棉鈴蟲成蟲對兩種關鍵花香氣味的聯系性學習
2015-02-07 05:49:07李為爭李慧玲李洋洋郭線茹原國輝
生態學報 2015年11期
李為爭, 王 瓊, 李慧玲, 王 玨, 李洋洋, 郭線茹, 原國輝
河南農業大學植物保護學院, 鄭州 450002
棉鈴蟲成蟲對兩種關鍵花香氣味的聯系性學習
李為爭, 王 瓊, 李慧玲, 王 玨, 李洋洋, 郭線茹, 原國輝*
河南農業大學植物保護學院, 鄭州 450002
為了明確棉鈴蟲對苯乙醛和乙酸苯甲酯兩種關鍵花香氣味的聯系性學習行為,在室內分別測定了不同學習訓練的棉鈴蟲雌雄成蟲對兩種關鍵花香氣味的選擇偏好性反應和雄成蟲的觸角電位反應。偏好性測定結果表明,無花香氣味接觸經歷的棉鈴蟲對苯乙醛和乙酸苯甲酯的選擇頻次均無顯著差異,且單純的花香氣味接觸經歷也不能誘導棉鈴蟲氣味偏好性的顯著改變,唯有花香氣味接觸經歷伴隨蔗糖溶液進行強化訓練時,才能誘導氣味偏好性的顯著性改變。從對兩種花香氣味的聯系性學習看,棉鈴蟲更容易將苯乙醛與蔗糖溶液進行聯系性學習,而不容易將乙酸苯甲酯與蔗糖溶液進行聯系性學習。從性別差異看,雌蟲比雄蟲具有更強的將花香氣味與蔗糖溶液進行聯系性學習的能力。觸角電位測定結果表明,不同學習訓練的棉鈴蟲雄成蟲對兩種花香氣味和綠葉氣味順- 3-己烯- 1-醇的EAG反應值之間不存在顯著性差異。總之,花香氣味伴隨食物資源的聯系性學習經歷,能夠明顯提高棉鈴蟲成蟲對花香氣味的選擇偏好,可以此為基礎進一步優化花香引誘劑的配方設計和應用技術。
棉鈴蟲; 花香氣味; 聯系性學習; 選擇偏好性
聯系性學習(Associative learning)是指通過經歷在兩種刺激間或一種刺激與一種反應間建立的一種聯系。這類學習行為在蜜蜂搜尋花蜜和寄生蜂搜索寄主的過程中較為常見[1- 2],且已發現一些植食性昆蟲中也存在聯系性學習[3]。如大帛斑蝶Idealeoconoe、斐豹蛺蝶Argyreushyperbius、菜粉蝶Pierisrapae和紅灰蝶Lycaenaphlaeas可以將花的顏色與花蜜進行聯系性學習[4],沙漠蝗Schistocercagregaria可以將嗅覺刺激與食物獎勵進行聯系性學習[5],煙芽夜蛾Heliothisvirescens可以將植物氣味與寄主適合度進行聯系性學習[6]。由于植食性昆蟲的學習行為有可能對害蟲防治方法的效果產生積極或消極影響,因此,深入研究植食性昆蟲的學習行為將有助于發展和完善害蟲綜合治理的策略和方法[7]。
