PGP9.5和神經肽Y在雙峰駝正常睪丸和隱睪的分布比較
2015-03-22 09:08:11袁莉剛閆振龍陶金忠陳國娟
畜牧獸醫學報 2015年2期
袁莉剛,閆振龍,陶金忠,劉 英,陳國娟
(1.甘肅農業大學動物醫學院,蘭州 730070 ;2.甘肅農業大學生命科學技術學院,蘭州 730070;3.寧夏大學 動物醫學院,銀川 750021)
PGP9.5和神經肽Y在雙峰駝正常睪丸和隱睪的分布比較
袁莉剛1*,閆振龍2,陶金忠3,劉 英1*,陳國娟1
(1.甘肅農業大學動物醫學院,蘭州 730070 ;2.甘肅農業大學生命科學技術學院,蘭州 730070;3.寧夏大學 動物醫學院,銀川 750021)
探討PGP9.5和神經肽Y在雙峰駝正常睪丸和隱睪的表達及在精子發生中的作用機制。采集性成熟未交配2~3歲成年雙峰駝正常睪丸及隱睪,應用免疫組織化學SP法檢測PGP9.5和神經肽Y在睪丸的定位,并通過圖像分析技術進行定量分析。結果表明:PGP9.5在正常睪丸支持細胞、各級生精細胞和動靜脈血管都有高密度陽性反應;神經肽Y在支持細胞呈中等陽性,各級生精細胞和動靜脈血管呈弱陽性,隱睪組中PGP9.5和神經肽Y表達位置相似于正常組,但表達量顯著降低。PGP9.5和神經肽Y在正常組和隱睪組Leydig細胞表達無差異,均為強陽性。可見PGP9.5及神經肽Y參與了雙峰駝正常睪丸和隱睪生精功能的調節,二者通過支持細胞及管周肌樣細胞對于隱睪生精微環境的調控能力降低,但隱睪內間質細胞的分泌并未受明顯影響。本研究為進一步研究雙峰駝睪丸神經遞質變化與隱睪癥發生關系及調控機制提供了參考。
雙峰駝;隱睪;PGP9.5;神經肽Y;免疫組織化學
哺乳動物睪丸功能主要受到交感神經和局部激素分泌的調節。蛋白基因產物9.5(protein gene product,PGP 9.5)由212個氨基酸組成,相對分子質量27 ku,作為一種泛神經標記物廣泛表達于神經元分化的各個階段,應用于器官和組織的發病機制和診斷研究[1]。神經肽Y(neuropeptide Y,NPY)由36個氨基酸組成,是神經系統中最豐富的短肽,具有參與激素分泌,調節生長發育、性行為等生物學功能[2]。研究表明,神經肽Y通過參與調控下丘腦—促性腺激素釋放激素(gonadotropin-releasing hormone,GnRH) 和垂體促性腺激素(Pituitary-gonadotropin)的釋放,調節睪丸生理功能。國內外研究顯示,PGP9.5和神經肽Y在人、豬、驢、單峰駝等哺乳動物睪丸內有顯著分布,在睪丸正常生理功能方面發揮著重要的調節作用[3-6]。哺乳動物下丘腦—垂體—睪丸內分泌軸和睪丸組織局部自分泌及旁分泌調節系統參與精子發生調控,睪丸受膽堿能神經、去甲腎上腺素能神經和肽能神經支配,神經遞質水平變化對正常生精過程必不可少,且與不育癥發生關系密切[7-8]。雙峰駝為中國遺傳資源保護品種,目前關于雙峰駝隱睪神經調控未見報道。本研究通過雙峰駝睪丸內PGP9.5和神經肽Y分布比較正常睪丸與隱睪內神經遞質的分布變化,對雙峰駝遺傳資源保護以及從神經方面治療隱睪癥具有重要意義。
1 材料與方法
1.1 材料與試劑
2012年12月,自寧夏回族自治區吳忠市澇河橋清真牛羊肉批發市場,采集健康性成熟未交配2~3歲雙峰駝。經頸部放血致死后立即采集睪丸組織標本,共20例,其中正常睪丸14例,隱睪6例。以4%中性甲醛溶液固定3 d后,制作石蠟切片,常規染色、觀察。正常組選擇組織結構發育良好的睪丸;隱睪組選擇結構典型的歸為一組,進一步分別進行免疫組織化學研究。NPY兔抗鼠多克隆抗體(bs-0071R)和PGP9.5兔抗鼠多克隆抗體(bs-3806R)購自北京博奧森生物技術有限公司;免疫組化染色試劑盒(sp-0023,由美國ZYMED公司生產)購自北京博奧森生物技術有限公司;DAB顯色試劑盒(ZLI-9018)和APES防脫玻片(ZLI-9502)購自北京中杉金橋生物技術有限公司。
