鎮(zhèn)海水庫擬柱孢藻的分離鑒定和氮磷對其生長的影響
2015-02-27 08:10:46戴景峻雷臘梅
水生生物學(xué)報 2015年3期
關(guān)鍵詞:生長
戴景峻 彭 亮 于 婷 雷臘梅
(暨南大學(xué)生態(tài)學(xué)系, 水體富營養(yǎng)化與赤潮防治廣東普通高校重點實驗室, 廣州 510632)
鎮(zhèn)海水庫擬柱孢藻的分離鑒定和氮磷對其生長的影響
戴景峻 彭 亮 于 婷 雷臘梅
(暨南大學(xué)生態(tài)學(xué)系, 水體富營養(yǎng)化與赤潮防治廣東普通高校重點實驗室, 廣州 510632)
以分離自廣東省鎮(zhèn)海水庫的擬柱孢藻 N8為對象, 探究其在不同磷濃度及氮磷濃度組合下的生長情況。結(jié)果表明, 擬柱孢藻N8對磷的適應(yīng)范圍很寬, 在0.02—5.12 mg/L磷濃度下均能生長, 最適生長磷濃度范圍為0.16—5.12 mg/L, 磷濃度的升高能顯著延長擬柱孢藻的對數(shù)生長期和提高生物量。動力學(xué)分析表明,擬柱孢藻N8有較低的KSP值, 對磷元素的親和性較高, 在磷營養(yǎng)貧乏的環(huán)境下更容易形成優(yōu)勢。在氮磷組合實驗中, 低氮(0.5 mg/L)顯著抑制擬柱孢藻的生長, 且這種生長抑制不受磷濃度的影響; 而在低磷(0.04 mg/L)條件下, 水體中氮濃度的增加會顯著促進(jìn)擬柱孢藻的生長, 擬柱孢藻在高氮中磷和高氮高磷下的生長顯著優(yōu)于其他氮磷組合條件。研究表明, 廣東省水庫擬柱孢藻的生長受磷的限制較弱, 氮是其生長的決定因子。
鎮(zhèn)海水庫; 擬柱孢藻; 氮磷營養(yǎng)鹽; 生長特性; 南亞熱帶
擬柱孢藻 (Cylindrospermopsis raciborskii) 是一種具有固氮能力的絲狀水華藍(lán)藻, 1978年于匈牙利巴拉頓湖首次被記錄[1]。該藻被認(rèn)為是熱帶亞熱帶地區(qū)的特征性種類, 有大量的報道稱其在澳大利亞北部、南美洲、非洲等國家或地區(qū)的湖泊和水庫中存在并成為優(yōu)勢種[1,2]。在過去 20多年, 擬柱孢藻在溫帶地區(qū)出現(xiàn)的頻率明顯增加, 已經(jīng)遍布整個歐洲和澳大利亞, 近年來在美國、波蘭、日本、加拿大、新西蘭等國家的水體中都有發(fā)現(xiàn)擬柱孢藻[3—9]。研究認(rèn)為擬柱孢藻具有較強(qiáng)的適應(yīng)能力, 隨著全球氣候變暖, 其種群分布不斷由熱帶亞熱帶地區(qū)向溫帶地區(qū)擴(kuò)散[10]; 但作為熱帶特征性種類, 其最佳生長溫度普遍在25℃以上, 因此擬柱孢藻在熱帶水體中可常年占據(jù)優(yōu)勢乃至形成持續(xù)性水華, 但在亞熱帶和溫帶地區(qū), 該藻僅能在夏季較高的溫度條件下形成優(yōu)勢種[11,12]。擬柱孢藻備受關(guān)注的另一原因是它能產(chǎn)生包括擬柱孢藻毒素(Cylindrospermopsin, CYN)在內(nèi)的多種毒素, CYN可抑制蛋白質(zhì)合成, 損傷 DNA, 具廣泛的細(xì)胞毒性, 嚴(yán)重危害人類健康[13]。
研究表明, 擬柱孢藻的表型可塑性強(qiáng), 對光照溫度等關(guān)鍵生長因子有較寬的生態(tài)輻[14]。該藻還具有一些獨特的生物學(xué)特性, 如良好的漂浮能力、耐受低光強(qiáng)、能固氮、可利用有機(jī)氮源、抵抗牧食者捕食等[1,15]。擬柱孢藻對磷有較高的親和力并且具有儲存磷的能力[16], 它的胞外堿性磷酸酶活性較高,能有效裂解并利用有機(jī)磷[17]。上述特征均賦予擬柱孢藻更強(qiáng)的競爭能力, 使其能在復(fù)雜多變的環(huán)境下生長并在合適的條件下暴發(fā)形成水華。
盡管目前對擬柱孢藻的適應(yīng)性特征已有較深入的認(rèn)識, 但研究發(fā)現(xiàn)擬柱孢藻擁有多種生態(tài)型, 每種生態(tài)型對水深、光強(qiáng)、營養(yǎng)物乃至有機(jī)物等環(huán)境因子的偏好都不一樣[18], 有必要根據(jù)地域探討有利于擬柱孢藻生長的關(guān)鍵因子。在我國的南亞熱帶地區(qū), 近幾年來擬柱孢藻逐漸成為水庫優(yōu)勢種并在部分水庫形成水華[19], 成為危害該地區(qū)水庫水質(zhì)安全的首要有害藻類。本文以從廣東省開平市鎮(zhèn)海水庫中分離的擬柱孢藻(Cylindrospermopsis raciborskiiN8)為材料, 在不同氮、磷濃度條件下進(jìn)行單因子和正交實驗, 研究擬柱孢藻對營養(yǎng)鹽的適應(yīng)性特征, 為揭示該地區(qū)擬柱孢藻水華發(fā)生機(jī)制提供基礎(chǔ)數(shù)據(jù)。
1 材料與方法
1.1 藻種分離、培養(yǎng)和鑒定
實驗用藻種C. raciborskii N8分離于廣東省開平市鎮(zhèn)海水庫(22°34′7′′N、112°32′58′′E)。在光學(xué)體視顯微鏡下, 用玻璃毛細(xì)管從水庫水樣中挑取單根藻絲, 無菌水洗滌5—6次后轉(zhuǎn)入3 mL BG-11培養(yǎng)基中[20], 置于28℃恒溫光照培養(yǎng)箱(FPG3萊福儀器)中培養(yǎng), 4周左右藻絲明顯生長后, 轉(zhuǎn)入250 mL錐形瓶中擴(kuò)大培養(yǎng), 光源采用白色熒光燈, 光照強(qiáng)度為 30 μE/(m2·s)(用LI-1400輻照度儀測定), 光暗周期為12h∶12h。
