C2株藍(lán)氏賈第鞭毛蟲ILP基因的克隆表達(dá)及蛋白結(jié)構(gòu)分析
2015-03-17 01:46:57張亞粉王皓鈺郝金童高紅丹林志強(qiáng)劉阿倩雷田田王一同
中國人獸共患病學(xué)報 2015年7期
張亞粉,劉 巖,王皓鈺,郝金童,高紅丹,林志強(qiáng),劉阿倩,雷田田,王一同,余 源
C2株藍(lán)氏賈第鞭毛蟲ILP基因的克隆表達(dá)及蛋白結(jié)構(gòu)分析
張亞粉,劉 巖,王皓鈺,郝金童,高紅丹,林志強(qiáng),劉阿倩,雷田田,王一同,余 源
目的 利用大腸桿菌對C2株藍(lán)氏賈第鞭毛蟲ILP蛋白(Impact-like protein)進(jìn)行克隆表達(dá),運用生物信息學(xué)軟件進(jìn)行蛋白結(jié)構(gòu)分析。方法 提取C2株藍(lán)氏賈第鞭毛蟲基因組DNA,PCR擴(kuò)增ILP基因,構(gòu)建pGM-T重組載體,挑選陽性克隆并進(jìn)行序列分析;將ILP基因連入原核表達(dá)載體pET-28a(+),并轉(zhuǎn)化大腸桿菌Rosetta(DE3),IPTG誘導(dǎo)表達(dá)。運用PSIPRED和SWISS-MODEL進(jìn)行蛋白結(jié)構(gòu)分析。結(jié)果 成功構(gòu)建了原核表達(dá)載體pET-28a(+)-ILP,該基因全長831 bp。SDS-PAGE結(jié)果顯示,目的蛋白條帶出現(xiàn)在相對分子量約33 kD的位置,與預(yù)期相符。Western blot結(jié)果表明,大腸桿菌成功表達(dá)了重組蛋白。結(jié)論 成功克隆、表達(dá)并分析C2株藍(lán)氏賈第鞭毛蟲ILP蛋白,為藍(lán)氏賈第鞭毛蟲ILP蛋白結(jié)構(gòu)與功能的研究提供了有價值的資料。
C2株藍(lán)氏賈第鞭毛蟲;克隆;表達(dá);ILP蛋白
藍(lán)氏賈第鞭毛蟲(Giardialamblia)簡稱賈第蟲,寄生于人和某些哺乳動物的小腸,引起賈第蟲病。賈第蟲具有2個細(xì)胞核和4對鞭毛,生活于厭氧或微需氧的環(huán)境中。本蟲是生物進(jìn)化過程中最早分化出來的真核生物之一[1-2],被稱為“生物活化石”。自上世紀(jì)80年代起,國內(nèi)學(xué)者已開始對本蟲進(jìn)行生物學(xué)實驗研究[1-4]。賈第蟲是真核生物進(jìn)化史的早期物種,其基因組已被測序[5-7],但很多基因的功能尚不明確。藍(lán)氏賈第鞭毛蟲作為原始的真核生物,與高等真核生物相比,其基因的生物學(xué)結(jié)構(gòu)與功能既有相似性又有獨特性。因此,對賈第蟲與高等真核生物相似基因的研究,有助于進(jìn)一步了解其與高等真核生物的區(qū)別及其在生物進(jìn)化中的地位。
真核生物ILP蛋白(protein impact)是一種重要的翻譯調(diào)控因子,在氨基酸含量較低的情況下,能夠保證基因高水平表達(dá)[8-10]。GenBank基因檢索表明Morrison等在WB株藍(lán)氏賈第鞭毛蟲發(fā)現(xiàn)其可能具有ILP蛋白基因序列。本研究以國內(nèi)C2株藍(lán)氏賈第鞭毛蟲為材料的基因克隆和表達(dá)結(jié)果表明,C2株藍(lán)氏賈第鞭毛蟲具有ILP蛋白基因序列。進(jìn)而通過對該基因測序結(jié)果進(jìn)行生物信息學(xué)分析,構(gòu)建了ILP蛋白的高級空間結(jié)構(gòu),為進(jìn)一步研究藍(lán)氏賈第鞭毛蟲ILP蛋白結(jié)構(gòu)與功能的關(guān)系,以及賈第蟲與高等真核生物基因功能的相似性提供了有價值的資料。