1.2 方法
采用免疫組織化學SP法進行染色,NPY和PGP9.5兩種多克隆抗體均1∶400稀釋后使用。陰性對照組用0.01 mol·L-1的PBS代替一抗(NPY和PGP9.5)。用OlympusDP71顯微鏡進行觀察并照相。
1.3 圖像分析
光鏡下觀察免疫組織化學切片,每組切片中隨機選取5張,在400倍和1 000倍顯微鏡下進行拍照,對400倍照片用DP軟件采集5個視野,首先參照A.M.Suburo等[9]采用半定量的形式對染色結果的分布密度進行描述,-,無陽性表達;+/-,偶有陽性表達;+,陽性表達;++,中等強度陽性表達;+++,強陽性表達;++++,高密度強陽性。然后用Image-Pro Plus6.0軟件進行圖像分析,測定陽性反應物的平均光密度,用SAS(1996)統計軟件中GLM模型,以ANOVA方法對數據進行分析。若各處理間差異顯著,則用Duncan’s多重比較進行檢驗。結果用最小二乘法的均值及平均標準誤表示,P<0.05為差異顯著,P<0.01為差異極顯著。
2 結 果
PGP9.5和神經肽Y在雙峰駝正常睪丸和隱睪表達的免疫組織化學染色見圖1、2。
2.1 PGP9.5在雙峰駝正常睪丸和隱睪的定位及分布特點
雙峰駝正常睪丸組織中PGP9.5肽能神經分布廣泛,結果顯示血管周圍、睪丸間質、曲精小管以及周邊均有不同程度分布(圖1a、b、c)。PGP9.5在支持細胞為高密度強陽性,精原細胞為強陽性,初級精母細胞和精子細胞為高密度強陽性,偶有中等陽性,管周肌樣細胞為中等陽性,Leydig細胞為強陽性,間質組織中偶有少量PGP9.5中等陽性的神經纖維分布。各級動脈血管管壁和其內皮細胞以及各級靜脈血管的內皮細胞PGP9.5為強陽性(表1)。在 14例正常組睪丸PGP9.5肽能神經的免疫組化結果顯示一致。
隱睪組織中PGP9.5肽能神經免疫陽性表達位置與正常組相似(圖1e、f、g),但在表達程度上低于正常組,組間差異極顯著(圖3,P<0.01)。PGP9.5在隱睪支持細胞和初級精母細胞為強陽性,精原細胞為中等陽性,管周肌樣細胞為陽性。Leydig細胞為中等陽性,間質組織中有少量陽性神經纖維分布。各級動脈血管和靜脈血管管壁PGP9.5為中等陽性(表1)。 6例隱睪組睪丸內PGP9.5肽能神經分布結果顯示一致。
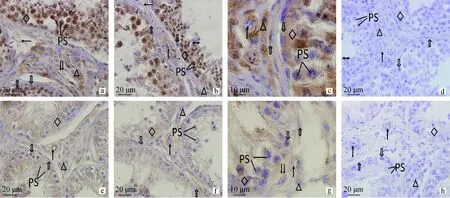
PS.初級精母細胞;←.神經纖維;△.Leydig細胞;↑.動脈血管;↓↓.靜脈血管;.肌樣細胞;.精原細胞;◇.支持細胞;?.精子。a、b.正常組,標尺20 μm;c.正常組,標尺10 μm;d.正常組對照,標尺20 μm;e、f.隱睪組,標尺20 μm;g.隱睪組,標尺10 μm;h.隱睪組對照,標尺20 μmPS.Primary spermatocytes;←.Nerve fibers;△.Leydig cells;↑.Arterial wall;↓↓.Vein wall;.Myoid peritubular cells;.Spermatogonia;◇.Sertoli cells;?.Sperm.a,b.The normal Bactrian camel testis,Bar=20 μm;c.The normal Bactrian camel testis,Bar=10 μm;d.The control of the normal Bactrian camel