對所分離的擬柱孢藻依據(jù)其形態(tài)特征進(jìn)行種類鑒定。取適量生長良好的藻液制作臨時裝片, 在10×40倍顯微鏡(ZEISS AX10)下, 記錄50根藻絲的長度以及單個細(xì)胞和異形胞(不一定出現(xiàn))的形態(tài)、長度和寬度。同時采用針對16S rDNA設(shè)計的藍(lán)藻特異性引物對27F和809R進(jìn)行PCR擴(kuò)增[21], PCR產(chǎn)物送深圳華大基因科技服務(wù)有限公司進(jìn)行序列測定, 獲得的序列采用BLAST進(jìn)行相似性分析。
1.2 不同磷濃度的單因子實驗
以BG-11培養(yǎng)基為基礎(chǔ)(BG-11中硝態(tài)氮濃度為247 mg/mL, 磷濃度為5.43 mg/mL), 調(diào)整磷濃度,配置9個磷濃度梯度培養(yǎng)基: 0.02、0.04、0.08、0.16、0.32、0.64、1.28、2.56、5.12 mg/L, 每個處理組設(shè)置3個平行。取一定體積生長良好的擬柱孢藻液以5000 r/min離心15min, 棄去上清液, 用15 mg/L的NaHCO3溶液洗滌后再次離心, 重復(fù)3次, 然后用無菌水稀釋, 饑餓培養(yǎng)2d后接種。實驗采用單種批量培養(yǎng), 取相同體積藻液轉(zhuǎn)移至裝有30 mL不同磷濃度培養(yǎng)基的試管中, 起始接種葉綠素a濃度為(31.0± 2.0) μg/L。置于(25±1)℃恒溫光照培養(yǎng)箱中培養(yǎng), 光照強(qiáng)度為30 μE/(m2·s), 光暗周期為12h∶12h。
1.3 氮、磷濃度組合正交實驗
以BG-11培養(yǎng)基為基礎(chǔ), 設(shè)置低、中、高3個氮濃度(LN=0.5、MN=5.0、HN=50 mg/L)和低、中、高3個磷濃度(LP=0.04、MP=0.4、HP=4.0 mg/L), 共有(LNLP、LNMP、LNHP、MNLP、MNMP、MNHP、HNLP、HNMP、HNHP)9個處理組, 每個處理組設(shè)置3個平行。接種方法及實驗條件同1.2, 起始葉綠素a濃度為(22.7±0.5) μg/L。
1.4 擬柱孢藻生長曲線和比生長速率的計算
每天定時取樣, 遮光處理20min后用TD-700 Fluorometre熒光儀測定藻液葉綠素a的濃度。當(dāng)每組實驗每天的平均增長值低于5%時, 認(rèn)為該實驗組已達(dá)到最大現(xiàn)存量, 即可停止測定。比生長速率μ(/d)指在某一段時間間隔內(nèi)藻類的生長速率, 計算公式為:
μ = ln (x2/x1) / (t2-t1)
式中, x1為t1時的生物量, x2為t2時的生物量, 此處均為葉綠素a的濃度。而t1、t2分別為擬柱孢藻指數(shù)生長期開始和結(jié)束的時間。
1.5 動力學(xué)參數(shù)的計算[22]
根據(jù)Monod方程μ=μmaxC/(C+KS)轉(zhuǎn)變成
1/μ=KS/(C·μmax)+1/μmax
式中, μ為擬柱孢藻的比生長速率(/d), μmax為在飽和濃度中藻類最大比生長速率(/d), C為限制性底物的濃度(mg/L), KS為半飽和常數(shù)。以統(tǒng)計學(xué)的最小二乘法求出上述方程的斜率(KS/μmax)和截距(1/μmax)。截距的倒數(shù)為 μmax, 其與斜率的乘積就是半飽和常數(shù)KS。
1.6 數(shù)據(jù)分析
實驗數(shù)據(jù)采用Origin 9.0作圖, 采用 SPSS 16.0軟件進(jìn)行One-way ANOVA方差分析, 顯著性水平為P<0.05。
2 結(jié)果
2.1 擬柱孢藻N8的形態(tài)學(xué)特征
從圖1可知, 本實驗室分離到的擬柱孢藻是單根絲體, 偽空泡多, 藻絲筆直不彎曲, 藍(lán)綠色, 相鄰的細(xì)胞壁沒有縊縮, 未觀察到有黏質(zhì)的膠鞘。藻絲長度變化極大, 范圍為17—1220 μm。沒有異形胞的藻體其兩端細(xì)胞的末端稍尖或鈍圓。營養(yǎng)細(xì)胞呈圓柱形, 長3.3—14.0 μm, 寬2.3—4.3 μm。異形胞頂生, 位于藻絲一端或兩端, 偶有間生, 呈長圓形或卵圓形, 長 3.5—4.5 μm, 寬 2.3—2.8 μm(圖1b)。利用核苷酸序列比對檢索工具BLASTN進(jìn)行同源性分析表明, N8藻株的16S rDNA(Genbank序列號: KJ747394)與來自澳大利亞、德國、匈牙利、美國、巴西等國家的擬柱孢藻藻株序列相似性達(dá)99%以上。根據(jù)上述形態(tài)和分子特征, 將該藻定為Cylindrospermopsis raciborskii, 在本實驗室編號為N8。
2.2 不同磷濃度對擬柱孢藻生長的影響