1 材料與方法
1.1 材料 C2株賈第蟲由首都醫(yī)科大學(xué)寄生蟲學(xué)教研室提供,原核表達(dá)質(zhì)粒pET-28a(+)由本實驗室保存;Rosetta(DE3) 菌株由中國醫(yī)學(xué)科學(xué)院北京協(xié)和醫(yī)學(xué)院血液學(xué)研究所王建祥教授惠贈;組織/細(xì)胞基因組DNA提取試劑盒購自北京艾德萊生物科技有限公司;T4 DNA 連接酶、pGM-T連接試劑盒購自TAKARA公司;限制性內(nèi)切酶NcoⅠ、XhoⅠ購自NEB公司;Taq DNA聚合酶、E.coliTop 10感受態(tài)細(xì)胞、Pfu DNA Polymerase、DNA Marker III、質(zhì)粒小提試劑盒、瓊脂糖凝膠回收試劑盒購自天根公司;Page Ruler購自Fermentas公司;His-tag標(biāo)簽抗體、PVDF膜購自康為世紀(jì)公司;引物由生工生物工程(上海)公司合成。
1.2 方法
1.2.1 蟲體的培養(yǎng) 復(fù)蘇用液氮保存的藍(lán)氏賈第鞭毛蟲C2株,用改良TYI-S-33培養(yǎng)基于37 ℃培養(yǎng)。48~72 h后,蟲體即呈對數(shù)生長期。選取蟲體生長旺盛的培養(yǎng)管,置4 ℃冰浴,15 min后取出,在雙手掌間多次滾搓培養(yǎng)管,用血球計數(shù)板計數(shù)蟲數(shù),再用培養(yǎng)基將蟲液濃度調(diào)為6×106~10×106個滋養(yǎng)體/mL[11]。
1.2.2 藍(lán)氏賈第鞭毛蟲基因組DNA提取 采用組織/細(xì)胞基因組DNA提取試劑盒提取藍(lán)氏賈第鞭毛蟲的基因組DNA,實驗操作按說明書進(jìn)行。通過紫外分光光度法鑒定基因組DNA的純度。
1.2.3 目的基因的PCR擴(kuò)增 參考GenBank報道的藍(lán)氏賈第鞭毛蟲ILP(XM 001705285)基因序列,采用Primer Premier 5軟件設(shè)計特異性引物。引物中加入特異性酶切位點(下劃線標(biāo)示酶切位點NcoⅠ、XhoⅠ)
ILP基因上游引物序列(P1):CATGCCATGGGCATGAATGAGCAGCTAGGCGC;
ILP基因下游引物序列(P2):CCGCTCGAGCAGCCCACTACAAAAGTTTGG;
以賈第蟲基因組DNA作為模板,P1、P2為引物,擴(kuò)增目的序列。反應(yīng)體系為50 μL:10×pfu PCR緩沖液5 μL、10 mmol/L dNTP 4 μL、模板DNA 1 μL、Pfu Taq酶0.5 μL、10 nmol/L上下游引物各2 μL。反應(yīng)條件為94 ℃預(yù)變性5 min;94 ℃變性30 s,56 ℃退火30 s,72 ℃延伸1 min,共30個循環(huán);最后72 ℃延伸5 min。加入Taq酶0.5 μL,72 ℃延伸15 min,使擴(kuò)增基因片段末端加入A尾。
