干旱荒漠白刺灌叢植被演替過程土壤種子庫變化特征
2015-03-15 06:41:55馬全林魏林源靳虎甲
生態學報 2015年7期
關鍵詞:物種
馬全林,盧 琦,魏林源,靳虎甲
1 中國林業科學研究院荒漠化研究所, 北京 100091 2 甘肅省治沙研究所荒漠化與風沙災害防治國家重點實驗室培育基地, 蘭州 730070
干旱荒漠白刺灌叢植被演替過程土壤種子庫變化特征
馬全林1,2,*,盧 琦1,魏林源2,靳虎甲2
1 中國林業科學研究院荒漠化研究所, 北京 100091 2 甘肅省治沙研究所荒漠化與風沙災害防治國家重點實驗室培育基地, 蘭州 730070
土壤種子庫是反映氣候與土地利用變化的指示器,也是荒漠植被更新與恢復的基礎。典型荒漠白刺灌叢植被廣泛分布于我國西北綠洲邊緣地帶,對保護綠洲免受風沙危害發揮著重要作用。但是,對干旱荒漠白刺灌叢植被土壤種子庫仍未見研究報道。應用空間代替時間的方法,選擇石羊河下游白刺灌叢植被演替的初始階段、穩定階段、衰退階段與嚴重衰退階段樣地,研究了白刺灌叢植被演替過程的土壤種子庫物種組成、時空分布和數量變化特征。結果表明:石羊河下游白刺灌叢植被演替過程中,土壤種子庫共出現9科18種植物,其中草本植物占到55%—80%,是白刺灌叢植被土壤種子庫的主體。不同演替階段土壤種子庫密度以穩定階段最高,達到660.7 粒/m2,是初始階段、衰退階段和嚴重衰退階段的5.6、14.5倍和6.2倍。不同沙堆部位,土壤種子庫密度以迎風坡中部與背風坡中部最高,分別達到329 粒/m2和309 粒/m2;沙堆土壤種子庫密度是堆間地的1.9倍,土壤種子庫主要分布于灌叢沙堆上。不同演替階段土壤種子庫間的相似性系數均在0.6以下,除穩定階段外,其他階段間均屬于中等相似水平;土壤種子庫與地上植被的相似性系數均在0.5以上,達到中等相似或極相似水平。該研究進一步證明了灌叢沙堆和土壤種子庫在白刺灌叢植被中的重要性,也說明人為干擾損害了白刺灌叢地下潛在植被及其恢復潛力。因此,需要開展人工種子庫或人工植被建設以控制干旱荒漠白刺灌叢沙堆活化,并實現退化白刺灌叢植被恢復。
石羊河下游; 白刺灌叢植被; 灌叢沙堆; 土壤種子庫; 相似性
土壤種子庫是反映氣候與土地利用變化的指示器,預示著將來與環境變化有關的植被變化,在連接過去、現在和將來的植物種群、群落結構與動態中起著重要的生態作用,也是荒漠植被更新與恢復的基礎[1- 2],因此研究土壤種子庫特征可為揭示植被退化/恢復演替機理提供重要依據。白刺灌叢(Nitrariaspp.)是我國干旱荒漠植被的重要建群種之一,廣泛分布于西北沙漠地區,其抗旱耐瘠薄,特別喜沙埋,埋后枝節生出不定根與新植株,攔阻并積聚風沙逐漸形成丘狀沙堆,在保障干旱荒漠區生態安全中發揮著極其重要的作用[3- 6]。石羊河下游民勤縣天然白刺灌叢植被面積達到15.2 萬hm2,占該區域天然灌木林的31.6%,是阻止騰格里和巴丹吉林兩大沙漠合攏,保障民勤綠洲生態安全的首要生態屏障。但是,受農業大規模用水引起的地表水大幅度減少與地下水位降低的影響,白刺灌叢植被出現了不同程度退化,并相繼發生了沙堆的風蝕、流動,嚴重影響和削弱了白刺灌叢植被的生態屏障作用[6- 9]。關于白刺灌叢植被的退化與恢復已成為干旱區生態研究的熱點之一,已開展了白刺灌叢植被退化原因、恢復技術以及土壤呼吸、風蝕等退化特征研究,但對作為潛在植被的地下土壤種子庫物種組成、演變規律以及影響因素等尚未見報道[6,9- 10]。
本研究采用空間代替時間的方法,在石羊河下游選擇不同演替階段的白刺灌叢植被,研究了其土壤種子庫的物種組成、數量特征及其與地上植被的關系,以揭示干旱荒漠白刺灌叢演植被演替過程中土壤種子庫的變化規律,探討白刺灌叢植被的自我修復能力,為退化白刺灌叢植被的恢復提供理論與技術支撐。
1 研究區概況
研究區選擇地處干旱荒漠區的甘肅省民勤縣(38°05′—39°06′ N,103°02′—104°02′ E),其屬于我國典型的綠洲型荒漠地區,東、西、北三面被騰格里沙漠與巴丹吉林沙漠包圍,氣候屬溫帶荒漠氣候,多年平均氣溫7.6 ℃,降水量113.2 mm,蒸發量2604.3 mm;全年盛行西北、西北偏西風,平均風速2.5 m/s,大風日數26.3 d,沙塵暴日數25.0 d,揚沙日數37.5 d,浮塵日數29.7 d(民勤治沙綜合試驗站1961—2011年氣象統計資料)。土壤多為風沙土,養分貧瘠且風蝕嚴重。地表水資源由20世紀50年代的5.731 億m3減少到本世紀初期的1.0 億m3左右。地下水位由1961年的2.2 m下降到2001年的18.4 m,每年以0.50—0.71 m的速度下降。2008年實施石羊河流域綜合治理工程后,通過增加石羊河下泄流量和關井壓田等措施的實施,緩解了地下水位下降。固沙植被中,喬木胡楊(Populuseuphratica)、沙棗(Elaeagnusangustifolia)已全部枯死,灌木梭梭(Haloxylonammodendron)、檸條(Caraganakorshinskii)、紅柳(Tamarixspp.)和白刺(Nitrariaspp.)出現嚴重衰敗[8]。水資源的減少,植被的退化使研究區成為全國浮塵、揚沙和沙塵暴發生最嚴重地區之一,也成為我國沙塵暴的主要策源地之一。