磷濃度為0.02—5.12 mg/L時, 擬柱孢藻均有明顯的指數(shù)生長期, 各處理組葉綠素 a的峰值隨磷濃度的升高而增加, 不同磷濃度組的生長趨勢在第 6天后出現(xiàn)分歧。磷濃度為0.02—0.08 mg/L時, 擬柱孢藻生長趨勢類似, 第2至第6天為指數(shù)生長期, 在第6至第10天處于穩(wěn)定期, 在第10天開始進(jìn)入衰亡期, 3個濃度組的葉綠素 a峰值無顯著差異(P>0.05)。而磷濃度為0.16—0.64 mg/L時, 擬柱孢藻的指數(shù)生長期維持較長時間(第2至第11天), 各處理組間葉綠素 a濃度無顯著差異(P>0.05)。磷濃度為1.28—5.12 mg/L時, 擬柱孢藻在實驗期間一直處于生長期(圖2)。
擬柱孢藻的比生長速率隨著磷濃度的升高呈升高趨勢, 在0.02—0.64 mg/L磷濃度下, 比生長速率從0.174 /d上升至0.243 /d, 在0.64—5.12 mg/L范圍內(nèi)的比生長速率則無明顯差異。比生長速率在0.08 mg/L處有較大增長, 低于0.08 mg/L與高于 0.08 mg/L的各組間均存在顯著差異(P<0.05) (圖3)。結(jié)果表明, 磷濃度大于 0.08 mg/L能顯著促進(jìn)擬柱孢藻的生長, 當(dāng)磷濃度達(dá)到1.28 mg/L后, 磷濃度的增加對比生長速率無顯著促進(jìn)作用。擬柱孢藻對磷濃度有較寬的適應(yīng)范圍, 而最適生長范圍為 0.16—1.28 mg/L。

圖1 擬柱孢藻的形態(tài)照片F(xiàn)ig. 1 Cylindrospermopsis raciborskii N8
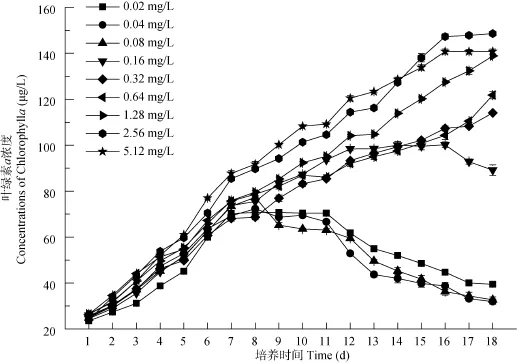
圖2 不同磷濃度下擬柱孢藻N8的生長曲線Fig. 2 Growth curves of C. raciborskii N8 under different phosphorus concentrations
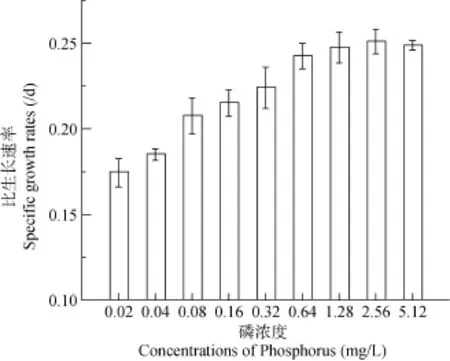
圖3 不同磷濃度下擬柱孢藻N8的比生長率Fig. 3 Specific growth rates of C. raciborskii N8 under different phosphorus concentrations
2.3 擬柱孢藻N8的營養(yǎng)生長動力學(xué)參數(shù)
半飽和常數(shù) KS表示生物物種對于營養(yǎng)物質(zhì)的親和性, 即在營養(yǎng)物質(zhì)濃度為KS時就能使種群增值率達(dá) μmax, 而親和性的強(qiáng)弱與其半飽和常數(shù)的值成反比[22]。實驗結(jié)果顯示: 擬柱孢藻N8以磷為限制性底物時的半飽和常數(shù)KSP=0.008 mg/L, μmax=0.239/d。
2.4 氮、磷濃度組合正交實驗
在三個低氮濃度組中, 擬柱孢藻基本停止生長,葉綠素 a濃度顯著低于其他處理組(P<0.05)。在MNLP和 HNLP組中, 擬柱孢藻的生長周期很短,指數(shù)生長時間僅3d, 葉綠素a峰值為45.3 μg/L。而MNMP和MNHP處理組的指數(shù)生長期為5d, 葉綠素 a峰值達(dá)到 73.5 μg/L。擬柱孢藻在 HNMP和HNHP下的生長最好, 指數(shù)生長期可達(dá)7d, 葉綠素a峰值可達(dá)(120±2) μg/L(圖4)。
擬柱孢藻的比生長速率的最大值均出現(xiàn)在 MN濃度組。在氮濃度較高(MN和HN)的情況下, 高磷濃度下的比生長速率顯著高于中、低磷濃度(P<0.05) (圖5)。
3 討論
3.1 擬柱孢藻N8的形態(tài)學(xué)特征
擬柱孢藻藻絲一般長50—250 μm, 營養(yǎng)細(xì)胞長5.0—10.0 μm, 寬 1.5—2.5 μm, 異形胞位于藻絲一端或兩端, 長3.5—10.0 μm, 寬1.5—2.5 μm[23—27]。而擬柱孢藻N8的藻絲更長(可達(dá)1220 μm), 營養(yǎng)細(xì)胞較寬, 異形胞則比較短, 最長僅為 4.5 μm, 可見分離自廣東省鎮(zhèn)海水庫的 N8藻株具較獨特的形態(tài)特征。擬柱孢藻的形態(tài)被認(rèn)為具高度的可塑性, 不同生境下的營養(yǎng)和捕食壓力差異均可導(dǎo)致其形態(tài)發(fā)生變化, 據(jù)Komárková等[28]的觀測, 即使在同一生境的種群內(nèi), 擬柱孢藻的形態(tài)變化也非常大; 以色列的一個湖泊中該藻藻絲末端則有6種不同的形態(tài)[25]。N8藻株形態(tài)獨特的原因也可能是長期實驗室培養(yǎng)所致, 有研究表明藻絲形態(tài)與培養(yǎng)溫度密切相關(guān)[23]。
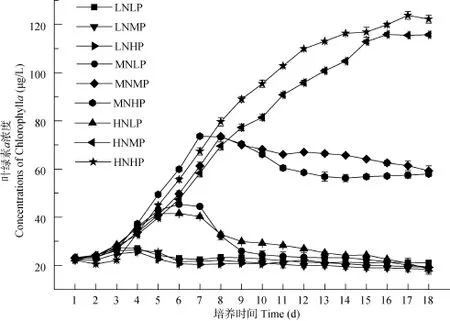
圖4 不同氮、磷濃度下擬柱孢藻N8的生長曲線Fig. 4 Growth curves of C. raciborskii N8 at different nitrogen and phosphorus concentrations

圖5 不同氮、磷濃度對擬柱孢藻N8比生長速率的影響Fig. 5 Effects of different nitrogen and phosphorus concentrations on specific growth rate of C. raciborskii N8
3.2 磷濃度對擬柱孢藻N8生長的影響
