血小板衍生生長因子調節休克血管反應性的非MLC20磷酸化機制與HSP27和Caldesmon的關系
2015-03-01 01:55:18彭小勇
創傷外科雜志 2015年2期
朱 娛,彭小勇,藍 丹,李 濤
嚴重創傷/休克晚期常出現血管的低反應性,表 現為全身血管對縮血管物質或舒血管物質的反應降低或不反應,是導致休克后期血壓不能有效提升、組織灌注難以改善、組織缺氧和損傷進行性加重的重要原因[1-2]。已有研究發現血管低反應性的發生與腎上腺能受體失敏、血管平滑肌細胞(vascular smooth muscle cell,VSMC)鉀、鈣功能失常及細胞膜超極化有關[3]。這些機制均通過抑制MLC20磷酸化發揮作用。近來研究表明:非MLC20磷酸化也參與血管平滑肌細胞的收縮。Lee等[4]報道了ET-1誘導兔子主動脈的收縮,但MLC20磷酸化水平沒有增加;Su等[5]發現組胺能使豬平滑肌細胞收縮,PI3K的抑制劑wortmannin能減少組胺引起的平滑肌細胞收縮,但不減少MLC20磷酸化水平。這些結果均表明非MLC20磷酸化通路參與平滑肌細胞收縮的調節。然而,非MLC20磷酸化通路是否參與休克后血管低反應性的發生,以及怎樣調節血管低反應性,尚不清楚。
許多研究證實:Caldesmon和27-kDa的熱休克蛋白(HSP27)通過調節肌球蛋白和肌動蛋白(不涉及MLC20磷酸化通路),參與平滑肌細胞的收縮。Caldesmon是調節細肌絲的蛋白,其C端結合肌動蛋白,抑制肌球蛋白ATP酶活性,導致平滑肌收縮減弱[6-9]。HSP27是熱休克蛋白家族的一員,廣泛分布于不同組織和細胞中。在平滑肌和心肌細胞,HSP27作為肌動蛋白的分子伴侶與細肌絲一起參與細胞骨架的調節。Caldesmon和HSP27是否通過非MLC20磷酸化參與休克后血管低反應性的發生尚不清楚。
血小板衍生的生長因子(platelet derived-growth factor,PDGF)是血管內皮生長因子中的重要成員,在許多組織和細胞中均有表達,包括血管內皮細胞、巨噬細胞和單核細胞[10]。PDGF除了在急性損傷和一些慢性損傷中能夠加速組織修復和傷口愈合外,還能引起血管平滑肌細胞收縮[11-13]。通過預實驗發現,PDGF能夠激活平滑肌細胞中Caldesmon和HSP27的表達。因此本實驗采用失血性休克和缺氧處理的腸系膜上動脈(SMA)作為模型,來探討PDGF在血管低反應中調節非MLC20磷酸化的機制,以及與Caldesmon和HSP27的關系。
材料與方法
1 動物模型及方法
動物模型:SD大鼠(200~250g),雌雄不拘,由第三軍醫大學第三附屬醫院實驗動物中心提供(所有動物遵照第三軍醫大學實驗動物指南規范操作),禁食過夜,自由飲水,3%戊巴比妥鈉麻醉,行左股動脈插管,用于監測血壓。插管后穩定10min,從左股動脈抽血使平均動脈壓降至40mmHg,維持2h,制成失血性休克模型。
血管的準備:從休克或假手術組的大鼠取出SMA,去掉周圍的結締組織后,將其切成2~3mm長的動脈環,用于張力測定。根據實驗要求,對血管環行缺氧或不缺氧處理,缺氧組血管環放到缺氧罐中,95%N2和 5%CO2以 10L/min充氧 15min,平衡10min,重復3次直到氧濃度<0.2%,維持2h,制成缺氧模型。