棉鈴蟲Helicoverpaarmigera是重要的多食性農業害蟲,幼蟲可取食為害30余科200多種植物,成蟲則具有取食多種植物花蜜作為補充營養的習性[8]。近年來,為了發展棉鈴蟲等蛾類害蟲的綠色防控,研制開發花香引誘劑大量誘殺成蟲受到關注[9],這類引誘劑多為單一或2—3種活性成分的混配制劑[10],其中苯乙醛對鱗翅目成蟲具有廣譜引誘活性[11],乙酸苯甲酯對鈴夜蛾屬Helicoverpa、實夜蛾屬Heliothis和金翅夜蛾屬Plusia的許多夜蛾具有特殊的引誘力[12- 13],這兩種揮發物大量存在于蛾類蜜源寄主和產卵寄主中,是激發棉鈴蟲等蛾類成蟲趨向行為反應的關鍵花香氣味[14]。田間誘捕結果表明,花香引誘劑不僅可以誘捕到多種蛾類,而且可以同時引誘雌、雄成蟲,具有明顯的廣譜誘蟲特點,展現出良好的開發應用前景[15]。然而,已有的研究表明,許多蛾類成蟲也具有聯系性學習行為,如未交配的棉鈴蟲兩性成蟲均偏好在有過取食經歷的蜜源寄主上覓食[16- 17],有過訪問蜜源植物或取食蔗糖經歷的棉鈴蟲成蟲對花香氣味表現出更強的選擇偏好[18]。一般認為,這種聯系性學習行為有助于棉鈴蟲高效識別有報酬的花[19],但是否會影響花香引誘劑的誘蛾效果,顯然是一個值得深入探討的問題。為此,本文以苯乙醛和乙酸苯甲酯兩種關鍵花香氣味物質為材料,研究了棉鈴蟲成蟲對它們的聯系性學習行為,期望為基于花香氣味物質的棉鈴蟲引誘劑篩選和應用提供科學依據。
1 材料與方法
1.1 供試昆蟲
供試棉鈴蟲蛹購于河南濟源白云實業有限公司,成蟲羽化后飼喂5%蔗糖水。后代幼蟲在實驗室人工氣候箱內用麥胚基人工飼料飼養,飼料主要成分為麥麩、酵母粉和干酪素,另添加維生素C、維生素復合粉、山梨酸和瓊脂粉等。人工氣候箱飼養條件為日溫(28±2)℃,夜溫(26±2)℃,相對濕度(50±10)%,光周期為16 L∶8 D。為保證幼蟲取食經歷和遺傳背景相對一致,選取連續飼養5代后的實驗種群用于測定。
每次測定前收集棉鈴蟲蛹,區分雌雄后分別放入相互隔離的不同人工氣候箱中。每日8:00和17:00各檢查一次羽化情況,及時將羽化的雌雄成蟲分別放入不同的L×W×H=20 cm × 20 cm× 20 cm籠中,飼喂5%蔗糖溶液。當日羽化的成蟲記為0日齡,次日取1日齡的未交配雌雄成蟲按試驗設計分別進行學習訓練,學習訓練24 h后取2日齡成蟲進行氣味選擇偏好性和觸角電位反應測定。
1.2 供試試劑
苯乙醛(Phenylacetaldehyde,PAA)和順- 3-己烯- 1-醇購于Sigma-Aldrich Co Ltd.,純度分別為98%和99%;乙酸苯甲酯(Benzyl acetate,BA)購于中國醫藥集團(上海)化學試劑有限公司,純度為95%;瓊脂粉購于Amresco公司,純度為99%;石蠟油購于天津科密歐公司,分析純。
