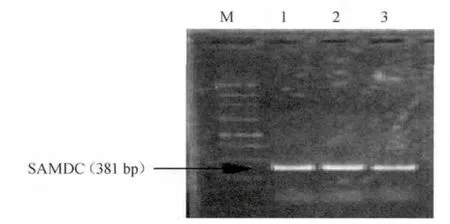
圖2 PCR擴增speD基因Fig.2 PCR amplification of speD
2.3 重組克隆質粒的菌落PCR鑒定
挑取單菌落,經菌落PCR檢測,結果見圖3。由圖3可知,2、4、6、7泳道在預期大小處有克隆條帶,初步認定為陽性重組菌。
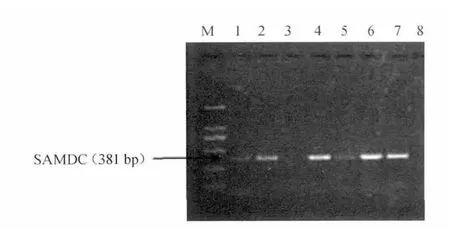
圖3 菌落PCR鑒定重組子Fig.3 PCR recombinant identification of colony
2.4 重組質粒PCR鑒定
將鑒定的陽性重組菌落4進行增菌培養,提取質粒,進行質粒PCR檢測,結果見圖4。由圖4可知,在預期大小處出現克隆條帶,說明目的基因已正確克隆至pGEM-T 載體中。將提取的重組質粒送生物公司測序,并命名為pGEM-speD。
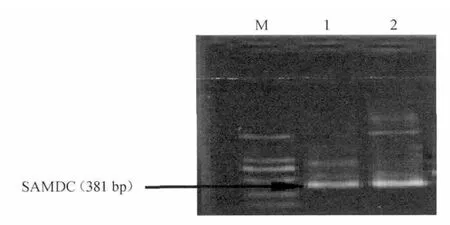
圖4 質粒PCR鑒定Fig.4 PCR identification of plasmid
2.5 speD的同源性分析及系統進化樹構建
在GenBank數據庫中將所克隆基因序列和推導的蛋白序列進行序列同源性比對,分析結果表明,B.subtilisBJ3-2的speD序列與已報道枯草芽孢桿菌的speD基因序列和蛋白序列高度同源,與萎縮芽孢桿菌(Bacillus atrophaeus)的核苷酸序列相似性達96%,與解淀粉芽孢桿菌(Bacillus amyloliquefaciens)達93%以上,與其他芽孢桿菌在85%以上。其氨基酸序列與芽孢桿菌NSP9.1(Bacillussp.NSP9.1)同源性達到97%、與印度芽孢桿菌(Bacillus indicus)LMG 22858同源性達到95%、與索諾拉沙漠芽孢桿菌(Bacillus sonorensis)達到96%,與芽孢桿菌屬其他多種菌種的SAMDC氨基酸序列在90%以上。表明所克隆的基因是編碼SAMDC的基因。
所有生物的腺苷甲硫氨酸脫羧酶都是依賴丙酮酰基的酶,依據其剪切位點、結構特征和酶活性離子,將之分為兩大類:Ⅰ類酶主要存在于細菌和古細菌;Ⅱ類酶主要存在于真核生物中。SAMDC活性酶是從來源于絲氨酸自我剪切而產生的具有催化活性的丙酮酰基修飾產生的,盡管兩類酶的形成都是利用丙酮酰基,經歷同樣的自身成熟過程,但是除了丙酮酰基位點外Ⅰ類和Ⅱ類酶幾乎沒有序列相似性[15]。
Ⅰ類酶可分為兩類,Ⅰ類A型:SAMDC主要包括革蘭氏陰性細菌,活性酶的形成需要二價金屬離子(如Mg2+)的參與催化形成(αβ)4四聚物;Ⅰ類B型:SAMDC包括革蘭氏陽性細菌和古細菌,如桿狀菌和詹氏甲烷球菌,形成(αβ)2二聚物,并且不需要鎂離子或其他的活性劑參與[16-17]。
Ⅱ類酶主要包括來自于動物、植物、真菌和錐體蟲(無脊椎)的腺苷甲硫氨酸脫羧酶。Ⅱ類A包括來自于動物的S-腺苷甲硫氨酸脫羧酶(AdoMetDC),是(αβ)2二聚物和需要腐胺作為激活因子。Ⅱ類B包括來自于植物的AdoMetDC,是(αβ)單體和不需要腐胺作為激活因子。Ⅱ類C包括來自于錐體蟲(無脊椎)的AdoMetDC異二聚體,具有αβπ′亞基結構。鐮狀瘧原蟲的AdoMetDC 是一種雙功能蛋白,C末端具有鳥氨酸脫羧酶活性。