testis,Bar=20 μm;e,f.The cryptorchidism Bactrian camel testis,Bar=20 μm;g.The cryptorchidism Bactrian camel testis,Bar=10 μm;h.The control of the cryptorchidism Bactrian camel testis,Bar=20 μm圖1 PGP9.5在雙峰駝正常睪丸和隱睪的分布(免疫組化染色)Fig.1 Expression of PGP 9.5 in the normal and cryptorchidism Bactrian camel testis (immunohistochemical staining)
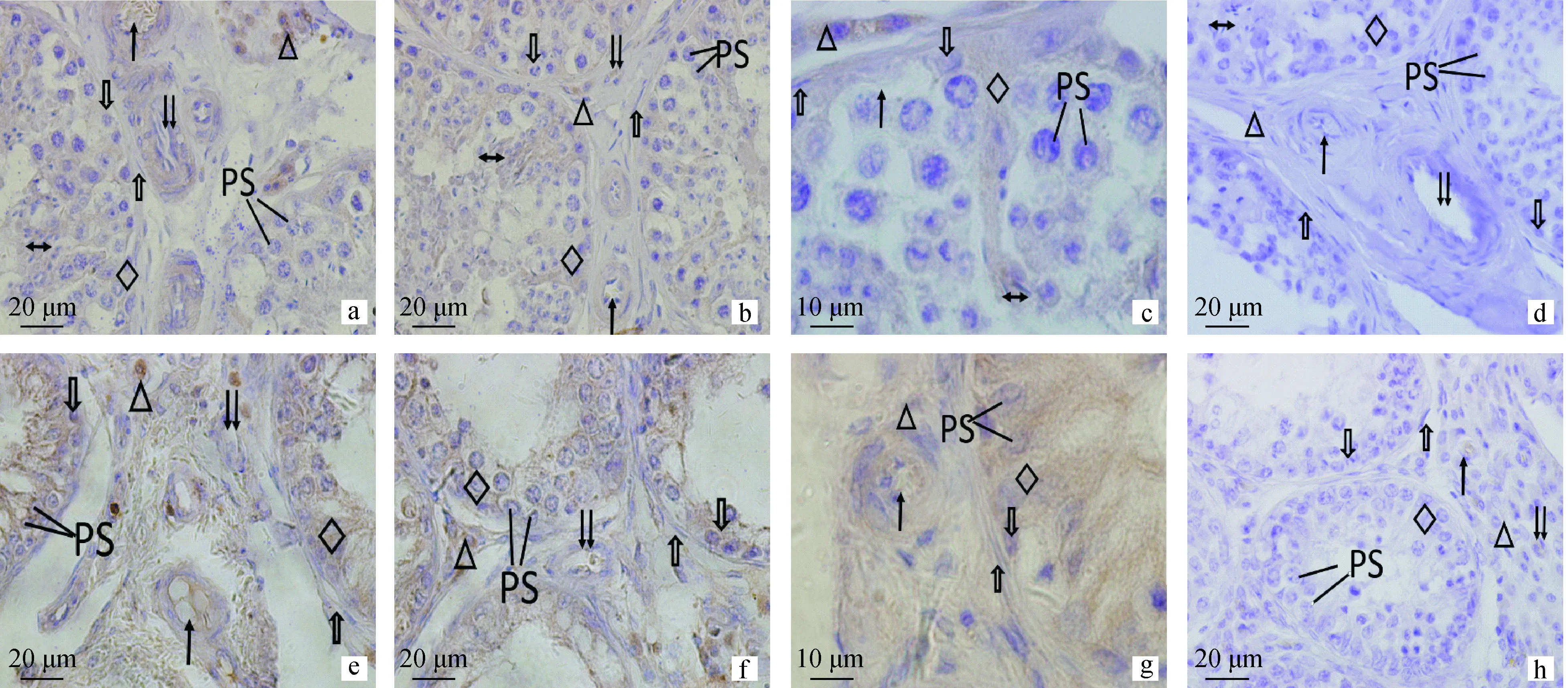
PS.初級精母細胞;←.神經纖維;△.Leydig細胞;↑.動脈血管;↓↓.靜脈血管;.肌樣細胞;.精原細胞;◇.支持細胞;?.精子。a、b.正常組,標尺20 μm;c.正常組,標尺10 μm;d.正常組對照,標尺20 μm;e、f.隱睪組,標尺20 μm;g.隱睪組,標尺10 μm;h.隱睪組對照,標尺20 μmPS.Primary spermatocytes;←.Nerve fibers;△.Leydig cells;↑.Arterial wall;↓↓.Vein wall;.Myoid peritubular cells;.Spermatogonia;◇.Sertoli cells;?.Sperm.a,b.The normal Bactrian camel testis,Bar=20 μm;c.The normal Bactrian camel testis,Bar=10 μm;d.The control of the normal Bactrian camel testis,Bar=20 μm;e,f.The cryptorchidism Bactrian camel testis,Bar=20 μm;g.The cryptorchidism Bactrian camel testis,Bar=10 μm;h.The control of the cryptorchidism Bactrian camel testis,Bar=20 μm圖2 NPY在雙峰駝正常睪丸和隱睪的分布(免疫組化染色)Fig.2 Expression of NPY in the normal and cryptorchidism Bactrian camel testis (immunohistochemical staining)
2.2 神經肽Y在雙峰駝正常睪丸和隱睪的定位及分布特點
雙峰駝正常睪丸組織中神經肽Y的表達主要定位在各級動靜脈血管管壁、Leydig細胞和支持細胞(圖2a、b、c)。結果顯示,神經肽Y在支持細胞為強陽性,精原細胞、初級精母細胞和次級精母細胞為中等陽性,精子細胞為中等陽性。管周肌樣細胞呈陽性表達。Leydig細胞為中等陽性,偶有強陽性。各級動靜脈為陽性,靜脈血管陽性表達較動脈血管弱(表1)。14例正常組睪丸內神經肽Y分布結果顯示一致。
雙峰駝隱睪組中支持細胞排列散亂,而且無明顯界限(圖2e、f、g),神經肽Y在支持細胞為中等陽性。精原細胞胞膜界限不清楚,為弱陽性表達。管周肌樣細胞偶有陽性表達。Leydig細胞為中等強度表達,間質組織中存在少量陽性神經纖維。各級動靜脈血管管壁或內皮細胞神經肽Y為中等強度或陽性,且在毛細血管為陰性表達(表1)。PGP9.5和神經肽Y在雙峰駝隱睪不同部位分布密度與正常組之間組間差異極顯著(圖3,P<0.01)。6例隱睪組睪丸內神經肽Y分布結果顯示一致。