將試劑按測定要求制備成不同的樣品。制備氣味物質緩釋樣品時,用長7.0 cm、直徑2.0 cm的平底指形管作為容器,每個管中首先注入400 μL苯乙醛或乙酸苯甲酯,然后迅速注入60 ℃左右的0.2%瓊脂膠20 mL,在室溫下冷卻后備用;另制備不添加氣味物質的20 mL瓊脂膠指形管樣品。預試驗表明,在實驗室內瓊脂膠包結后的氣味物質50 d可揮發完畢,氣味釋放速率約為8 μL/d。制備電生理測定樣品時,將苯乙醛、乙酸苯甲酯和順- 3-己烯- 1-醇分別用石蠟油稀釋100倍作為待測樣品。
1.3 學習訓練
1.3.1 學習室的準備
學習室為L×W×H= 30 cm × 30 cm × 20 cm的潔凈玻璃室。學習訓練前根據測定設計取上述指形管樣品1個,開口向上放在1個高2 cm、直徑6 cm的培養皿正中央,圍繞指形管在培養皿中填充脫脂棉與皿口齊平,然后將培養皿放在學習室底部中央。每次學習訓練后,更換培養皿,并用脫脂棉蘸取無水乙醇均勻擦拭學習室內壁,然后用蒸餾水沖洗干凈,在太陽光下晾曬干,以備下次使用。
1.3.2 學習訓練設置
共設置3組學習訓練,雌雄成蟲分別隔離進行,共10個處理。第1組為無花香氣味經歷組,簡稱無經歷組(Na?ve group):取不含氣味物質的瓊脂膠指形管樣品,開口向上放在培養皿中間,填充脫脂棉后均勻滴加5%蔗糖溶液使其完全浸潤。第2組為花香氣味經歷組,通稱暴露組(Exposure group):取含有氣味物質的瓊脂膠指形管樣品,開口向上放在培養皿中間,填充脫脂棉后均勻滴加蒸餾水使其完全浸潤。第3組為氣味與食物關聯經歷組,通稱條件化組(Conditioning group):取含有氣味物質的瓊脂膠指形管樣品,開口向上放在培養皿中間,填充脫脂棉后均勻滴加5%蔗糖溶液使其完全浸潤。設置完成后進行學習訓練,于19:00—22:00取1日齡的未交配雌雄成蟲分別放入不同學習室,密閉后移入人工氣候箱中,學習訓練24 h后取出用于測定。
1.4 氣味偏好性測定
1.4.1 測定裝置
測定在L×W×H= 100 cm × 100 cm × 100 cm玻璃籠罩內進行,籠罩頂板可拆卸,頂板正中央有一個直徑2.0 cm的圓孔供釋放試蟲,籠罩外上方中央懸掛5 W紅色燈泡作為觀察光源。籠罩內底部4個角落各放置市售小型向日葵花模型一朵,花盤直徑3.0 cm,由內至外共3層花瓣,第1層花瓣長2.0 cm,第2和第3層花瓣長3.5 cm。另取苯乙醛和乙酸苯甲酯氣味樣品各2個,交叉配對懸掛在花盤上方,指形管開口向下,開口部分距離花盤約3.0 cm。每次測定完成后將玻璃籠罩水平旋轉90 °,每測定10次后更換新的向日葵花模型。