血管反應性測定:將已處理的血管環懸掛于一對不銹鋼絲上,一端置于固定柱上,另一端與肌張力換能器相連,放入注有K-H液[Krebs-Henseleit solution,組成(mM):NaCl 118、KCl 4.7、NaHCO325、KH2PO4 1.03、MgSO4·7H2O 0.45、CaCl22.5、葡萄糖11.1、pH 7.4]的離體器官灌流浴槽中,持續充入95%O2和5%CO2混合氣體,給予血管環初張力0.5g、37℃恒溫孵育120min,每30min換液1次,待張力曲線平穩后,依次加入終濃度分別為 10-9、10-8、10-7、10-6、10-5和 10-4mol/L 的去甲腎上腺素(NE)。血管環對NE的反應性用濃度累計法測定,記錄不同NE濃度下各血管環產生的最大收縮力,以收縮力/血管環重量(g/mg組織)為量化標準,作量-效曲線和Emax圖評價血管收縮反應性。
MLC20磷酸化:取大鼠的腸系膜動脈置于含丙酮、二硫蘇糖醇(DTT)的保存液中-70℃保存固定3d,然后將血管晾干后剪碎,加蛋白提取液混勻后在搖床上振蕩1.5h,15 000rmp離心15min后取上清,即為蛋白。
HSP27和Caldesmon的表達:制備蛋白樣品后置于-70℃保存備用。變性后用10%SDS-PAGE凝膠電泳分離蛋白,濕轉法將蛋白轉移到PVDF膜上,用5%脫脂奶粉封閉2h后,三羥甲基氨基甲烷緩沖液(TBST)洗滌3次,用HSP27和HSP27磷酸化(p-HSP27)抗體1∶1000,CaD和 CaD磷酸化(p-CaD)抗體1∶1000孵育4℃過夜,TBST洗滌3次后,與二抗1∶10000室溫孵育1h,TBST洗滌4次后,使用電化學發光液(ECL)進行反應,暗室曝光。使用β-肌動蛋白(β-actin)作為陽性對照。
2 實驗步驟
2.1 非MLC20磷酸化途徑在PDGF調節休克后血管反應性中的作用 將48只大鼠隨機分成6組,每組8只,分別為:假手術組、休克組、休克+PDGF(40、60、80、100ng/ml)組。SMA 用于測定對 NE 的反應性[14](AD 設備,Castle Hill,NSW,澳大利亞),剩下的腸系膜動脈用于測定MLC20磷酸化水平。在休克+PDGF(40、60、80、100ng/ml)組,SMA 和腸系膜動脈均孵育 PDGF(40、60、80、100ng/ml)15min,再觀察血管的收縮反應和MLC20磷酸化水平。用甘油聚丙烯凝膠電泳測定MLC20磷酸化水平,用累積的去甲腎上腺素濃度來反映SMA的反應性[15]。
2.2 Caldesmon和HSP27在PDGF調節血管反應性中的作用及與MLC20途徑的關系 為了觀察Caldesmon和HSP27在PDGF是否通過非MLC20磷酸化調節血管的收縮反應中的作用,用不同濃度的PDGF作用后,觀察 Caldesmon(p-Caldesmon)和HSP27(p-HSP27)的蛋白水平變化,以及用Caldesmon和HSP27的反義寡核苷酸(AODN)抑制后(CaldesmonAODN序列:5-TCTCTTCCTCCTCCTCCTCCT-3';HSP27AODN序列:5-ATCTCCACCACGCCTTCCTT-3'),觀察PDGF在非MLC20磷酸化中調節血管的收縮反應性和MLC20磷酸化水平。