1.2.4 pGM-T-ILP克隆載體的構(gòu)建與序列分析 PCR產(chǎn)物經(jīng)1.0%瓊脂糖凝膠電泳,利用膠回收試劑盒進(jìn)行回收,經(jīng)T4 DNA連接酶催化連接pGM-T載體,轉(zhuǎn)化E.coliTop10感受態(tài)細(xì)胞,并將轉(zhuǎn)化菌涂于含有氨芐青霉素和X-gal及IPTG的LB平板上,37 ℃培養(yǎng)12 h。挑取陽性單菌落接種于5 mL LB 培養(yǎng)基(含100 mg/L氨芐青霉素) 的試管中, 37 ℃振蕩過夜。質(zhì)粒小提試劑盒提取細(xì)菌質(zhì)粒,ILP基因重組載體經(jīng)NcoⅠ、XhoⅠ雙酶切,1%瓊脂糖凝膠電泳鑒定陽性克隆,陽性克隆菌送上海生工公司測序。
1.2.5 pET-28a(+)-ILP表達(dá)載體的構(gòu)建 將pET-28a(+)載體和pGM- T-ILP分別用NcoⅠ、XhoⅠ雙酶切,利用膠回收試劑盒進(jìn)行回收,回收產(chǎn)物經(jīng)瓊脂糖凝膠電泳估濃度后經(jīng)T4連接酶催化連接,轉(zhuǎn)化E.coliTop10感受態(tài)細(xì)胞。通過Kana霉素和氯霉素雙抗性平板篩選陽性克隆,利用NcoⅠ、XhoⅠ雙酶切進(jìn)行鑒定。
1.2.6 ILP誘導(dǎo)表達(dá) 經(jīng)雙酶切鑒定正確的質(zhì)粒轉(zhuǎn)化Rosetta(DE3),經(jīng)卡那霉素和氯霉素雙抗平板篩選陽性單菌落并擴(kuò)大培養(yǎng),加入不同終濃度(0.5~2.0 mmol/L)的異丙基-β-D-硫代半乳糖苷(IPTG)誘導(dǎo)表達(dá)2~4 h。離心收集菌體、超聲裂解后離心收集上清,取40 μL進(jìn)行SDS-PAGE,檢測目的蛋白表達(dá)。
1.2.7 Western blot鑒定重組蛋白表達(dá) 離心收集菌體、超聲裂解后離心,取上清40 μL進(jìn)行SDS-PAGE,300 mA轉(zhuǎn)印PVDF膜0.5 h,抗His-Tag鼠源多克隆抗體作為一抗,兔抗鼠IgG抗體作為二抗進(jìn)行Western blot鑒定。
1.2.8 ILP蛋白生物信息學(xué)分析 利用SWISS-MODEL (http://swissmodel.expasy.org/)和PSIPRED蛋白序列分析服務(wù)器對C2藍(lán)氏賈第鞭毛蟲ILP(XM 001705285)進(jìn)行序列分析,構(gòu)建ILP蛋白模型,預(yù)測其高級空間結(jié)構(gòu)。
2 結(jié) 果
2.1 pGM-T-ILP克隆載體的構(gòu)建 PCR擴(kuò)增藍(lán)氏賈第鞭毛蟲ILP基因,1.0%瓊脂糖凝膠電泳結(jié)果顯示,在831 bp附近有明顯條帶,與預(yù)期大小相符(圖1)。ILP基因重組載體經(jīng)NcoⅠ、XhoⅠ雙酶切,1%瓊脂糖凝膠電泳,可見831 bp的ILP目的基因片段及3 037 bp的pGM-T載體電泳條帶(圖2)。
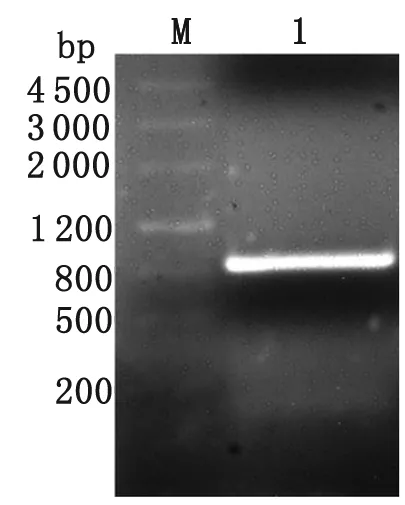
M: DNA marker; 1: ILP PCR product.