表1 PGP9.5和神經肽Y在雙峰駝睪丸正常組和隱睪組中不同部位的分布密度
Table 1 The density of PGP9.5 and NPY in different parts of the normal and cryptorchidism of Bactrian camel testis

組別Group支持細胞Sertolicells精原細胞Spermatogonia精子細胞SpermatidLeydig細胞Leydigcells管周肌樣細胞Myoidperitubularcells動脈血管壁Arterialwall靜脈血管壁VeinwallPGP9.5-正常組PGP9.5-Thenormalgroup++++++++++++++++++++++PGP9.5-隱睪組PGP9.5-Thecryptorchidismgroup++++++++++++NPY-正常組NPY-Thenormalgroup++++++++++++NPY-隱睪組NPY-Thecryptorchidismgroup++++++/-+++

標有不同字母的組間差異極顯著(P<0.01),標有相同字母的組間無差異(P>0.05)Different letters mean significant difference between the groups (P<0.01),the same letters mean no difference between groups (P>0.05)圖3 PGP9.5和神經肽Y在雙峰駝睪丸正常組和隱睪組中免疫組織化學表達Fig.3 The Immunohistochemistry expression of PGP9.5 and NPY in the normal and cryptorchidism of Bactrian camel testis
3 討 論
哺乳動物睪丸內分布著豐富的神經纖維,其主要來自精索上神經和精索下神經。國內外研究顯示諸多哺乳動物的睪丸內均有PGP9.5和神經肽Y的大量表達,A.Sainsbury等[10]研究發現PGP9.5在牛生精細胞、精原細胞以及精原母細胞均陽性表達;位蘭等[11]報道雛鴕鳥睪丸精母細胞神經肽Y陽性表達,表明PGP9.5和神經肽Y可能參與了睪丸生精功能調節。睪丸內神經對精子的發生有著重要的調節作用,S.H.Chow等及龔永光等[12-13]研究表明切除小鼠精索神經引起精原細胞凋亡。本研究結果顯示在雙峰駝睪丸中PGP9.5和神經肽Y在正常組各級生精細胞中表達較強,而在隱睪表達較弱,且組間差異極顯著(表1,P<0.01),表明PGP9.5及神經肽Y參與了雙峰駝正常睪丸和隱睪生精功能的調節,二者通過支持細胞及管周肌樣細胞對于隱睪生精微環境的調控能力降低。
血睪屏障(blood-testis barrier,BTB)主要由支持細胞的緊密連接,曲細精管的基膜及其間質組成,為精子的發生提供了一種特定的免疫微環境。血睪屏障的通透性改變可刺激機體產生抗體,進而損害生精上皮,致使睪丸生精功能降低或者丟失[14]。支持細胞是睪丸免疫微環境的主要參與者,可使攜帶自身特異抗原的生精細胞免受自身免疫系統的攻擊。除此之外,還為生精細胞提供必須的營養物質和生長因子以及物理支撐[15]。本研究中PGP9.5與神經肽Y在雙峰駝正常組和隱睪組睪丸支持細胞陽性表達,表明肽能神經對支持細胞具有調節作用。研究表明[16],摘除犬睪丸交感神經后引起曲細精管的退化和間質組織增生,進而影響精母細胞分裂和精子產生。隱睪組在支持細胞的表達顯著弱于正常組,提示肽能神經對支持細胞的調控能力減弱,但關于肽能神經分布減弱是否導致支持細胞形態結構的改變,仍需進一步研究。