研究發現,枯草芽孢桿菌的SAMDC與古生菌的SAMDC密切相關。B.subtilisBJ3-2與B.subtilis168屬于枯草芽孢桿菌,詹氏甲烷球菌(Methanocaldococcus jannaschii)和嗜熱泉生古細菌(Aeropyrum pernixK1)屬于古生菌,海棲熱孢菌(Thermotoga maritima)都屬于Ⅰ類B型AdoMetDC,且海棲熱孢菌(T.maritima)的AdoMetDC的晶體結構的構建為Ⅰ類B型AdoMetDC(PDB code 1TLU)的酶活性以及結構的分析提供了理論基礎[17]。選擇這些細菌的SAMDC的氨基酸序列作比對,推測B.subtilisBJ3-2的SAMDC酶原剪切位點及酶活性位點。根據大腸桿菌(Escherichia coli)和人類(Homo sapiens)、擬南芥(Arabidopsis thaliana)的SAMDC氨基酸序列以及同源性較高的芽孢桿菌SAMDC序列構建系統進化樹,進一步分析它們之間的進化關系。
ClustalW多序列比對,GeneDoc輸出,圖5結果顯示B.subtilisBJ3-2的SAMDC基因屬于丙酮酸依賴型脫羧酶的Ⅰ類B型S-腺苷甲硫氨酸脫羧酶家族,具有酶原剪切結構域:(53 GVSGVVIISESHLTIH 68)結構域,其中ESH三個氨基酸殘基高度保守,包括有丙酮酰前體絲氨酸和酶原剪切位點;第二個保守結構域:(82 TCG 84)結構域,在這個結構域中CG兩個氨基酸殘基高度保守,具有親核催化作用的半胱氨酸殘基,對酶的催化活性具有重要作用。

圖5 B.subtilis BJ3-2和其他細菌SAMDC的氨基酸多序列比對Fig.5 Multiple alignments between amino acid sequence of B.subtilis BJ3-2 and other bacteria
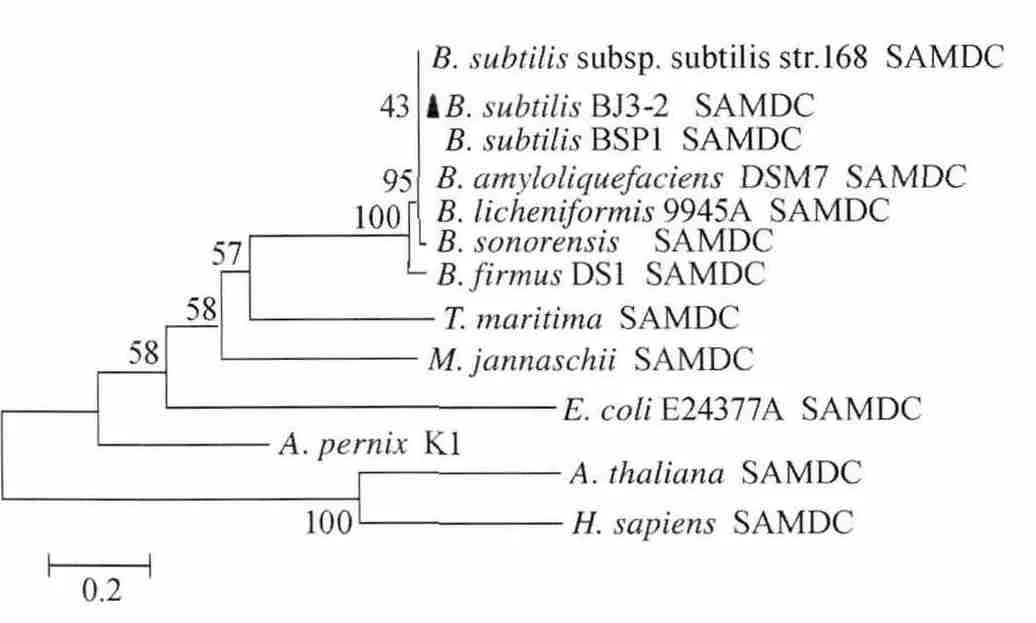
圖6 不同類型S-腺苷甲硫氨酸脫羧酶的氨基酸序列遺傳進化樹Fig.6 Phylogenetic tree constructed with amino acid sequences of different types of S-adenosylmethionine decarboxylase
從圖6可以看出,枯草芽孢桿菌(B.subtilis)的SAMDC位于遺傳距離較近的同一進化分支,彼此親緣關系較近,B.subtilisBJ3-2與B.subtilis168和B.subtilisBSP1的SAMDC遺傳距離最近,詹氏甲烷球菌(M.jannaschii)、嗜熱泉生古細菌(A.pernix)K1 和海棲熱孢菌(T.maritima)的SAMDC遺傳距離次之,與E.coli[A7ZHK9]的SAMDC位于遺傳距離稍遠的同一進化分支,與人類(H.Sapiens)[NP_001625]、擬南芥(A.thaliana)[AEE73813]的SAMDC遺傳距離最遠。以上分析顯示B.subtilisBJ3-2 的SAMDC具有丙酮酸依賴型脫羧酶酶原剪切結構域,屬于典型的Ⅰ類B型SAMDC家族成員。
3 結論
SAMDC是多胺合成代謝過程中的限速酶,是精胺和亞精胺合成的關鍵酶,本實驗依據GenBank中枯草芽孢桿菌168菌株(登錄號為AL009126.3)的speD編碼基因序列,設計了特異性引物,通過PCR技術從B.subtilisBJ3-2菌株中擴增得到SAMDC基因序列,與預期大小相吻合。通過序列比對、保守結構域分析,B.subtilisBJ3-2 的SAMDC具有丙酮酸依賴型脫羧酶酶原剪切結構域[53 GVSGVVIISESHLTIH 68]和保守結構域[82 TCG 84],屬于典型的的Ⅰ類B型S-腺苷甲硫氨酸脫羧酶家族成員,其關鍵的活性位點氨基酸殘基,即絲氨酸(Ser,S)Ser55、絲氨酸Ser63、組氨酸(His,H)His68、半胱氨酸(Cys,C)Cys83,與底物結合、酶的催化及酶原的加工過程相關。