Caldesmon或HSP27AODN(終濃度100u M)與轉染試劑以體積比5∶1轉染正常大鼠血管環,24h后行缺氧處理,同時用100ng/ml PDGF孵育血管環。實驗當天,血管環按上述方法缺氧,觀察SMA對NE的收縮反應及Caldesmon和HSP27的表達情況。
3 統計分析
結 果
1 非MLC20磷酸化途徑在PDGF調節休克后血管反應性中的作用
血管反應性:與假手術組比較,休克后SMA的血管反應性顯著下降,不同濃度的 PDGF(40、60、80、100ng/ml)可以改善血管反應性,呈劑量依賴性;用不同濃度 PDGF(40、60、80、100ng/ml)孵育后,血管反應性從休克組的0.708(g/mg組織)分別增加到1.232、1.371、1.671、1.803(g/mg 組織)。與休克組比較,PDGF不同濃度增加血管反應性比例分別為74.01%、93.64%、136.01%、154.66%(圖1)。

圖1 不同濃度PDGF對血管反應性的影響
MLC20磷酸化:與假手術組比較,腸系膜動脈的MLC20磷酸化顯著降低,從假手術組的32.56%降低到休克組10.26%。不同濃度的PDGF增加MLC20磷酸化,但PDGF不同濃度間的MLC20磷酸化水平無顯著性差異,分別為 30.23%(40ng/ml)、31.25%(60ng/ml)、31.02%(80ng/ml)、31.56%(100ng/ml)(圖2)。
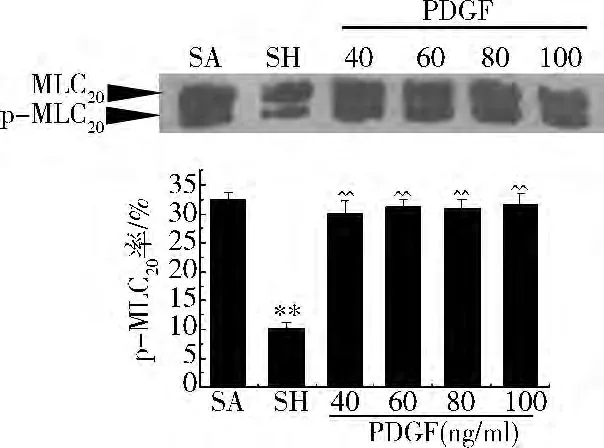
圖2 不同濃度PDGF的MLC20磷酸化
2 Caldesmon和HSP27在非MLC20磷酸化途徑中調節血管反應性的作用
Caldesmon和HSP27磷酸化:在對照組、休克組和不同濃度的PDGF組中,Caldesmon和HSP27的蛋白表達無顯著性變化,但休克后Caldesmon的磷酸化增加而 HSP27的磷酸化降低,不同濃度的PDGF(40、60、80和100ng/ml)可以逐漸降低休克引起的Caldesmon磷酸化的增加,而增加休克導致的HSP27磷酸化的下降(圖3)。
血管反應性:與對照組比較,缺氧后血管反應性降低,PDGF100ng/ml可以增加SMA的血管反應性,HSP27AODN可以抑制PDGF引起的血管反應性的增加,CaldesmonAODN可進一步增加PDGF誘導的血管反應性的增加(圖4)。
MLC20磷酸化:與對照組比較,休克組的腸系膜動脈的MLC20磷酸化降低,PDGF100ng/ml可以增加休克后腸系膜動脈的MLC20磷酸化,HSP27和CaldesmonAODN不能抑制PDGF引起的MLC20磷酸化的增加(圖5)。
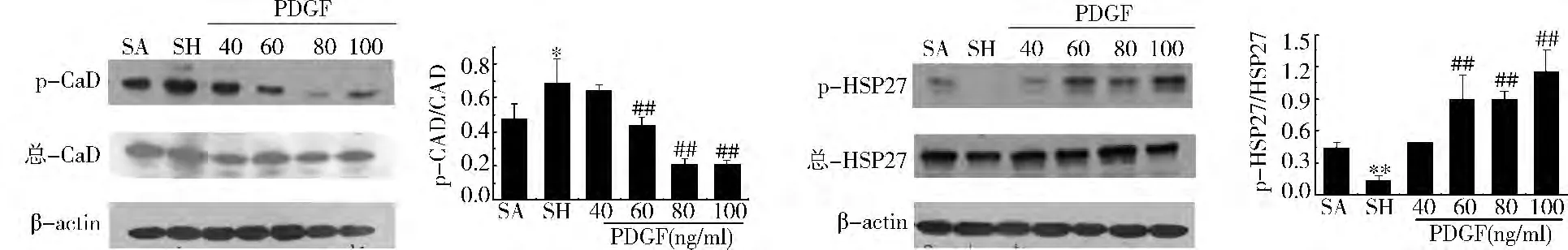
圖3 不同濃度PDGF對Caldesmon和HSP27磷酸化水平的影響
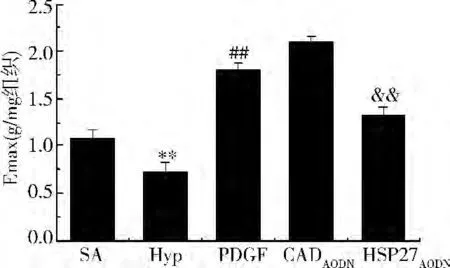