2 研究方法
2.1 樣地設置
以唐古特白刺(NitrariaTangutorum)荒漠灌叢植被為研究對象,根據白刺灌叢植被生長及灌叢沙堆形態和結皮狀況,采用空間代替時間序列的方法,選擇石羊河下游民勤縣紅土井、青土湖、三角城、沙井子白刺灌叢沙堆樣地分別代表白刺灌叢植被演替的初始階段、穩定階段、衰退階段和嚴重衰退階段[10]。其中,三角城衰退階段和沙井子嚴重衰退階段樣地位于綠洲邊緣,受綠洲地下水過度利用的嚴重人為干擾影響。
2.2 土壤種子庫取樣、萌發與鑒定
2010年7月(荒漠植物開花、結實期),在白刺灌叢植被不同演替階段樣地,選擇平均大小的沙堆5個,每個沙堆沿西北—東南走向(研究區主風向),在沙堆迎風坡腳、迎風面中部、頂部、背風面中部、背風坡腳和堆間地使用專制取樣框采集土壤樣品,采樣框大小25 cm×25 cm。由于沙漠地區種子庫主要分布于表層土壤,因此采樣深度僅設置0—5 cm[ 2]。
為縮短萌發培養時間,提高種子萌發率,土壤種子庫樣品用0.2 mm的小網土壤篩進行濃縮處理[2]。濃縮樣品充分混勻后平鋪于高25 cm,半徑20 cm的圓形花盆(底部裝有120 ℃高溫處理的沙丘沙),厚度為1—2.5 cm,上蓋保護性紗網防止外來種子污染,在簡易溫室中進行萌發培養[2]。用灑水壺適時澆水保持土壤濕潤,逐日觀察記錄種子萌發情況,對萌發的不同幼苗進行標記,待能鑒別出植物種后除去,萌發培養持續1月時間。
2.3 白刺灌叢植被及沙堆調查
在不同演替階段樣地,間隔200 m設置30 m×30 m大樣方3個,調查白刺灌叢植被組成與數量特征,測定樣方內每個白刺灌叢沙堆的長軸、短軸、高度以及白刺灌叢高度、枝條長度和生物量等。同時,在每個大樣方內設置1 m×1 m的小樣方5個,調查草本植物的組成與數量,并統計地上植被種類組成及其密度、蓋度等。
不同演替階段樣地植被蓋度以及白刺灌叢沙堆長軸、短軸、高度及其覆蓋度、生物量總體差異顯著。其中,白刺灌叢沙堆長軸、短軸、高度均以衰退階段最大,初始階段最小,沙堆大小總體隨白刺灌叢植被演替過程逐漸增大,但嚴重衰敗階段沙堆因風蝕活化而變小。沙堆灌叢覆蓋度、單位面積生物量以及群落蓋度以穩定階段最大,嚴重衰退階段最小,隨白刺灌叢植被演替過程呈先增大后降低趨勢(表1)。
2.4 白刺灌叢植被區土壤調查
在不同演替階段樣地,調查沙堆結皮覆蓋面積比例,挖取3個0—100 cm 土壤剖面,每20 cm為一層測定土壤容重、水分與粒度組成。在沙堆迎風坡腳、迎風面中部、頂部、背風面中部、背風坡腳和堆間地,測定0—5 cm表層土壤的容重、水分與粒度特征。
白刺灌叢植被不同演替階段0—5 cm土壤物理性狀與沙堆結皮差異明顯。其中,沙堆結皮經歷形成、發展與破損的演變過程,結皮厚度、覆蓋度和生物結皮比例呈現先增加后減少的趨勢。表層土壤容重、沙粒成分隨演替過程呈先減小后增大趨勢;土壤水分、粘粒與粉粒成分呈先增大后減小趨勢(表 2)。

表1 不同演替階段白刺灌叢植被與沙堆基本特征Table 1 Main characteristics of Nitraria vegetation and nebkhas at the different succession stages
不同字母間表示差異顯著

表2 白刺灌叢植被不同演替階段沙堆表層土壤特征Table 2 Topsoil characteristics of Nitraria nebkhas at the different succession stages
白刺灌叢沙堆不同部位0—5 cm土壤粒度組成均以沙粒成分為主,且各部位間沙粒含量差異不顯著。土壤容重以堆頂部最大,除堆間地外,與其他沙堆部位無顯著差異。土壤水分、粘粒含量和粉粒含量以堆間地最大,與沙堆各部位差異顯著,但坡腳、迎風坡中部、堆頂部與背風坡間無顯著差異(表 3)。

表3 白刺灌叢沙堆不同部位表層土壤特征Table 3 Topsoil characteristics at the different positions of Nitraria nebkhas
2.5 數據分析
應用Excel和SPSS16.0軟件完成數據處理和統計分析,采用LSD最小顯著差數法對白刺灌叢植被、土壤與沙堆特征及其土壤種子密度進行顯著性分析(取顯著度0.05)。
根據土壤種子庫萌發計數計算白刺灌叢植被土壤種子庫密度、多樣性指數以及相似性系數。土壤種子庫密度用單位面積土壤內所含有的活力種子數量來表示(粒/m2)。土壤種子庫物種多樣性采用最常用的Simpson和Shannon-Wiener多樣性指數以及Pielou均勻度指數測度[11]。不同演替階段土壤種子庫、土壤種子庫與地上植被相似性采用Jaccard相似性系數測度,當Jaccard相似性系數達到0.00—0.25時為極不相似,0.25—0.50時為中等不相似,0.50—0.75時為中等相似,0.75—1.00時為極相似[12]。