能快速吸收磷并適應(yīng)低磷環(huán)境的特性一直被認(rèn)為是擬柱孢藻能在全球范圍內(nèi)快速擴(kuò)張的關(guān)鍵因子之一[1,16], 在法國和澳大利亞的水體中, 擬柱孢藻的優(yōu)勢度常常在低磷條件下表現(xiàn)得更為明顯[7,29]。本研究結(jié)果顯示, 擬柱孢藻 N8在磷濃度低至0.02 mg/L時仍能維持較長時間的生長, 說明它對低磷有較強(qiáng)的適應(yīng)性。N8藻株的比生長速率在0.64—5.12 mg/L磷濃度下差異不顯著, 表明擬柱孢藻不但耐受低磷, 且能在較寬范圍的磷濃度下快速生長。Bonilla等[14]指出高度的表型可塑性和寬廣的環(huán)境適應(yīng)性有助于擬柱孢藻適應(yīng)不同的氣候條件, 因此廣東省擬柱孢藻的形態(tài)獨特性和對磷的適應(yīng)性是促進(jìn)其擴(kuò)張的重要因素。
本研究測定的擬柱孢藻 N8株以磷為限制性底物時的半飽和常數(shù)為 0.008 mg/L, 顯著高于來自巴拉頓湖的ACT9502藻株(KSP=0.0015–0.0025 mg/L),這種差異可能是藻株自身獨特性所致, 也可能與實驗條件相關(guān)。本文采用的是批量培養(yǎng), 而Istvánovics等[30]進(jìn)行的是恒化實驗。與銅綠微囊藻(KSP=0.009)、水華微囊藻(KSP=0.014)相比[31], 擬柱孢藻 N8株的KSP值更低, 因而會在磷濃度較低的水體中更具生長優(yōu)勢。地處南亞熱帶的廣東省, 土壤以含鐵的紅壤土為主, 由此導(dǎo)致多數(shù)水庫的入庫磷濃度很低,盡管近年來人類活動干擾的加劇, 水體富營養(yǎng)化有升高趨勢, 多數(shù)水庫依然為磷限制水體[19,32], 這可能是近年來廣東省水庫擬柱孢藻能取代微囊藻而成為水庫優(yōu)勢種的原因之一。
3.3 氮濃度對擬柱孢藻N8生長的影響
擬柱孢藻是一種固氮藍(lán)藻, 當(dāng)無機(jī)氮不足時,它可以進(jìn)行固氮以滿足生長要求[1]。但在本研究中, N8藻株在低氮濃度下(0.5 mg/L)的生長基本停止,且這種生長停滯與磷濃度的高低無關(guān), 擬柱孢藻的生物量僅能在高氮(50 mg/L)條件下達(dá)到較高水平,這表明氮對擬柱孢藻的生長起決定作用, 固氮并沒有在缺氮的情況下發(fā)揮作用。一般認(rèn)為固氮需要消耗大量能量, 是擬柱孢藻最少采用的氮攝取方式[29]。Spr?ber等[33]提出擬柱孢藻固氮系統(tǒng)的停止和啟動依賴于細(xì)胞自身的氮含量, 因此 N8藻株固氮酶的合成和異形胞的形成可能需要更低的氮濃度來啟動。磷元素對擬柱孢藻 N8株生長的限制明顯弱于氮, 在低磷條件下(0.04 mg/L), 氮濃度的升高(MN和 HN)可使擬柱孢藻維持一段時間的生長, 其生物量可在磷限制但氮充足(MPHN)的條件下達(dá)到較高水平, 這與Kenesi等[34]的研究報道一致。
4 結(jié)論
N8藻株從形態(tài)和分子角度上鑒定為擬柱孢藻,其絲體長度和細(xì)胞大小均比其他地區(qū)的擬柱孢藻大,具有較獨特的形態(tài)特征。擬柱孢藻 N8的最適生長磷濃度范圍為 0.16—5.12 mg/L, 半飽和常數(shù) KSP= 0.008 mg/L, 對磷的親和性較高, 在培養(yǎng)基中磷濃度低至 0.02 mg/L時仍能維持較長時間的生長, 說明該藻株耐受低磷并對磷有較強(qiáng)的適應(yīng)性。低氮顯著限制擬柱孢藻 N8的生長, 這種抑制不因磷濃度的升高而解除。相反, 氮濃度的升高可延長擬柱孢藻在磷限制性條件下的生長時間, 因此氮對擬柱孢藻N8的生長限制作用比磷更為顯著。
[1] Padisák J. Cylindrospermopsis raciborskii (Wo?oszynska) Seenayya and Subba Raju, an expanding highly adaptive cyanobacterium: worldwide distribution and review of its ecology [J]. Archiv Für Hydrobiologie Supplementband Monographische Beitrage, 1997, 107(4): 563—593
[2] Li R, Carmichael W W, Brittain S, et al. Isolation and identification of the cyanotoxincylindrospermopsin and deoxycylindrospermopsin from a Thailand strain of Cylindrospermopsis raciborskii (Cyanobacteria) [J]. Toxicon,2001, 39(7): 973—980
[3] Fastner J, Heinze R, Humpage A R, et al. Cylindrospermopsin occurrence in two German lakes and preliminary assessment of toxicity and toxin production of Cylindrospermopsis raciborskii (Cyanobacteria) isolates [J]. Toxicon, 2003, 42(3): 313—321