管周肌樣細胞一方面參與構成曲細精管的固有膜,另一方面通過分泌收縮素等因子調節精子發生和睪丸的發育[17]。研究表明,肌樣細胞運輸功能與肽能神經的調節有關[18-19],其形態及分泌成分的改變會影響生精小管局部微環境從而導致男性不育[20]。本研究中PGP9.5和神經肽Y在管周肌樣細胞的陽性表達,且正常組顯著高于隱睪組(表1,P<0.01),表明隱睪內PGP9.5和神經肽Y通過管周肌樣細胞對微環境的調控能力減弱。
睪丸Leydig細胞睪酮的分泌主要受下丘腦—垂體調控,神經肽Y參與調控下丘腦—GnRH的脈沖釋放,通過旁分泌或者通過釋放多種神經肽對Leydig細胞類固醇激素的合成進行調節。國內外研究發現,精索去神經支配或者電刺激精索神經均會引起睪酮水平異常[21-22]。本研究PGP9.5和神經肽Y在雙峰駝正常組和隱睪組中Leydig細胞均有顯著陽性表達,且無明顯組間差異,表明肽能神經對隱睪內間質細胞的分泌調控并未明顯改變。
睪丸神經遞質水平變化與不育癥發生關系密切,PGP9.5及神經肽Y在雙峰駝正常睪丸和隱睪的定位及分布特點比較表明, PGP9.5及神經肽Y參與了睪丸生精功能的調節,且對隱睪支持細胞及管周肌樣細胞的調控能力減弱,進而影響睪丸內部免疫微環境的調節,但是隱睪內睪酮的正常分泌并未受影響,本研究為進一步研究雙峰駝隱睪生精細胞發育微生態環境與神經調控機制提供了參考。
[1] 凌 麗,薛金偉.神經纖維標記物-蛋白基因產物9.5(PGP9.5)的研究概況[J].中國老年學雜志,2010,30(21):3226-3228. LIN L,XUE J W.Research overview of nerve fiber marker protein gene product 9.5 (PGP9.5) [J]ChineseJournalofGerontology,2010,30(21):3226-3228.(in Chinese)
[2] 王曉煒,崔毓桂,陳子慶,等.神經肽 Y在大鼠睪丸內的表達及其意義[J].中華男科學,2004,10(5):334-336,339. WANG X W,CUI Y G,CHEN Z Q,et al.Expression of Neuropeptide Y in rat testes and its significance[J].NationalJournalofAndrology,2004,10(5):334-336,339.(in Chinese)
[3] GONG Y G,FENG M M,HU X N,et al.Peptidergic not monoaminergic fibers profusely innervate the young adult human testis[J].JAnat,2009,214(3):330-338.
[4] WROBEL K H,BRANDL B.The autonomous innervation of the porcine testis in the period from birth to adulthood[J].AnnAnat,1998,180(2):145-156.
[5] WROBEL K H,MOUSTAFA M N.On the innervation of the donkey testis[J].AnnAnat,2000,182(1):13-22.