3 結果分析
3.1 土壤種子庫的物種組成變化
石羊河下游白刺灌叢植被土壤種子庫共記錄到9科18種植物,其中初始階段、穩定階段、衰退階段與嚴重衰退階段分別記錄到9科14種、6科9種、7科13種和8科10種植物。種子庫物種組成中,藜科6種,菊科3種,禾本科2種,蒺藜科2種,藍雪科、豆科、百合科、檉柳科和蓼科各1種(表4)。白刺種群種子庫密度以穩定階段最高,達到9.33 粒/m2,且隨白刺灌叢植被演演替過程呈先增加后降低的趨勢,這與地上白刺種群數量及其生長狀況相一致(表1,表4)。草本植物畫眉草、苦豆子和半灌木紅砂種子在不同演替階段均有分布,在維持白刺灌叢群落穩定性中具有重要作用。1年生植物沙米是流動沙丘的指示植物,伴隨白刺灌叢退化及裸沙斑塊的出現,在衰退階段開始出現。多年生草本植物沙蔥僅在初始階段出現,而鹽爪爪僅在穩定階段出現。顯然,土壤種子庫物種組成及其種子庫密度大小反映了白刺灌叢地上植被物種組成及其種群數量。
表4 白刺灌叢植被不同演替階段土壤種子庫的物種組成及其密度
Table 4 Species composition and seed density of soil seed bank at the different succession stages ofNitrariavegetation(粒/m2, mean±SE)

植物Species科Family初始階段Initialstage穩定階段Stablestage衰退階段Degradedstage嚴重衰退階段Severelydegradedstage物種數Speciesnumber14913101年生草本Annualherb五星蒿Bassiadasyphylla藜科Chenopodiaceae0.67±0.1800.67±0.180.67±0.18沙米Agriophyllumsquarrosum藜科Chenopodiaceae000.67±0.181.33±0.18蟲實Corispermumpatelliform藜科Chenopodiaceae32.00±6.3300.67±0.180鹽生草Halogetonglomeratus藜科Chenopodiaceae7.33±1.583.33±0.84072.63±14.25刺蓬Salsolaruthenica藜科Chenopodiaceae2.00±0.4400.67±0.180.67±0.18畫眉草Eragrostispilosa禾本科Gramineae45.33±9.86496.7±134.82.67±0.5721.33±4.81砂藍刺頭Echinopsgmelini菊科Compositae0.67±0.18002.00±0.44蒿Artemisiaspp.菊科Compositae2.00±0.5500.67±0.180多年生草本Perennialherb黃花補血草Limoniumaureum藍雪科Plumbaginaceae000.67±0.185.33±1.14苦豆子Sophoraalopecuroide豆科Leguminosae6.67±1.530.67±0.1817.33±3.641.33±0.33隱子草Cleistogenessquarrosa禾本科Gramineae8.00±2.12143.33±54.2200駱駝蓬Peganumharmala蒺藜科Zygophyllaceae4.67±0.960.67±0.180.67±0.180沙蔥Alliummongolicum百合科Liliaceae0.67±0.18000半灌木Semi-shrub沙蒿Artemisiasphaerocephala菊科Compositae2.00±0.444.67±1.1112.33±3.970紅砂Reaumuriasongarica檉柳科Tamaricaceae1.33±0.370.67±0.180.67±0.180.67±0.18鹽爪爪Kalidiumfoliatum藜科Chenopodiaceae01.33±0.3700灌木Shrub白刺NitrariaTangutorum蒺藜科Zygophyllaceae4.00±0.889.33±1.936.67±1.361.33±0.32沙拐棗Calligonummongolicum蓼科Polygonaceae001.33±0.370
3.2 土壤種子庫的生活型變化
石羊河下游白刺灌叢植被土壤種子庫出現1年生草本、多年生草本、半灌木和灌木植物,未見喬木、藤本植物(表4)。其中,草本植物是土壤種子庫的主體,初始階段、穩定階段、衰退階段和嚴重衰退階段,所占物種總數比例分別達到78.6%、55.6%、69.2%和80.0%。多年生草本植物所占物種總數比例僅穩定階段達到33.3%,其他階段均未達到1/3。而半灌木、灌木植物所占物種總數比例也以穩定階段最高,達到44.4%,其他階段均未達到1/3(圖1)。顯然,伴隨白刺灌叢植被的演替過程,土壤種子庫中1年生草本植物占物種總數比例呈先減少后增加的趨勢,而多年生草本、半灌木、灌木植物占物種總數比例呈先增加后減少的趨勢。