圖4 HSP27AODN和CaldesmonAODN在PDGF調節休克血管反應性中的作用

圖5 HSP27AODN和CaldesmonAODN對MLC20磷酸化的影響
討 論
血管低反應嚴重影響著疾病的發生、發展和治療,它是許多危重癥的重要死亡原因之一。目前的研究表明休克后血管低反應性的發生機制與平滑肌細胞受體失敏、膜超極化以及我們實驗室提出的鈣失敏有關。嚴重創傷或休克后,高濃度的血管活性物質,大量的一氧化氮(NO)、內皮素、腫瘤壞死因子(TNF)、白細胞介素(IL)或內源性的阿片肽引起受體降解、內化或去磷酸化;在休克或創傷時,三磷酸腺苷(ATP)缺乏導致了ATP敏感的ATP依賴的鉀離子通道(KATP)激活,細胞內鈣濃度降低,MLC20磷酸化和血管反應性下降。我們先前的實驗進一步證明了鈣敏感性的降低,Rho蛋白(Rho kinase)和蛋白激酶 C(PKC)通過抑制 MLCP的活性,增加MLC20磷酸化調節鈣敏感性[16-18]。這些引起休克后血管低反應性的機制主要是通過降低MLC20磷酸化水平從而降低血管的收縮反應。
目前的研究表明,非MLC20磷酸化通路參與了休克后血管低反應性的發生。Caldesmon和HSP27也參與休克后血管低反應性的調節。Caldesmon是一種特殊的肌動蛋白結合蛋白,在體內廣泛存在,分為輕型和重型。重型鈣調蛋白結合蛋白只在血管和內臟平滑肌以及肌上皮細胞有表達,其C端包括肌動蛋白結合域,原肌球蛋白結合域和鈣調蛋白結合域,在N端具有肌球蛋白結合域[18]。重型鈣調蛋白不僅可以起到抑制肌球蛋白Mg-ATP酶活性的作用,還可以抑制平滑肌磷酸化肌球蛋白所引發的肌動蛋白絲的移動,是一種重要的調節蛋白。Caldesmon磷酸化后不再抑制ATP酶活性,有利于肌肉收縮。當平滑肌受外界刺激時,可通過特異性激活PKCε,啟動促分裂原活化蛋白激酶(MAPK)途徑,通過一系列級聯反應,引起Caldesmon間接磷酸化,進而調節平滑肌的收縮。
HSP27是小分子量HSPs(small HSPs,sHSPs)亞家族中的重要一員,具有保護細胞免受各種應激因素的損傷的功能,促進蛋白質的正確折疊;與細胞的增殖、分化及細胞凋亡的信號轉導調節密切相關。在多種應激條件下,HSP27作為分子伴侶發揮保護細胞的功能。黃捷等[19]研究結果顯示HSP27直接參與了PDGF誘導的細胞遷移過程,論證了HSP27是血管平滑肌細胞遷移的關鍵調節因子。
本實驗結果證實,休克后血管反應性明顯降低,不同濃度PDGF能劑量依賴地增加休克血管反應性,卻不能劑量依賴地增加MLC20磷酸化水平。休克后Caldesmon的磷酸化增加而HSP27的磷酸化降低,不同濃度的 PDGF(40、60、80和100ng/ml)可以逐漸降低休克引起的Caldesmon磷酸化的增加,而逐漸增加休克導致的HSP27磷酸化的下降。說明PDGF調節休克血管反應性的非MLC20磷酸化機制與HSP27和Caldesmon相關。
[1]陳墾,劉良明.缺氧誘導因子1ɑ對失血性休克大鼠腸系膜上動脈血管環舒張反應性的調控作用[J].第三軍醫大學學報,2010,32(4):319 -323.