[6] SALEH A M,ALAMELDIN M A,ABDELMONIEM M E,et al.Immunohistochemical investigations of the autonomous nerve distribution in the testis of the camel (Camelusdromedarius) [J].AnnAnat,2002,184(3):209-220.
[7] MADDOCKS S,KERN S,SETCHELL B P.Investigating local regulation of the testes of ruminants[J].JReprodFertilSuppl,1995,49:309-319.
[8] DOSHI S B,KHULLAR K,SHARMA R K,et al.Role of reactive nitrogen species in male infertility [J/OL].ReprodBiolEndocrinol,2012,10:109.[2014-12-23].http://www.rbej.com/content/10//109.
[9] SUBURO A M,CHIOCCHIO S R,CANTSOLER M V,et al.Peptidergic innervation of blood vessels and interstitial cells in the testis of the cat[J].JAndrol,2002,23(1):121-134.
[10] SAINSBURY A,SCHWARZER C,COUZENS M,et al.Y4 receptor knockout rescues fertility in ob/ob mice[J].GenesDev,2002,16(9):1077-1088.
[11] 位 蘭,羅來強,彭克美,等.雛鴕鳥睪丸的結構及睪丸內神經肽Y的表達[J].中國獸醫學報,2009,29(12):1599-1602,1606. WEI L,LUO L Q,PENG K M,et al.Morphologic characteristics of testis and distribution of NPY in testis of the ostrich chicks[J].ChineseJournalofVeterinaryScience,2009,29(12):1599-1602,1606.(in Chinese)
[12] CHOW S H,GIGLIO W,ANESETTI R,et al.The effects of testicular denervation on spermatogenesis in the Sprague-Dawley rat[J].Neuroendocrinology,2000,72(1):37-45.
[13] 龔永光,楊宇如,張 煒,等.睪丸去神經支配對大鼠生殖細胞凋亡的影響[J].中華男科學,2006,12(11):968-973. GONG Y G,YANG Y R,ZHANG W,et al.Apoptosis in male germ cells induced by testicular denervation[J].NationalJournalofAndrology,2006,12(11):968-973.(in Chinese)
[14] MITAL P,HINTON B T,DUFOUR J M.The blood-testis and blood-epididymis barriers are more than just their tight junctions[J].BiolReprod,2011,84(5):851-858.