3.3 土壤種子庫的密度變化
白刺灌叢植被不同演替演替階段的土壤種子庫密度大小明顯不同。其中,穩定階段最大,達到660.7 粒/m2,是初始階段、衰退階段和嚴重衰退階段的5.6、14.5倍和6.2倍;衰退階段最小,僅為45.7 粒/m2,占到初始階段、穩定階段和嚴重衰退階段土壤種子庫密度的38.9%、6.9%和42.5%(圖2)。
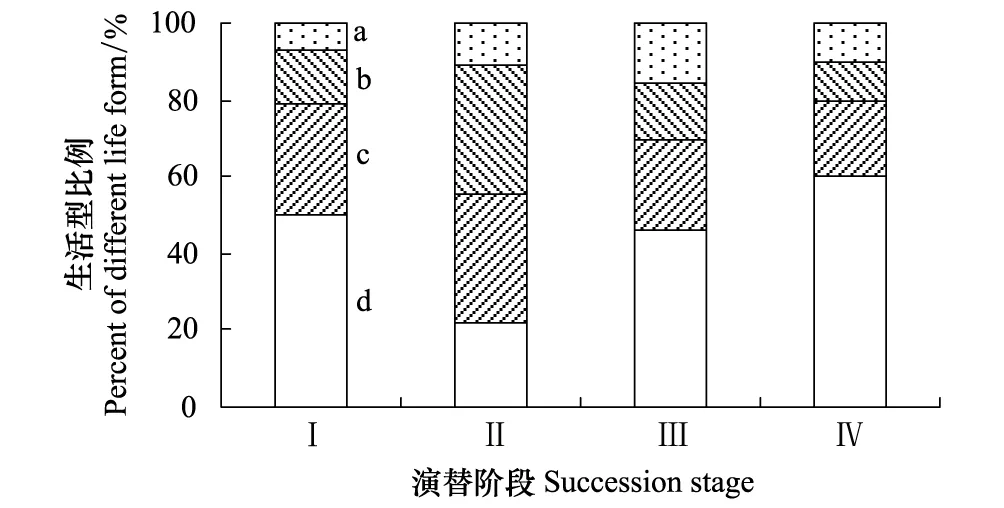
圖1 白刺灌叢植被不同演替階段土壤種子庫生活型譜Fig.1 Life form spectrum of soil seed banks at the different succession stages of Nitraria vegetation

圖2 白刺灌叢植被不同演替階段土壤種子庫密度Fig.2 Seed bank density at the different succession stages of Nitraria vegetation

圖3 白刺灌叢沙堆不同部位的土壤種子庫密度Fig.3 Seed bank density at the different positions of Nitraria nebkhas
白刺灌叢沙堆不同部位的土壤種子庫密度大小也明顯不同。其中,迎風坡中部與背風坡中部最大,分別達到329 粒/m2、309 粒/m2,與迎風坡腳、背風坡腳和堆間地差異顯著,但與沙堆頂部差異不顯著;迎風坡腳與背風坡腳土壤種子庫密度分別達到106 粒/m2和100 粒/m2,與沙堆頂部和堆間地無顯著差異(圖3)。顯然,白刺灌叢植被土壤種子庫主要分布在灌叢沙堆上,沙堆平均土壤種子庫密度達到211 粒/m2,是堆間地的1.9倍。
3.4 土壤種子庫的物種多樣性變化
白刺灌叢植被演替過程中,土壤種子庫的物種多樣性呈先降低后增加再降低的趨勢。其中,初始階段土壤種子庫的Simpson、Shannon-Wiener多樣性指數和Pielou均勻度指數最高,具有較高的物種多樣性;衰退階段土壤種子庫種子數量少,但是物種組成多,也具有較高的物種多樣性;而穩定階段與嚴重衰退階段土壤種子庫物種多樣性指數相近,土壤種子庫物種多樣性較低(表5)。

表5 白刺灌叢植被不同演替階段土壤種子庫的物種多樣性Table 5 Species diversity of soil seed bank at the different succession stages of Nitraria vegetation
3.5 土壤種子庫的相似性
白刺灌叢植被演替過程中,初始階段與其他階段的土壤種子庫相似性系數均超過0.5,處于中等相似水平。穩定階段土壤種子數量大,但物種組成數少,與衰退階段、嚴重衰退階段土壤種子庫的相似性屬于中等不相似水平。而衰退階段與嚴重衰退階段白刺灌叢退化,相似性系數達到0.53,也處于中等相似水平(表6)。
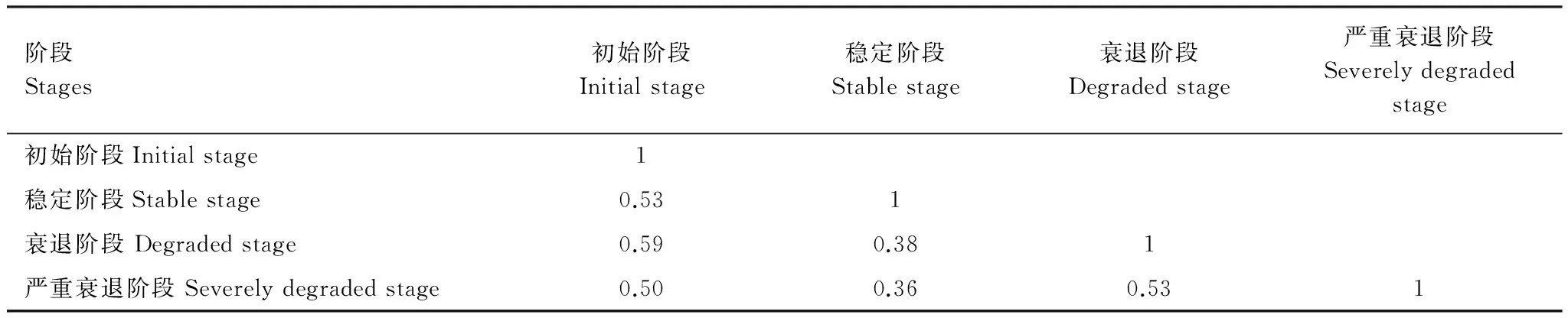
表6 白刺灌叢植被不同演替階段土壤種子庫的相似性系數Table 6 Similarity coefficients of soil seed banks between the different succession stages of Nitraria vegetation
3.6 土壤種子庫與地上植被的關系
白刺灌叢植被土壤種子庫與地上植被的共有物種數多,所占地上植被、種子庫物種的比例均超過65%。其中,初始階段共有物種數達到12種,占土壤種子庫物種數的80.0%,占地上植被物種數的66.7%;穩定階段達到8種,占種子庫物種數的88.9%,占地上植被物種數的88.9%;衰退階段達到12種,占種子庫物種數的92.3%,占地上植被物種數的75.0%;嚴重衰退階段達到7種,占種子庫種數的77.8%,占地上植被種數的77.8%。
白刺灌叢植被演替過程中,土壤種子庫與地上植被的相似性系數呈先增大后減小的趨勢,其中穩定階段的相似性系數最高,達到0.80,屬于極相似水平;初始階段、衰退階段與嚴重衰退階段也分別達到0.57、0.71和0.64,屬于中等相似水平(圖4)。顯然,石羊河下游白刺灌叢植被土壤種子庫與地上植被的相似性高,土壤種子庫較好地反映了地上植被。

圖4 白刺灌叢植被不同演替階段土壤種子庫與現有地上植被的關系Fig.4 Relationship between the standing vegetation and soil seed bank at the different succession stages of Nitraria vegetation
4 討論
土壤種子庫是指一定面積土壤中有生活力或發芽力種子的集合,包括短暫土壤種子庫和持久土壤種子庫[13,14]。在我國干旱荒漠區,7月進入雨季,荒漠植物已處于旺盛生長期,此時測得的土壤種子庫可認為持久土壤種子庫,可以反映群落的演替趨勢。不同的植被生態系統,種子在土壤中分布層次不同,土壤種子庫的取樣層次與深度不同[8,15]。對于干旱荒漠生態系統土壤種子庫,表層5 cm之下已經很少有種子,因此本研究僅采集表層0—5 cm土壤樣品研究白刺灌叢植被的土壤種子庫[8,15- 17]。