1.4.2 測定方法
測定在19: 30—21: 30成蟲活動取食高峰期進行,室溫(26±2)℃。每次測定取不同學習經歷的成蟲1頭放入玻璃籠罩內,觀察其在靜息空氣中的飛行和降落情況,如果試蟲降落在某個向日葵花上并停留10 s以上,則視為做出了選擇;如果試蟲無規則飛行達10 min,則視為未作出選擇。每頭成蟲僅測試1次,共10個處理,每個處理測定40頭成蟲。
1.5 觸角電位測定
1.5.1 測定裝置
觸角電位儀購置于德國Syntech公司,由前置放大器(Syntech AC/DC UN-06)、刺激氣流控制器(Syntech CS- 55)、數據采集/放大器(Syntech IDAC- 4)和微動操作儀(Syntech MP- 15)組成,數據采集軟件為EAG 2000。兩根Ag/AgCl電極分別作為參比電極和記錄電極,插入2根玻璃毛細管中,管內注滿Kaissling電極緩沖液。緩沖液配制方法為:取6.4 mmol KCl,20.0 mmol KH2PO4,12.0 mmol MgCl2,1.0 mmol CaCl2,354 mmol葡萄糖,12.0 mmol NaCl和9.6 mmol KOH混合均勻,然后添加蒸餾水定容至1 L。
1.5.2 測定方法
考慮到花香氣味對雌蟲具有引誘取食和引誘產卵雙重作用,而對雄蟲只有取食引誘作用[20],因此,觸角電位反應只測定雄蟲。共設苯乙醛和乙酸苯甲酯2個花香氣味處理,以順- 3-己烯- 1-醇為參照,以石蠟油為對照,分別測定不同學習經歷棉鈴蟲雄蟲的觸角電位反應。測定前先在雙目體視顯微鏡下將觸角從基部切下并切去端部幾節,立即將離體觸角的端部和基部分別與記錄電極和參比電極連接。然后調試儀器,待基線平穩后進行測定。測定時取稀釋后的待測樣品0.2 μL滴加在6 cm × 0.5 cm濾紙條上放入進樣管中,管口距離觸角1 cm。刺激物和濕潤的潔凈氣流通過兩個獨立管道吹送到離體觸角上,并用刺激氣流控制器將流速控制在20 mL/s。每次連續刺激時間為0.02 s,刺激間隔時間為60 s,反應電壓信號通過數據采集/放大器記錄并輸出到電腦中保持。4個待測樣品采用隨機化順序進行刺激測定,完成一輪刺激后,仍隨機抽取進行下一輪刺激,每根觸角共測定6輪,每種學習經歷雄成蟲重復測定4根觸角。
1.6 統計分析
采用SPSS 19.0統計軟件進行分析。其中氣味偏好性測定結果采用2檢驗方法,比較不同處理間的偏好性差異。觸角電位測定結果采用雙因素方差分析(Two-way ANOVA),比較不同處理間的觸角電位反應差異。
2 結果與分析