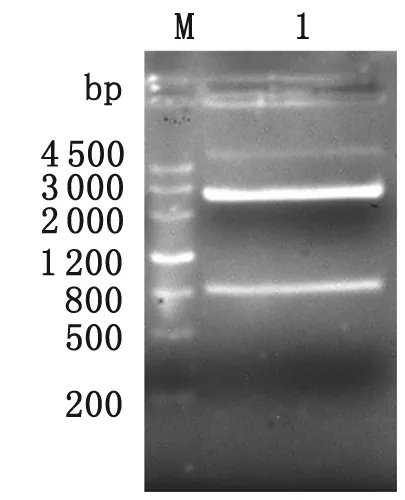
M: DNA marker; 1: pGM-T-ILP digested byNcoⅠandXhoⅠ.
圖2 pGM-T-ILP重組載體NcoⅠandXhoⅠ雙酶切鑒定
Fig.2 Restriction enzymes analysis of recombinant plasmid pGM-T-ILP
2.2 pET28a(+)-ILP表達(dá)載體的構(gòu)建 經(jīng)NcoⅠ、XhoⅠ雙酶切pET28a(+)-ILP重組載體,電泳可見831 bp的ILP基因目的片段及5 369 bp的載體pET28a(+)兩條電泳條帶(圖3)。
2.3 ILP蛋白的誘導(dǎo)表達(dá)鑒定 將構(gòu)建好的重組質(zhì)粒轉(zhuǎn)化到Rosetta(DE3)中表達(dá),菌液分別加入不同終濃度IPTG(0.5、1.0、1.5、2.0 mmol/L),未加IPTG誘導(dǎo)的轉(zhuǎn)化菌作為陰性對照,SDS-PAGE電泳結(jié)果顯示,在33 kD處出現(xiàn)目的條帶,與預(yù)期結(jié)果一致。IPTG終濃度為1.0 mmol/L誘導(dǎo)4 h,ILP表達(dá)量最高(圖4)。Western blot鑒定結(jié)果表明,在33 kD處出現(xiàn)重組蛋白(圖5)。
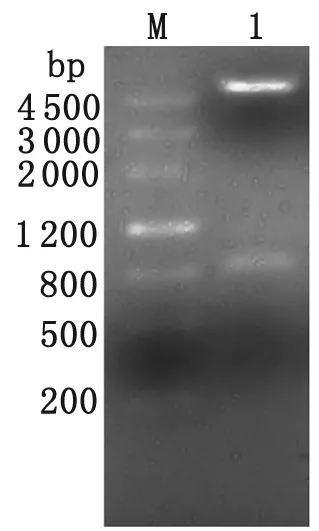
M: DNA marker; 1: pET28a(+)-ILP digested byNcoⅠandXhoⅠ
圖3 pET28a(+)-ILP重組載體NcoⅠandXhoⅠ雙酶切鑒定
Fig.3 Restriction enzymes analysis of recombinant plasmid pET28a(+)-ILP

M: PAGE ruler; 1-4: induced with 0.5-2.0 mmol/L IPTG for 2 h; 5-8: induced for 4 h; 9: without IPTG.