[2]Liu LM,Yang GM,Zhu Y,et al.Role of non - MLC20 phosphorylation pathway in the regulation of vascular reactivity during shock[J].J Surg Res,2014,187(2):571 -580.
[3]Liang JL,Yang GM,Li T,et al.Interleukin 1βattenuates vascular α1 adrenergic receptors expression following lipopolysaccharide-induced endotoxemia in rabbits:involvement of JAK2 - STAT3 pathway[J].J Trauma Acute Care Surg,2014,76(3):762 -770.
[4]Lee HM,Won KJ,Kim J,et al.Endothelin - 1 induces contraction via a syk-mediated p38 mitogen-activated protein kinase pathway in rat aortic smooth muscle[J].J Pharmacol Sci,2007,103(4):427 -434.
[5]Su XL,Smolock EM,Marcel KN,et al.Phosphatidylinositol 3-kinase modulates vascular smooth muscle contraction by calcium and myosin light chain phosphorylation-independent and-dependent pathways[J].Am J Physiol Heart Circ Physiol,2004,286(2):657 -666.
[6]周琴,易光輝.HSP27對動脈損傷的細胞保護機制[J].中國優生與遺傳雜志,2010,18(11):137-139.
[7]Lee YR,Lee CK,Park HJ,et al.c - Jun N-terminal kinase contributes to norepinephrine-induced contraction through phosphorylation of caldesmon in rat aortic smooth muscle[J].J Pharmacol Sci,2006,100(2):119 -125.
[8]徐彥龍,高靚,等.針刺調節腦梗死大鼠腦血管平滑肌CaP、CaD變化的實驗研究[J].遼寧中醫雜志,2012,39:(9):1864-1866.
[9]Mayanagi T,Sobue K.Diversification of caldesmon-linked actin cytoskeleton in cell motility[J].Cell Adh Migr,2011,5(2):150 -159.
[10]Hoch RV,Soriano P.Roles of PDGF in animal development[J].Development,2003,130(20):4679 - 4784.
[11]Judith R,Nithya M,Rose C,et al.Application of a PDGF- containing novel gel for cutaneous wound healing[J].Life Sci,2010,87(1):1 -8.
[12]Mulder G,Tallis AJ,Marshall VT,et al.Treatment of nonhealing diabetic foot ulcers with a platelet-derived growth factor gene-activated matrix(GAM501):results of a phase 1/2 trial[J].Wound Repair Regen,2009,17(6):772-779.
[13]Zhao Y,Lv W,Piao H,et al.Role of platelet- derived growth factor-BB(PDGF-BB)in human pulmonary artery smooth muscle cell proliferation[J].J Recept Signal Transduct Res,2014,[Epub ahead of print].
[14]Li T,Yang GM,Zhu Y,et al.The mechanism by which RhoA regulatesvascularreactivity afterhemorrhagic shock in rats[J].Am J Physiol,2010,299(2):H292 -299.
[15]劉嘉,梁新華,焦炎.小鼠胃和闌尾組織中h-CD降解規律與死亡時間的相關性究[J].中國醫學導報,2011,8(6):17 -19.
[16]Li T,Fang YQ,Zhu Y,et al.Beneficial effect of activation of PKC on hemorrhagic shock in rats[J].J Trauma,2010,68(4):865 -873.
[17]閔清,白育庭,劉晶,等.山楂葉總黃酮對大鼠血管環的作用及其機制探討[J].中國藥理學通報,2011,27(4):585-590.
[18]Li T,Fang YQ,Yang GM,et al.Effects of the balance in activity of RhoA and Rac1 on the shock-induced biphasic change of vascular reactivity in rats[J].Ann Surg,2011,253(1):185 -193.
[19]黃捷,謝良地,許昌聲,等.RNA干擾熱休克蛋白27基因對自發性高血壓大鼠血管平滑肌細胞增殖、遷移的影響[J].中華高血壓雜志,2010,18(8):761 -764.