[4] Dokulil M T, Mayer J. Population dynamics and photosynthetic rates of a Cylindrospermopsis-Limnothrix association in a highly eutrophic urban lake, Alte Donau, Vienna, Austria [J]. Algological Studies, 1996, 83: 179—195
[5] Stirling D J, Quilliam M A. First report of the cyanobacterial toxin cylindrospermopsin in New Zealand [J]. Toxicon, 2001, 39(8): 1219—1222
[6] Kokocinski M, Stefaniak K, Mankiewicz-Boczek J, et al. The ecology of the invasive cyanobacterium Cylindrospermopsis raciborskii (Nostocales, Cyanophyta) in two hypereutrophic lakes dominated by Planktothrix agardhii (Oscillatoriales, Cyanophyta) [J]. European Journal of Phycology, 2010, 45(4): 365—374
[7] Briand J F, Robillot C, Quiblier-Llobéras C, et al. Environmental context of Cylindrospermopsis raciborskii (Cyanobacteria) blooms in a shallow pond in France [J]. Water Research, 2002, 36(13): 3183—3192
[8] Hamilton P B, Ley L M, Dean S, et al. The occurrence of the cyanobacterium Cylindrospermopsis raciborskii in Constance Lake: an exotic cyanoprokaryote new to Canada [J]. Phycologia, 2005, 44(1): 17—25
[9] Stuken A, Rucker J, Endrulat T, et al. Distribution of three alien cyanobacterial species (Nostocales) in northeast Germany: Cylindrospermopsis raciborskii, Anabaena bergii and Aphanizomenon aphanizomenoides [J]. Phycologia, 2006, 45(6): 696—703
[10] Briand J F, Leboulanger C, Humbert J F. Cylindrospermopsis raciborskii (cyanobacteria) invasion at midlatitudes: selection, wide physiological tolerance, or global warming [J]? Journal of Phycology, 2004, 40(2): 231—238
[11] Bouvy M, Falc?o D, Marinho M, et al. Occurrence of Cylindrospermopsis (Cyanobacteria) in 39 Brazilian tropical reservoirs during the 1998 drought [J]. Aquatic Microbial Ecology, 2000, 23(1): 13—27
[12] Wiedner C, Rücker J, Brüggemann R, et al. Climate change affects timing and size of populations of an invasive cyanobacterium in temperate regions [J]. Oecologia, 2007, 152(3): 473—484
[13] Moreira C, Azevedo J, Antunes A, et al. Cylindrospermopsin: occurrence, methods of detection and toxicology [J]. Journal of Applied Microbiology, 2013, 114(3): 605—620
[14] Bonilla S, Aubriot L, Soares MCS, et al. What drives the distribution of the bloom-forming cyanobacteria Planktothrixagardhii and Cylindrospermopsis raciborskii [J]? FEMS Microbiology Ecology, 2012, 79(3): 594—607