圖4 不同時間、不同濃度IPTG誘導(dǎo)ILP蛋白表達(dá)SDS-PAGE電泳
Fig.4 SDS-PAGE analysis of the ILP protein expressed in Rosetta (DE3)
2.4 ILP蛋白生物信息學(xué)分析數(shù)據(jù) PSIPRED蛋白序列分析結(jié)果顯示,ILP蛋白全長序列主要包含6個α螺旋和7個β折疊,其余部分以無規(guī)則卷曲為主(圖6)。
將ILP蛋白氨基酸序列提交至SWISS-MODEL建模服務(wù)系統(tǒng),選擇NMR作為建模方法,結(jié)果顯示ILP蛋白與GCN2 eIF2 alpha kinase N端結(jié)構(gòu)域同源性為17.7%,能夠預(yù)測出ILP蛋白部分空間結(jié)構(gòu),包含3個β折疊和3個α螺旋,推測此結(jié)構(gòu)可能與抑制GCN2蛋白激酶的活性相關(guān)(圖7)。
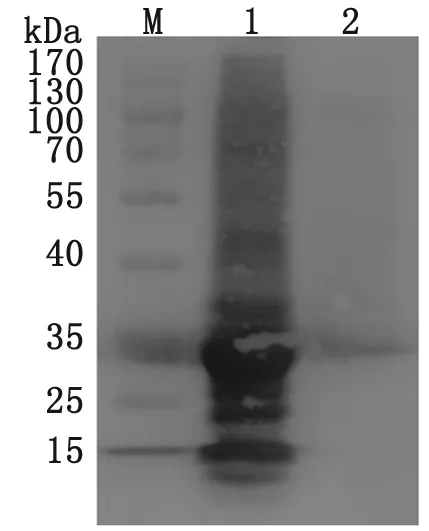
M: PAGE ruler; 1: induced with 1.0 mmol/L IPTG for 4 h.
圖5 ILP蛋白Western blot分析
Fig.5 Western blot analysis of the ILP protein

圖6 PSIPRED分析ILP蛋白的二級結(jié)構(gòu)
Fig.6 Secondary structure ofGiardialambliaILP protein with PSIPRED
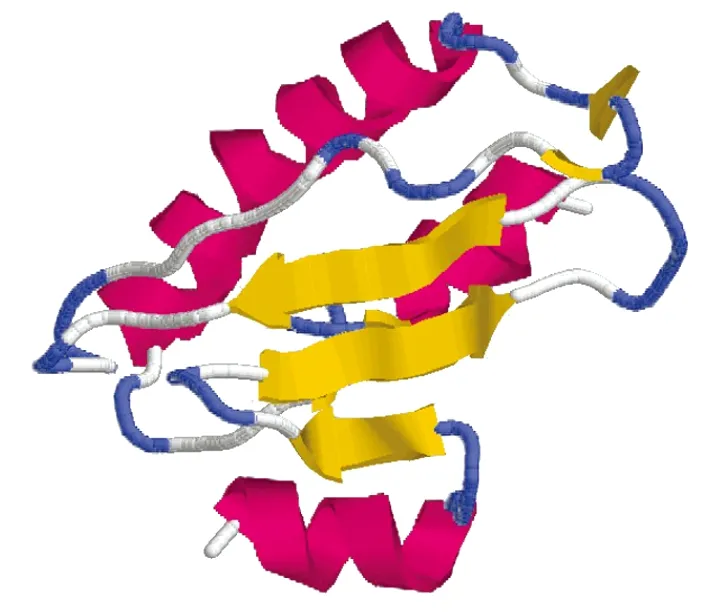
圖7 SWISS-MODEL模擬賈第蟲ILP蛋白三維模型
Fig.7GiardialambliaILP structure predicted by SWISS-MODEL
3 討 論
本研究參考GenBank報道的WB株藍(lán)氏賈第鞭毛蟲ILP(XM 001705285)基因序列設(shè)計引物,利用原核表達(dá)技術(shù)對C2株藍(lán)氏賈第鞭毛蟲ILP蛋白進(jìn)行克隆表達(dá),結(jié)果表明C2株藍(lán)氏賈第鞭毛蟲具有ILP基因序列。本研究中使用的大腸桿菌Rosetta(DE3)是BL21的改良菌種,能夠補(bǔ)充BL21缺乏的6種稀有密碼子所對應(yīng)的tRNA,提高重組質(zhì)粒的表達(dá)效果[12]。本研究中使用的pET28a(+)載體是原核表達(dá)中最常用的載體之一,克隆到pET28a(+)載體上的目的基因受噬菌體T7強(qiáng)轉(zhuǎn)錄及翻譯信號控制,不被宿主細(xì)胞RNA聚合酶識別。無IPTG誘導(dǎo)劑時,克隆到pET28a(+)載體的基因?qū)嶋H上是被關(guān)閉的,因此不會由于表達(dá)的蛋白有毒性而引起質(zhì)粒不穩(wěn)定。加入IPTG誘導(dǎo)劑能夠誘導(dǎo)表達(dá)T7 RNA聚合酶,啟動重組蛋白表達(dá),增加實驗結(jié)果的可靠性[13]。此外,pET28a(+)載體自帶His標(biāo)簽,Western blot檢測重組蛋白表達(dá)時能夠利用His標(biāo)簽作為重組蛋白表達(dá)的信號。