[15] LI N,WANG T,HAN D.Structural,cellular and molecular aspects of immune privilege in the testis [J/OL].FrontImmunol,2012,3:152.[2014-12-23].http://journal.frontiersin.org/Journal/10.3389/fimmu.2012.00152/full.
[16] KUNTZ A.Experimental degeneration in the testis of the dog[J].AnaRec,1919,17(4):220-234.
[17] VERHOEVEN G,HOEBEN E,DE GENDT K.Peritubular cell-Sertoli cell interactions:factors involved in PmodS activity[J].Andrologia,2000,32(1):42-45.
[18] PRINCE F P.Ultrastructural evidence of adrenergic,as well as cholinergic,nerve varicosities in relation to the lamina propria of the human seminiferous tubules during childhood[J].TissueCell,1996,28(5):507-513.
[19] MIYAKE K,YAMAMOTO M,MITSUYA H.Pharmacological and histological evidence for adrenergic innervation of the myoid cells in the rat seminiferous tubule[J].TohokuJExpMed,1986,149(1):79-87.
[20] MAYERHOFER A.Human testicular peritubular cells:more than meets the eye[J].Reproduction,2013,145(5):107-116.
[21] FRANKEL A I,RYAN E L.Testicular innervation is necessary for the response of plasma testosterone levels to acute stress[J].BiolReprod,1981,24(3):491-495.
[22] CHIOCCHIO S R,SUBURO A M,VLADUCIC E,et al.Differential effects of superior and inferior spermatic nerves on testosterone secretion and spermatic blood flow in cats[J].Endocrinology,1999,140(3):1036-1043.
(編輯 白永平)
The Distribution of PGP9.5 and NPY in the Normal Testis and Cryptorchidism of Bactrian Camel
YUAN Li-gang1*,YAN Zhen-long2,TAO Jin-zhong3,LIU Ying1*,CHEN Guo-juan1
(1.CollegeofVeterinaryMedicine,GansuAgriculturalUniversity,Lanzhou730070,China; 2.CollegeofLifeScienceandTechnology,GansuAgriculturalUniversity,Lanzhou730070,China; 3.CollegeofVeterinaryMedicine,NingxiaUniversity,Yinchuan750021,China)
The aim of the present study was to investigate the expression of PGP9.5 and NPY in the normal testis and cryptorchidism of Bactrian camel.The cryptorchidism and the normal testis of bactrian camel were studied by immunohistochemical SP technique and IPP (Image-Pro Plus) statistics method.The results showed that the PGP9.5 immunoreactive products distributed strongly in the Sertoli cells,sperm cells,arteries and veins of the normal testis,as well as NPY was moderate positive in Sertoli cell,sperm cells and arteries but weak positive in veins,in addition,Leydig cells in adult camels were strongly immunostained with NPY not only in normal testis but also in cryptorchidism.However,immunoreactivity of PGP9.5 and NPY were detected in cryptorchidism testis with significant differences intensities compared with the normal.These findings suggest that PGP9.5 and NPY are detected in the majority of cells in the seminiferous tubules and the blood vessels are involved in spermatogenesis and secretion of testosterone in Bactrian camel testis,they may play important roles in Bactrian camel male fertility and infertility.
Bactrian camel;cryptorchidism;PGP9.5;NPY;immunohistochemistry
10.11843/j.issn.0366-6964.2015.02.018
2014-06-16
國家自然科學研究基金(31160488);甘肅省自然科學基金項目(145RJZA223);甘肅省教育廳(0902B-03)
袁莉剛(1974-),女,甘肅隴南人,教授,主要從事動物發育生物學研究,Tel:0931-7631229
*通信作者:袁莉剛, E-mail:yuan2918@126.com;劉 英,E-mail:Liuy11@gsau.edu.cn
S852.162
A
0366-6964(2015)02-0303-06