[15] See J H, Bronk D A, Lewitus A J. Uptake of Spartinaderived humic nitrogen by estuarine phytoplankton in axenic and nonaxenic culture [J]. Limnology and Oceanography, 2006, 51(5): 2290—2299
[16] Istvánovics V, Shafik H M, Présing M, et al. Growth and phosphate uptake kinetics of the cyanobacterium, Cylindrospermopsis raciborskii (cyanophyceae) in throughflow cultures [J]. Freshwater Biology, 2000, 43(2): 257—275
[17] Wu Z, Shi J, Li R. Comparative studies on photosynthesis and phosphate metabolism of Cylindrospermopsis raciborskii with Microcystis aeruginosa and Aphanizomenon flos-aquae [J]. Harmful Algae, 2009, 8(6): 910—915
[18] Piccini C, Aubriot L, Fabre A, et al. Genetic and ecophysiological differences of South American Cylindrospermopsis raciborskii isolates support the hypothesis of multiple ecotypes [J]. Harmful Algae, 2011, 10(6): 644—653
[19] Jiang Q M, Hou W, Gu J G, et al. Nutritional status and population characteristics of Cyanobacteria in small and medium sized reservoirs in Guangzhou, southern China [J]. Ecology and Environment Sciences, 2010, 19(10): 2461—2467 [江啟明, 侯偉, 顧繼光, 等. 廣州市典型中小型水庫營養(yǎng)狀態(tài)與藍(lán)藻種群特征. 生態(tài)環(huán)境學(xué)報, 2010, 19(10): 2461—2467]
[20] Rippka R, Deruelles J B, Waterbury M, et al. Genetics assignments, strain stories and properties of pure cultures of cyanobacteria [J]. Journal of General and Applied Microbiology, 1979, 11: 1–61
[21] Vollmer W, Joris B, Charlier P, et al. Bacterial peptidoglycan (murein) hydrolases [J]. FEMS Microbiology Reviews, 2008, 32(2): 259—286
[22] Xu H, Yang L Z, Mao H, et al. Dynamic studies on the effect of phosphorus on the growthof Microcystis aeruginosa and Scendesmus obliquus [J]. Ecology and Environment, 2006, 15(5): 921—924 [許海, 楊林章, 茅華, 等. 銅綠微囊藻、斜生柵藻生長的磷營養(yǎng)動力學(xué)特征. 生態(tài)環(huán)境, 2006, 15(5): 921—924]
[23] Soares M C S, Lürling M, Huszar V L M. Growth and temperature-related phenotypic plasticity in the cyanobacterium Cylindrospermopsis raciborskii [J]. Phycological Research, 2013, 61(1): 61—67
[24] Saker M L, Neilan B A. Varied diazotrophies, morphologies, and toxicities of genetically similar isolates of Cylindrospermopsis raciborskii (Nostocales, Cyanophyceae) from Northern Australia [J]. Applied and Environmental Microbiology, 2001, 67(4): 1839—1845