為了進(jìn)一步了解C2株藍(lán)氏賈第鞭毛蟲ILP蛋白的結(jié)構(gòu)與功能,本研究利用SWISS-MODEL和PSIPRED蛋白序列分析服務(wù)器對C2藍(lán)氏賈第鞭毛蟲ILP蛋白進(jìn)行分析,構(gòu)建ILP蛋白模型。SWISS-MODEL是日內(nèi)瓦生物醫(yī)學(xué)研究所(Geneva Biomedical Research Institute)建立發(fā)展起來的分子建模服務(wù)系統(tǒng)(Molecular Modeling Server)。基于同源建模的原理,SWISS-MODEL從布魯克海文蛋白質(zhì)結(jié)構(gòu)數(shù)據(jù)庫(Brookhaven PDB)中提取蛋白質(zhì)查詢序列的模擬結(jié)構(gòu)信息,用已知結(jié)構(gòu)蛋白建立未知結(jié)構(gòu)蛋白的分子模型[14-15]。由于SWISS-MODEL是基于已知生物大分子結(jié)構(gòu)基礎(chǔ)上的分子建模系統(tǒng),需要龐大的分子建模基礎(chǔ)數(shù)據(jù)庫才能建立正確的結(jié)構(gòu)模型,有限的數(shù)據(jù)資源使得該方法具有一定的局限性。為了彌補(bǔ)SWISS-MODEL服務(wù)器的不足,獲得ILP蛋白更多的結(jié)構(gòu)信息,本研究同時采用能夠根據(jù)氨基酸排列順序預(yù)測蛋白全長二級結(jié)構(gòu)的PSIPRED服務(wù)器分析ILP蛋白二級結(jié)構(gòu),結(jié)果表明ILP蛋白全長序列主要包含6個α螺旋和7個β折疊,其余部分以無規(guī)則卷曲為主,與GCN2 eIF2 alpha kinase N端結(jié)構(gòu)域區(qū)域具有一定的同源性,同源區(qū)域包含3個β折疊和3個α螺旋。
本研究通過SWISS-MODEL和PSIPRED蛋白序列分析服務(wù)器成功預(yù)測出ILP蛋白與GCN2 eIF2 alpha kinase N端結(jié)構(gòu)域具有一定的同源性,推測其功能可能與抑制GCN2蛋白激酶的活性相關(guān)。有研究表明ILP蛋白作為翻譯調(diào)控因子,能夠與GCN1/GCN1L1相互作用,抑制GCN2蛋白激酶的活性,阻止ATF4的合成,在氨基酸含量較低的情況下保證基因高水平表達(dá)[16],但是ILP蛋白發(fā)揮其調(diào)控功能的具體機(jī)制尚不明確,此外,抑制ILP蛋白表達(dá)可能導(dǎo)致藍(lán)氏賈第鞭毛蟲基因表達(dá)受阻,降低藍(lán)氏賈第鞭毛蟲的生長繁殖速度,對藍(lán)氏賈第鞭毛蟲的防治可能會起到一定的效果,今后的工作應(yīng)圍繞這些問題進(jìn)一步研究。總之,本研究為進(jìn)一步明確藍(lán)氏賈第鞭毛蟲ILP蛋白的結(jié)構(gòu)與功能,賈第蟲與高等真核生物基因功能的相似性,以及藍(lán)氏賈第鞭毛蟲的防治提供了新思路。
[1]Lu SQ. Domestic research ofGiardialamblia[J]. Acta Parasitologica Et Medica Entomologica Sinica, 1999, 6(4): 193-200. (in Chinese) 盧思奇.國內(nèi)藍(lán)氏賈第鞭毛蟲研究[J].寄生蟲與醫(yī)學(xué)昆蟲報,1999,6(4):193-200.