在革蘭氏陽性菌枯草芽孢桿菌中其多胺的合成是始于精氨酸,由精氨酸脫羧酶、胍基丁胺酶、S-腺苷甲硫氨酸脫羧酶以及亞精胺合酶、精胺合酶共同參與作用的。所以應同時對這幾個基因進行研究,以進一步了解枯草芽孢桿菌生物多胺的合成途徑。在后續實驗中可以選擇合適的表達載體進行蛋白表達純化,對其酶活力和酶學性質進行研究。綜合細菌型豆豉生物發酵特點,使得氨基酸脫羧酶既能滿足豆豉發酵條件的需要又能有效控制豆豉產品中生物胺的含量,為有效保證豆豉產品的生物安全性提供理論指導。
[1]何慶華,吳永寧,印遇龍.食品中生物胺研究進展[J].中國食品衛生雜志,2007,19(5):451-454.
[2]曲映紅.發酵食品中生物胺的研究進展[J].食品工業,2012(3):115-118.
[3]劉 景,任 婧,孫克杰.食品中生物胺的安全性研究進展[J].食品科學,2013,35(5):322-326.
[4]汪 天,孟艷瓊,谷 鳳,等.多胺代謝及其與農產品貯藏保鮮關系的研究[C]//2005 安徽食品安全博士論壇,合肥,2005.
[5]耿衛東,李艷軍,張新宇,等.棉花S-腺苷甲硫氨酸脫羧酶基因的克隆及低溫下的表達分析[J].作物學報,2012,38(9):1649-1656.
[6]GALSTON A W,SAWHNEY R K.Polyamines in plant physiology[J].Plant Physiology,1990,94(2):406-410.
[7]劉志勇,王孝宣,高建昌,等.番茄S-腺苷蛋氨酸脫羧酶基因SlSAMDC1的克隆與序列分析[J].園藝學報,2008,35(8):1137-1146.
[8]路玉蘭,孫艷香,馮 雪,等.百脈根S-腺苷甲硫氨酸脫羧酶基因克隆與表達分析[J].華北農學報,2013,28(2):78-85.
[9]吳擁軍,孟望霓,李耀中.純種強化發酵細菌型豆豉研究[J].中國釀造,2011,30(9):56-60.
[10]蘇 悟,鄭小芬,徐睿烜,等.1 種細菌型豆豉自然發酵過程中生物胺的變化[J].食品與發酵工業,2014,40(7):40-45.
[11]張建華,沈 翔,于湘莉.納豆發酵過程中的生物胺[J].上海交通大學學報,2007,25(1):2-5.
[12]賈東旭,吳擁軍,李耀中,等.細菌型豆豉發酵芽孢桿菌的篩選與鑒定[J].食品科學,2009,30(5):217-221.
[13]J.薩姆布魯克,D.W 拉塞爾.黃培堂,等譯.分子克隆實驗指南(第三版)[M].北京:科學出版社,2002.
[14]張麗娜,榮昌鶴,何 遠,等.常用系統發育樹構建算法和軟件鳥瞰[J].動物學研究,2013,34(6):640-650.
[15]SEKOWSKA A,COPPEE J Y,LE CAER J P,et al.S-adenosylmethionine decarboxylase ofBacillus subtilis is closely related to archaebacterial counterparts[J].Mol Microbiol,2000,36(5):1135-1147.
[16]張佳景,丁淑麗,鄒宜靜,等.植物腺苷甲硫氨酸脫羧酶研究進展[J].細胞生物學雜志,2008,30(5):622-628.
[17]BALE S,EALICK S E.Structural biology of S-adenosylmethionine decarboxylase[J].Amino Acids,2010,38(2):451-460.