[25] Alster A, Kaplan-Levy R N, Sukenik A, et al. Morphology and phylogeny of a non-toxic invasive Cylindrospermopsis raciborskii from a Mediterranean Lake [J]. Hydrobiologia, 2010, 639(1): 115—128
[26] Padisák J. Occurrence of Anabaenopsis raciborskii Wolosz. in the pond Tómalom near Sopron, Hungary [J]. Acta Botanica Hungarica, 1991, 36: 163—165
[27] Komárek J, Komárková J. Phenotype diversity of the cyanoprokaryotic genus Cylindrospermopsis (Nostocales);review 2002 [J]. Czech Phycology, 2003, 3: 1—30
[28] Komárková J, Laudares-Silva R, Senna P A C. Extreme morphology of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) in the Lagoa do Peri, a freshwater costal lagoon, Santa Catarina, Brazil [J]. Algological Studies, 1999, 94: 207—222
[29] Burford M A, Mcneale K L, Mckenzie-smith F J. The role of nitrogen in promoting the toxic cyanophyte Cylindrospermopsis raciborskii in a subtropical water reservoir [J]. Freshwater Biology, 2006, 51(11): 2143—2153
[30] Istvánovics V, Kovács A, V?r?s L, et al. Phosphorus cycling in a large, reconstructed wetland, the lower Kis-Balaton Reservoir (Hungary) [J]. Verh Internat Verein Limnol, 1997, 26: 323—329
[31] Xie L J,Miu H F, Yan Q, et al. Effects of phophorus limitation to the growth of Microcystis aeruginosa and Microcystis flos-aquae [J]. Shanghai Environmental Sciences, 2012, 30(3): 124—129 [謝利娟, 繆恒鋒, 嚴(yán)群, 等. 磷限制對銅綠和水華微囊藻生長情況的影響. 上海環(huán)境科學(xué), 2011, 30(3): 124—128]
[32] Hu R, Lin Q Q, Duan S S, et al. Distribution of chlorophyll-a and phosphorus in subtropical reservoirs, South China [J]. Ecologic Science, 2002, 21(4): 289—295 [胡韌, 林秋奇,段瞬山, 等. 熱帶亞熱帶水庫浮游植物葉綠素a與磷分布的特征. 生態(tài)科學(xué), 2002, 21(4): 289—295]
[33] Spr?ber P, Shafik H M, Présing M, et al. Nitrogen uptake and fixation in the cyanobacterium Cylindrospermopsis raciborskii under different nitrogen conditions [J]. Hydrobiologia, 2003, 506(1—3): 169—174
[34] Kenesi G, Shafik H M, Kovács A W, et al. Effect of nitrogen forms on growth, cell composition and N2fixation of Cylindrospermopsis raciborskii in phosphoruslimited chemostat cultures [J]. Hydrobiologia, 2009, 623(1): 191—202
THE EFFECTS OF PHOSPHORUS AND NITROGEN ON THE GROWTH OF CYLINDROSPERMOPSIS RACIBORSKII N8 ISOLATED FROM THE ZHENHAI RESERVOIR
DAI Jing-Jun, PENG Liang, YU Ting and LEI La-Mei