[2]Tian XF, Lu SQ. Cytoskeleton ofGiardialamblia[J]. World Chin J Digestol, 2005, 13(12): 1434-1436. (in Chinese) 田喜鳳,盧思奇.藍(lán)氏賈第鞭毛蟲的細(xì)胞骨架[J].世界華人消化雜志, 2005, 13(12): 1434-1436.
[3]Adam RD. Biology ofGiardialamblia[J]. Clin Microbiol Rev, 2001, 14(3): 447-475.
[4]Sandhu H, Mahajan RC, Ganguly NK. Flow cytometric assessment of the effect of drugs onGiardialambliatrophozoites in vitro[J]. Mol Cell Biochem, 2004, 265: 151-160.
[5]Gardner TB, Hill DR. Treatment of giardiasis[J]. Clin Microbiol Rev, 2001, 14(1): 114-128. DOI: 10.1128/CMR.14.1.114-128.2001
[6]Ali SA, Hill DR. Giardia intestinalis[J]. Curr Opin Infect Dis, 2003, 16(5): 453-460.
[7]Triana O, Galanti N, Olea N, et al. Chromatin and histones fromGiardialamblia: A new puzzle in primitive eukaryotes[J]. J Cellular Biochem, 2001, 82(4): 573-582. DOI: 10.1002/jcb.1159/full
[8]Okamura K, Hagiwara-Takeuchi Y, Li T, et al. Comparative genome analysis of the mouse imprinted gene impact and its nonimprinted human homolog IMPACT: toward the structural basis for species-specific imprinting[J]. Genome Res, 2000, 10(12): 1878-1889. DOI: 10.1101/gr.139200
[9]Gerhard DS, Wagner L, Feingold EA. The status, quality, and expansion of the NIH full-length cDNA project: the Mammalian Gene Collection (MGC)[J]. Genome Res,2004, 14 (10B): 2121-2127. DOI: 10.1101/gr.2596504
[10]Ota T, Suzuki Y, Nishikawa T, et al. Complete sequencing and characterization of 21,243 full-length human cDNAs[J]. Nat Genet, 2004, 36(1): 40-45. DOI: 10.1038/ng1285
[11]He B, Liu GW, Cao L, et al.Invitroidentification of the cytoskeleton proteins ofGiardialambliausing a MS technique[J]. J Pathog Biol, 2010, 5(12): 898-900. (in Chinese) 何冰,劉廣偉,曹蕾,等.質(zhì)譜技術(shù)鑒定體外藍(lán)氏賈第鞭毛蟲的細(xì)胞骨架蛋白[J].中國病原生物學(xué)雜志,2010,5(12):898-900.
[12]Wang YH, Zheng GX, Ju HM, et al. Cloning and expression of H2A gene fromGiardialamblia[J]. J Trop Med, 2011, 11(10): 1107-1110. (in Chinese) 王云華,鄭國俠,巨紅妹,等.藍(lán)氏賈第鞭毛蟲中國株組蛋白H2A基因的克隆與重組表達(dá)[J].熱帶醫(yī)學(xué)雜志,2011,11(10):1107-1110.