土壤種子庫是植被更新與恢復的基礎,只有具有豐富種子庫的退化生態系統,在較短的時間內能夠實現恢復[8]。不同生態系統土壤種子庫的種類組成與數量差異顯著,森林、草地的種子庫組成偏復雜,種類較多,而荒漠區的土壤種子庫組成比較簡單[8,14- 22]。地處干旱荒漠區的石羊河下游白刺灌叢植被土壤種子庫物種組成數共計18種,種子庫密度介于45.7—660.7 粒/m2,總體高于干旱、極端干旱地區沙漠[19- 20],低于半干旱、半濕潤地區沙漠[21- 22]。與騰格里沙漠南緣和科爾沁沙漠地區相比[21,23],干旱荒漠區白刺灌叢植被土壤種子庫組成相對簡單,種子庫密度與多樣性指數較低,與典型干旱、極端干旱區的研究結論相似[9,24- 25],這與白刺灌叢植被群落組成相對單一和種子植物相對較少密切相關,但根本上還是惡劣干旱、風蝕沙埋生境影響的結果。
土壤種子庫與地上植被的關系研究是土壤種子庫研究的熱點內容之一,可認識控制植被物種組成的機理,掌握群落的恢復力,控制外來物種,恢復群落多樣性,評價干預效果等,已在不同生態系統開展了大量研究[26- 29]。前期研究發現森林生態系統土壤種子庫與地上植被具有較低相似性,均值達到31%;草地生態系統具有較高相似性,均值達到54%;濕地生態系統具有中等相似性,均值達到47%;一年生植物為主導的群落土壤種子庫與地上植被的相似性高[30- 32]。本研究發現石羊河流域白刺荒漠灌叢土壤種子庫與地上植被的相似性高,其中穩定階段相似性系數達到0.80,屬于極相似水平;初始階段、衰退階段與嚴重衰退階段也分別達到0.57、0.71和0.64,屬于中等相似水平,與前期同類研究結果一致。同時,土壤種子庫與地上植被的相似性受演替階段、干擾和環境條件等的影響。Kalamees、Leck等研究發現草地系統土壤種子庫與地上植被的相似性隨演替時間降低,在演替后期階段地上植被物種組成以長命植物為主,但是種子庫由短命植物占主導,相似性降低[33- 35]。放牧等干擾活動驅動所有生態系統物種組成變化,其中森林與濕地生態系統土壤種子庫與地上植被相似性隨干擾時間降低,草地生態系統隨干擾時間增加[31- 32,36- 37]。本研究區地處石羊河下游,白刺灌叢荒漠植被沒有受到放牧等直接干擾,但是受到水資源過度開發利用的間接干擾,地表水大幅度減少和地下水位下降加速了白刺灌叢沙堆的衰退,降低了土壤種子庫與地上植被的相似性,使得石羊河流域白刺灌叢植被土壤種子庫與地上植被相似性隨演替過程呈現先增大后減小趨勢,這與前期研究有所不同。
灌叢沙堆是灌叢植被阻擋風沙流的產物,常作為干旱區土壤風蝕和土地退化的標志,間接反映了沙漠化的進程[38- 39];同時灌叢沙堆具有明顯的“肥島”效應,是水分、養分的匯集區[39- 40],具有重要的研究價值。李秋艷和方海燕研究發現泡泡刺灌叢沙堆土壤種子密度從灌叢下到灌叢間地逐漸減少,而且大部分種子分布在灌叢沙堆,幾乎沒有種子分布在灌叢間地[41];Fuentes等發現智利常綠有刺灌木叢下的土壤種子數量是灌叢間地的10倍[42];Russell和Schupp發現桃花心木灌叢下的土壤種子數量是灌叢間地的4倍[43]。本研究發現白刺灌叢植被土壤種子也主要分布于灌叢沙堆上,灌叢沙堆土壤種子數量是灌叢間地的1.9倍,從而說明灌叢沙堆也是種子的匯集區,其對種子的保存與擴散具有重要的作用。灌叢及其生長狀況、沙源和風力強度是控制沙堆形成與發育的主要因素。其中,沙堆迎風坡植被蓋度高,為氣流減速區,表現為風蝕;頂部為氣流加速區,表現為強烈侵蝕;背風坡為氣流減速區,表現為堆積[44- 45]。從而在沙堆不同部位形成了微環境,迎風坡沙堆表層沉積物粒徑相對較粗,而背風坡相對較細[46]。本研究發現白刺灌叢沙堆迎風坡與背風坡土壤種子數量大,與迎風和背風坡底差異顯著,但是迎風坡、背風坡和頂部差異不顯著,這與李秋艷和方海燕對泡泡刺灌叢沙堆種子庫的研究結論一致,說明迎風坡與背風坡環境差異并未影響種子庫數量[41]。顯然,灌叢沙堆不同部位種子庫可能受植被、風力、風蝕沙埋以及采樣時間和深度等多種因素的影響,目前對這方面的研究較為薄弱。
土壤種子庫組成、大小及其分布受植被、種子大小、繁殖方式和外部因素水分、地形、風速、動物與昆蟲采食等的綜合影響[8,47- 48]。在白刺灌叢植被演替的初始階段,灌叢沙堆最小,沙漠化程度最輕,地表沙物質組成粒度粗,適宜隱子草、沙蔥等多年生植物生長,但隨細沙物質堆積增多,就會逐漸退出白刺灌叢。在白刺灌叢植被演替的衰敗階段,隨裸沙斑塊形成,沙米種子出現,標志沙漠化程度加重。生物土壤結皮是判斷風蝕程度的重要指標,其通過阻止種子進入土壤和改變降水入滲及其再分配,影響土壤種子庫組成與結構[47,49]。在石羊河下游白刺灌叢演替過程中,灌叢沙堆的穩定主要表現為結皮的形成與逐漸加厚,限制了以分蘗擴張為生存對策的白刺灌叢的發展;灌叢沙堆的衰敗表現為結皮破損與瓦解,促進沙米、黃花補血草等草本植物的發展。