(Department of Ecology and Key Laboratory of Eutrophication and Red Tide Prevention of Guangdong Higher Education Institutes, Jinan University, Guangzhou 510632, China)
In this study we examined the effects of nitrate and phosphate on the growth of Cylindrospermopsis raciborskii N8 that was isolated from the Zhenhai Reservoir in Guangdong Province. We used a crossed factorial design for three phosphorus levels and three nitrogen levels, and thus generated nine different growth conditions for C. raciborskii. Our results showed that C. raciborskii could adapt to a wide range of phosphorus concentrations (from 0.02 mg/mL to 5.12 mg/mL) and the optimal concentrations for the growth was between 0.16 mg/mL and 5.12 mg/mL. The exponential stage and biomass of C. raciborskii could be significantly enhanced by the increase in the phosphorus level. The kinetic analysis revealed a low KSPvalue of C. raciborskii N8. This suggested that C. raciborskii N8 could have high affinity for phosphorus and readily become the dominant species in phosphorus-lacking conditions. The results of the nitrogen and phosphorus orthogonal experiments demonstrated that when the nitrogen level was low (0.5 mg/mL), the growth of C. raciborskii N8 was markedly inhibited regardless of the phosphorus level. Moreover, at low phosphorus concentration (0.04 mg/mL) the proliferation of C. raciborskii N8 could be boosted by the increase in nitrogen concentration. It was also found that the growth of C. raciborskii N8 could be significantly improved by HNMP and HNHP treatment. The highlights of our findings were that C. raciborskii in Guangdong reservoirs was highly tolerant to low phosphorus conditions, and nitrogen could be a key factor in its growth.
the Zhenhai Reservoir; Cylindrospermopsis raciborskii; Nitrogen and Phosphorus; Growth characteristics; Southern subtropics
Q142
A
1000-3207(2015)03-0533-07
10.7541/2015.70
2014-05-05;
2014-08-14
暨南大學(xué)“國家級大學(xué)生創(chuàng)新創(chuàng)業(yè)訓(xùn)練計劃”項目(1210559020); 廣東省水利科技創(chuàng)新項目(2011-02)資助
戴景峻(1991—), 男, 廣東大埔人; 本科生; 研究方向為藻類生物學(xué)。E-mail: 343996767@qq.com
雷臘梅, 女, 副教授; E-mail: tleilam@jnu.edu.cn
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(shù)(2020年3期)2020-12-06 07:32:54
現(xiàn)代裝飾(2020年11期)2020-11-27 01:47:48
中學(xué)生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