[13]Wang Y, Yu Y, Wang WL, et al. Prokaryotic expression and purification of α-4 giardin ofGiardialamblia[J]. Chin J Zoonoses, 2011, 27(6): 465-469.(in Chinese) 王洋,余源,王衛(wèi)亮,等. 藍(lán)氏賈第鞭毛蟲α-4 賈第素的原核表達(dá)及純[J].中國人獸共患病學(xué)報,2011,27(6): 465-469.)
[14]Xia XY, Wu QY, An LM et al. A novel P20R mutation in the alpha-B crystallin gene causes autosomal dominant congenital posterior polar cataracts in a Chinese family[J]. BMC Ophthalmol, 2014, 14(1): 108-111. DOI: 10.1186/1471-2415-14-108
[15]Kumar A, Pillay B, Olaniran AO. Two structurally different dienelactone hydrolases (TfdEI and TfdEII) from Cupriavidus necator JMP134 plasmid pJP4 catalyse cis- and trans-dienelactones with similar efficiency[J]. PLoS One, 2014, 9(7): 104-109.
[16]Habibi D, Jalili RB, Forouzandeh F, et al. High expression of IMPACT protein promotes resistance to indoleamine 2, 3-dioxygenase-induced cell death[J]. J Cell Physiol, 2010, 225(1): 196-205. DOI: 10.1002/jcp.22220
Cloning, expression and protein structural analysis of ILP gene from C2Giardialamblia
ZHANG Ya-fen,LIU Yan,WANG Hao-yu,HAO Jin-tong,GAO Hong-dan,LIN Zhi-qiang,LIU A-qian,LEI Tian-tian,WANG Yi-tong,YU Yuan
(CollegeofLifeScience,NorthChinaUniversityofScienceandTechnology,Tangshan063000,China)
The aim of this study is to clone, express and analyze ILP (Impact-like protein) gene from C2Giardialamblia. Genome DNA of C2Giardialambliawas extracted with DNA extraction kit. Specific primers were designed by primer design software Primer Premier 5. ILP gene was amplified by PCR and cloned into pGM-T vector, and transformed intoE.coli(Top10) competent bacteria cells. Positive clone was chosen and the sequence was analyzed. The pGM-T-ILP recombinant vector was digested by restriction endonucleaseNcoⅠandXhoⅠ. Target gene was restructured into pET28a(+) vector at theNcoⅠandXhoⅠrestriction sites to generate recombinant expression vector pET28a(+)-ILP. The vector was transformed intoE.coliRosetta (DE3) and induced with IPTG (isopropyl β-D-thiogalactopyranoside). SDS-PAGE result showed the objective protein band in the position of the relative molecular weight was 33 kDa. Western blot analysis with His-Tag antibody indicated that the ILP gene was successfully expressed in the prokaryotic Rosetta (DE3). Our results provided valuable experimental data for further understanding the structure and function of ILP gene.
C2Giardialamblia; clone; expression; ILP gene
Yu Yuan, Email: yuyuan5188@163.com
國家自然科學(xué)基金(No. 31471954),唐山市科技支持計劃項目(No.12140209A-33),河北青年科學(xué)基金(No. C2012401039)聯(lián)合資助,河北聯(lián)合大學(xué)培養(yǎng)基金(No. GP201308)
余源,Email:yuyuan5188@163.com
華北理工大學(xué)生命科學(xué)學(xué)院,唐山 063000
10.3969/cjz.j.issn.1002-2694.2015.07.005
R382.2
A
1002-2694(2015)07-0618-05
2014-09-30;
2015-05-05

Supported by the National Natural Science Foundation of China (No. 31471954), the Scientific and Technological Support Projects from Tangshan (No. 12140209A-33), the Youth Science Fund Project in Hebei Province (No. C2012401039), and the Hebei United University Training Fund (No. GP201308)