干旱荒漠白刺灌叢植被演替受到自然與人為等多種因素的影響,其中干旱區地下水位變化直接影響天然植被的生長發育,當潛水位下降到一定深度,灌叢難以從土壤中吸收到足夠的水分,植被就會衰退死亡,從而加速灌叢沙堆的衰亡[50]。本研究區域白刺灌叢植被的衰退過程主要受農業生產活動的嚴重干擾引起,沙漠邊緣大面積農田的開墾和地下水的過度開采導致地下水位快速下降,綠洲邊緣地下水位達到20 m以下,從而破壞了適宜白刺灌叢生長的生態水文條件,這是引起白刺灌叢衰退的主要原因[4- 6,10]。因此,本研究區白刺灌叢植被演替過程,特別是后期階段是一種人為干擾下的退化演變過程,土壤種子庫物種組成簡單,種子庫密度較低,退化白刺灌叢植被的自我恢復能力不足。為控制沙堆活化需要開展人工種子庫或人工植被建設,以促進耐旱植物種群的發展,是該區域白刺灌叢植被恢復和沙漠化治理的有效途徑。
5 結論
石羊河下游白刺灌叢植被演替過程中,土壤種子庫共出現9科18種植物,包括一年生草本、多年生草本、半灌木和灌木植物,其中草本植物比例達到55%—80%,是白刺灌叢植被土壤種子庫的主體。不同演替階段,土壤種子庫密度以穩定階段最高,達到660.7 粒/m2,是初始階段、衰退階段和嚴重衰退階段的5.6、14.5倍和6.2倍,初始階段、衰退階段和嚴重衰退階段間差異不顯著。白刺灌叢植被土壤種子庫主要分布于灌叢沙堆,沙堆土壤種子庫密度是堆間地的1.9倍,其中沙堆迎風坡中部與背風坡中部土壤種子庫密度最高,分別達到329 粒/m2和309 粒/m2。不同演替階段土壤種子庫間的相似性系數均在0.6以下,除穩定階段外,其他階段間均處于中等相似水平;土壤種子庫與地上植被的相似性系數均在0.5以上,達到中等相似或極相似水平。該研究結果進一步證明了灌叢沙堆在白刺灌叢植被中的重要性,也說明人為干擾損害了白刺灌叢地下潛在植被及其恢復潛力。
[1] Milberg P, Hansson M L. Soil seed bank and species turnover in a limestone grassland. Journal of Vegetation Sciences, 1993, 5(1): 35- 42.
[2] 李秋艷, 趙文智. 干旱區土壤種子庫的研究進展. 地球科學進展, 2005, 20(3): 350- 358.
[3] 李雙福, 張啟昌, 張啟超, 宗成武, 田憲鋒. 白刺屬植物研究進展. 北華大學學報: 自然科學版, 2005, 6(1): 78- 81.
[4] 孫祥, 于卓. 白刺根系的研究. 中國沙漠, 1992, 12(4): 50- 54.
[5] 賈寶全, 蔡體久, 高志海, 丁峰, 張國忠. 白刺灌叢沙包生物量的預測模型. 干旱區資源與環境, 2002, 16(1): 96- 99.
[6] 彭飛, 王濤, 劉立超, 黃翠華. 民勤荒漠綠洲過渡帶白刺灌叢沙堆演化階段及其空間格局. 中國沙漠, 2012, 32(3): 593- 599.
[7] 劉建泉. 甘肅民勤西沙窩唐古特白刺群落的生態特性. 植物資源與環境學報, 2002, 11(3): 36- 40.
[8] 楊自輝, 高志海. 荒漠綠洲邊緣降水和地下水對白刺群落消長的影響. 應用生態學報, 2000, 11(6): 927- 930.
[9] 杜建會, 嚴平, 展秀麗, 俄有浩. 民勤綠洲白刺灌叢沙堆不同演化階段表面抗蝕性及其影響因素. 應用生態學報, 2008, 19(4): 763- 768.
[10] 靳虎甲, 馬全林, 張有佳, 曾新德. 石羊河下游白刺灌叢演替發育過程的土壤呼吸及其影響因素分析. 中國沙漠, 2012, 32(1): 140- 147.
[11] 馬克平, 劉玉明. 生物群落多樣性的測度方法 Ⅰ a多樣性的測度方法 (下). 生物多樣性, 1994, 2(4): 231- 239.
[12] 沈蕊, 張建利, 何彪, 李峰, 張志明, 周睿, 歐曉昆. 元江流域干熱河谷草地植物群落結構特征與相似性分析. 生態環境學報, 2010, 19(12): 2821- 2825.
[13] 于順利, 陳宏偉, 郎南軍. 土壤種子庫的分類系統和種子在土壤中的持久性. 生態學報, 2007, 27(5): 2099- 2108.
[14] Coffin D P, Lauenroth W K. Spatial and temporal variation in the seed bank of a semiarid grassland. American Journal of Botany, 1989, 76(1): 53- 58.
[15] Marone L, Cueto V R, Milesi F A, de Casenave J L. Soil seed bank composition over desert microhabitats: patterns and plausible mechanisms. Canadian Journal of Botany, 2004, 82(12): 1809- 1816.
[16] Wang S M, Zhang X, Li Y, Zhang L, Xiong Y C, Wang G. Spatial distribution patterns of the soil seed bank ofStipagrostispennata(Trin.) de Winter in the Gurbantonggut desert of north-west China. Journal of Arid Environments, 2005, 63(1): 203- 222.
[17] 馬全林, 張德魁, 劉有軍, 靳虎甲, 陳芳. 石羊河中游沙漠化逆轉過程土壤種子庫的動態變化. 生態學報, 2011, 31(4): 989- 997.
[18] 張志權. 土壤種子庫. 生態學雜志, 1996, 15(6): 36- 42.
[19] Li N, Feng G, Tian C Y. Characteristics and dynamics of the soil seed bank at the north edge of Taklimakan desert. Science in China Series D: Earth Sciences, 2007, 50(S1): 122- 127.
[20] 王增如, 徐海量, 尹林克, 李吉玫, 張占江, 李媛. 塔里木河下游荒漠化過程土壤種子庫特征. 中國沙漠, 2009, 29(5): 885- 889.
[21] 趙麗婭, 李鋒瑞, 張華, 王先之. 科爾沁沙地圍封沙質草甸土壤種子庫特征的研究. 生態學雜志, 2004, 23(2): 45- 49.
[22] 沈有信, 趙春燕. 中國土壤種子庫研究進展與挑戰. 應用生態學報, 2009, 20(2): 467- 473.
[23] 王剛, 梁學功. 沙坡頭人工固沙區的種子庫動態. 植物學報, 1995, 37(3): 231- 237.
[24] 曾彥軍, 王彥榮, 南志標, 衛東, 陳善科, 李保爾. 阿拉善干旱荒漠區不同植被類型土壤種子庫研究. 應用生態學報, 2003, 14(9): 1457- 1463.
[25] 李吉玫, 徐海量,張占江, 葉茂, 王增如, 李媛. 塔里木河下游不同退化區地表植被和土壤種子庫特征. 生態學報, 2008, 28(8): 3626- 3636.
[26] Carter C T, Ungar I A. Aboveground vegetation, seed bank and soil analysis of a 31-year-old forest restoration on coal mine spoil in southeastern Ohio. American Midland Naturalist, 2002, 147(1): 44- 59.
[27] Amiaud B, Touzard B. The relationships between soil seed bank, aboveground vegetation and disturbances in old embanked marshlands of western France. Flora-Morphology, Distribution, Functional Ecology of Plants, 2004, 199(1): 25- 35.
[28] Bossuyt B, Hermy M. Seed bank assembly follows vegetation succession in dune slacks. Journal of Vegetation Science, 2004, 15(4): 449- 456.
[29] Kirkham F W, Kent M. Soil seed bank composition in relation to the above-ground vegetation in fertilized and unfertilized hay meadows on a Somerset peat moor. Journal of Applied Ecology, 1997, 34(4): 889- 902.
[30] Osem Y, Perevolotsky A, Kigel J. Similarity between seed bank and vegetation in a semi-arid annual plant community: the role of productivity and grazing. Journal of Vegetation Science, 2006, 17(1): 29- 36.
[31] Hopfensperger K N. A review of similarity between seed bank and standing vegetation across ecosystems. Oikos, 2007, 116(9): 1438- 1448.
[32] 李鋒瑞, 趙麗婭, 王樹芳, 王先之. 封育對退化沙質草地土壤種子庫與地上群落結構的影響. 草業學報, 2003, 12(4): 90- 99.
[33] Kalamees R, Zobel M. Soil seed bank composition in different successional stages of a species rich wooded meadow in Laelatu, western Estonia. Acta Oecologica, 1998, 19(2): 175- 180.
[34] Chambers J C. Seed and vegetation dynamics in an alpine herb field: effects of disturbance type. Canadian Journal of Botany, 1993, 71(3): 471- 485.
[35] Leck M A, Leck C F. A ten-year seed bank study of old field succession in central New Jersey. Journal of the Torrey Botanical Society, 1998, 125(1): 11- 32.
[36] Chaideftou E, Thanos C A, Bergmeier E, Kallimanis A, Dimopoulos P. Seed bank composition and above-ground vegetation in response to grazing in sub-Mediterranean oak forests (NW Greece). Plant Ecology, 2009, 201(1): 255- 265.
[37] Chang E R, Jefferies R L, Carleton T J. Relationship between vegetation and soil seed banks in an arctic coastal marsh. Journal of Ecology, 2001, 89(3): 367- 384.
[38] Tengberg A. Nebkhas dunes as indicators of wind erosion and land degradation in the Sahel zone of Burkina Faso. Journal of Arid Environments, 1995, 30 (3): 265- 282.
[39] 杜建會, 嚴平, 董玉祥. 干旱地區灌叢沙堆研究現狀與展望. 地理學報, 2010, 65(3): 339- 350.
[40] Seifert C L, Cox R T, Forman S L, Foti T L, Wasklewicz T A, McColgan A T. Relict nebkhas (pimple mounds) record prolonged late Holocene drought in the forested region of south-central United States. Quaternary Research, 2009, 71(3): 329- 339.
[41] 李秋艷, 方海燕. 荒漠綠洲邊緣區泡泡刺灌叢土壤種子庫研究. 干旱區研究, 2008, 25(4): 502- 506.
[42] Fuentes E R, Otaiza R D, Alliende M C, Hoffmann A, Poiani A. Shrub clumps of the Chilean matorral vegetation: structure and possible maintenance mechanisms. Oecologia, 1984, 62(3): 405- 411.
[43] Russell S K, Schupp E W. Effects of microhabitat patchiness on patterns of seed dispersal and seed predation ofCercocarpusledifolius(Rosaceae). Oikos, 1998, 81(3): 434- 443.
[44] Brown G, Porembski S. The maintenance of species diversity by miniature dunes in a sand-depletedHaloxylonsalicornicumcommunity in Kuwait. Journal of Arid Environments, 1997, 37(3): 461- 473.
[45] 武勝利, 李志忠, 惠軍, 肖晨曦, 孫秋梅. 灌叢沙堆表面壓力分布特征的實驗研究. 干旱區地理, 2006, 29(6): 790- 796.
[46] 張萍, 哈斯, 岳興玲, 莊燕美. 白刺灌叢沙堆形態與沉積特征. 干旱區地理, 2008, 31(6): 926- 932.
[47] Guo Q F, Rundel P W, Goodall D W. Horizontal and vertical distribution of desert seed banks: Patterns, causes, and implications. Journal of Arid Environments, 1998, 38(3): 465- 478.
[48] Li X R, Ma F Y, Xiao H L, Wang X P, Kim K C. Long-term effects of revegetation on soil water content of sand dunes in arid region of northern China. Journal of Arid Environments, 2004, 57(1): 1- 16.
[49] 蘇延桂, 李新榮, 張景光, 楊麗雯. 生物土壤結皮對土壤種子庫的影響. 中國沙漠, 2006, 26(6): 997- 1001.
[50] 鄭丹, 李衛紅, 陳亞鵬, 劉加珍. 干旱區地下水與天然植被關系研究綜述. 資源科學, 2005, 27(4): 160- 167.
Varying characteristics of soil seed banks during the succession process ofNitrariaTangutorumvegetation in an arid desert area
MA Quanlin1,2,*,LU Qi1,WEI Linyuan2,JIN Hujia2
1InstituteofDesertificationStudies,ChineseAcademyofForestry,Beijing100091,China2StateKeyLaboratoryBreedingBaseofGansuDesertificationandAeolianSandDisasterCombating,GansuDesertControlResearchInstitute,Lanzhou730070,China
The seed bank is the collection of viable seeds present on or within the soil at any given time. Soil seed bank is an important indicator in response to changes in land utilization and climate, and also forms the basis for desert vegetation restoration and development.Nitrariavegetation is typical of desert vegetation and with nebkhas is widely distributed along the periphery of oasis in the arid regions of Northwest China, playing an important role in desertification control around oases. In recent decades,Nitrariavegetation in the oasis-desert ecotone has been seriously degraded due to the rapid decrease of the groundwater table caused by agricultural over development. However, there is little information regarding the change in characteristics of the soil seed bank ofNitrarianebkhas in arid desert areas. Using the method of substituting space for time, the species composition, distribution pattern and quantitative variation of soil seed banks ofN.Tangutorumnebkhas in initial, stable, degraded and severely degraded stages were studied in the lower reaches of the Shiyang River. Results showed that the soil seed bank ofN.Tangutorumnebkha in the lower reaches of the Shiyang River was composed of 18 species, belonging to 9 families. Herb species accounted for 55%—80% and were the main body of soil seed banks. The soil seed density of the stable stage was 660.7 seeds/m2, and was the highest, being 5.6, 14.5 and 6.2 times greater than those measured in the initial stage, degraded stage and severely degraded stage, respectively. The soil seed density of the degraded stage was 45.7 seeds/m2, and was the lowest, only accounting for 38.9%, 6.9% and 42.5% of those measured in the initial stage, stable stage and severely degraded stage, respectively. The soil seed density of theN.Tangutorumpopulation at the different succession stages was also significantly different, showing a decreasing trend after the initial increase. The soil seed density ofN.Tangutorumpopulation at the stable stage reached 9.33 seeds/m2. The soil seed density in different positions ofN.Tangutorumnebkhas was significantly different and was mainly distributed in the central parts of the windward and leeward slopes ofN.Tangutorumnebkhas, with seed densities of 329 seeds/m2and 309 seeds/m2, respectively. The mean soil seed density of the nebkhas was 1.9 times that in the depressions within the nebkhas. Similarity coefficients of soil seed banks between the different succession stages were less than 0.6, which were recognized as having a moderate similarity level except for the stable stage which showed a low similarity coefficient. There were more common species which accounted for over 65% of vegetation above ground and the soil seed bank. Similarity coefficients between vegetation above ground and the soil seed bank were more than 0.5, which showed a moderate to extreme similarity level, and demonstrated a decreasing trend after the initial increase. The findings have proven the importance of soil seed banks andNitrarianebkhas inNitrariavegetation and also suggest that human disturbance has impacted potential underground vegetation and its restoration capacity. Therefore, man-made soil seed banks or vegetation construction should be adopted to containNitrarianebkhas activation and restore degradedNitrariavegetation in arid desert areas.
the lower reaches of the Shiyang River;Nitrariavegetation; nebkhas; soil seed bank; similarity
國家973項目(2012CB723203); 國家自然科學基金項目(41061030, 31270754); 中國博士后科學基金項目(20110490474, 2012T50162); 國家科技支撐計劃(2012BAD16B0203)
2013- 06- 04;
日期:2014- 05- 08
10.5846/stxb201306041326
*通訊作者Corresponding author.E-mail: mql925@126.com
馬全林,盧琦,魏林源,靳虎甲.干旱荒漠白刺灌叢植被演替過程土壤種子庫變化特征.生態學報,2015,35(7):2285- 2294.
Ma Q L,Lu Q,Wei L Y,Jin H J.Varying characteristics of soil seed banks during the succession process ofNitrariaTangutorumvegetation in an arid desert area.Acta Ecologica Sinica,2015,35(7):2285- 2294.
猜你喜歡
課堂內外·初中版(科學少年)(2025年1期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2025年2期)2025-02-28 00:00:00
課堂內外·初中版(科學少年)(2024年12期)2024-12-02 00:00:00
英語世界(2023年10期)2023-11-17 09:18:18
中學生博覽(2022年7期)2022-06-21 21:48:14
大科技·百科新說(2021年8期)2021-11-03 10:55:16
學苑創造·A版(2021年5期)2021-06-28 19:51:42
少兒美術(快樂歷史地理)(2020年9期)2020-03-19 05:10:56
科學大眾(中學)(2019年3期)2019-05-17 10:04:30
汽車觀察(2018年10期)2018-11-06 07:05:26