2.1 棉鈴蟲對兩種花香氣味的聯系性學習
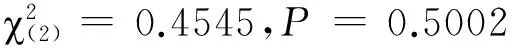
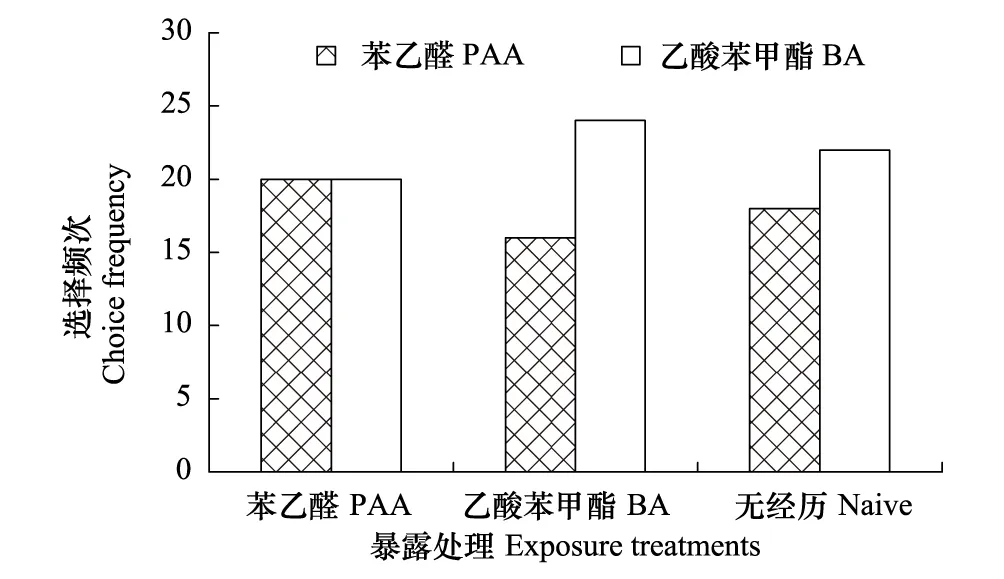
圖1 氣味暴露處理對棉鈴蟲雌成蟲氣味偏好性的影響
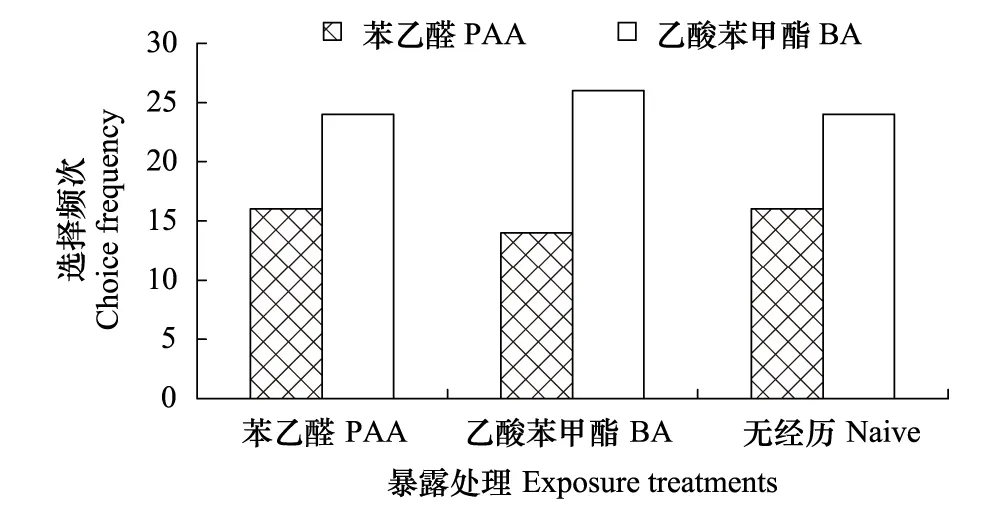
圖2 氣味暴露處理對棉鈴蟲雄成蟲氣味偏好性的影響
2.2 棉鈴蟲對花香氣味與食物的聯系性學習
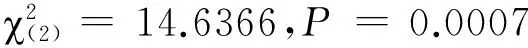

圖3 條件化處理對棉鈴蟲雌成蟲氣味偏好性的影響
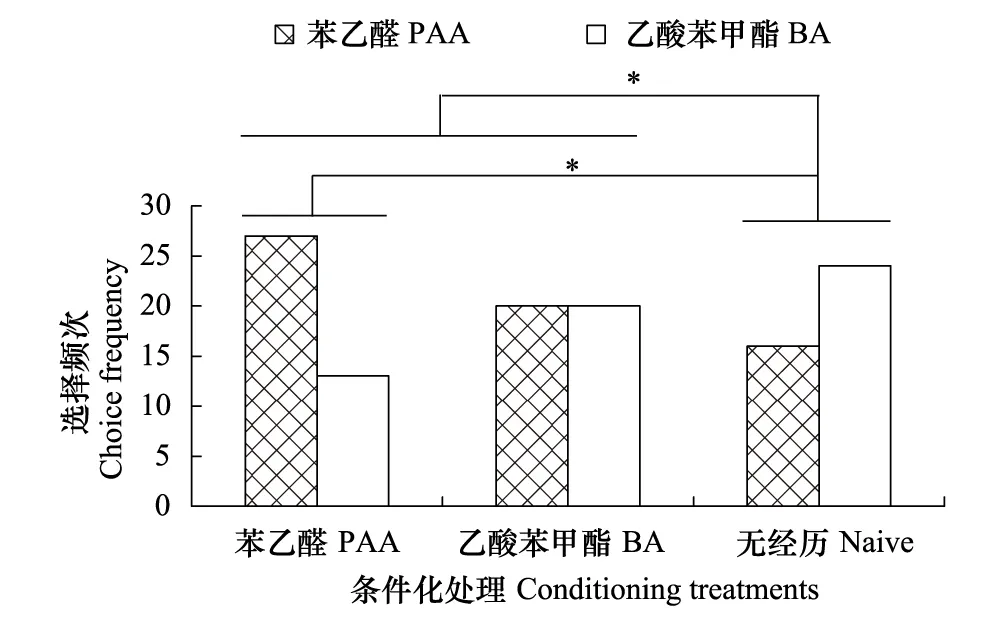
圖4 條件化處理對棉鈴蟲雄成蟲氣味偏好性的影響
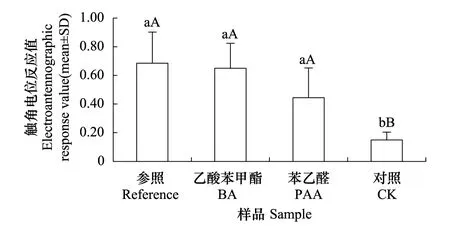
圖5 棉鈴蟲雄蟲對不同氣味樣品的觸角電位反應
2.3 棉鈴蟲雄蟲對兩種花香氣味的觸角電位反應
測定不同學習經歷的棉鈴蟲雄蟲對不同花香氣味的觸角電位(EAG)反應,方差分析結果表明,不同處理樣品觸發的EAG反應值之間存在極顯著差異(F= 21.0456,F(0.01,3,36)= 4.3771),而無經歷組、暴露組和條件化組的EAG反應值之間則不存在顯著性差異(F= 0.9115,F(0.05,2,36)= 3.2594,P= 0.4110),兩者也不存在顯著交互作用(F= 0.3029,F(0.05,2,36)= 2.3638,P= 0.9313)。分別計算不同處理樣品激發的觸角電位反應值(圖5),多重比較結果表明,苯乙醛和乙酸苯甲酯與參照化合物順- 3-己烯- 1-醇之間不存在顯著性差異,但三者均與對照石蠟油之間存在顯著性差異。綜合考慮2.2棉鈴蟲雄蟲可以將苯乙醛與食物資源進行聯系性學習的研究結果,可以認為雄蟲對苯乙醛的聯系性學習是發生在中樞神經系統水平上的,而不是發生在嗅覺感受器水平上的。
3 討論
在長期的協同進化過程中,蛾類昆蟲與顯花植物形成了密切的關系,植物花器釋放的花香氣味可以吸引多種蛾類成蟲,而絕大多數蛾類成蟲可以取食多種植物的花蜜[21]。如棉鈴蟲的蜜源寄主就非常廣泛,包括胡蘿卜、芹菜、大蔥、洋蔥、向日葵、茼蒿、蛇目菊、玉米花絲等[22]。由于這些蜜源植物的開花期并不完全一致,不同世代的棉鈴蟲面臨的蜜源寄主格局也會不同,這就需要一種機制來更好地適應多變的蜜源寄主環境。一般認為,本能性偏好主要是幫助新羽化的成蟲識別蜜源寄主存在的環境,而學習行為可以強化成蟲對食物伴隨的嗅覺刺激的反應[18]。因此,與遺傳決定的本能性偏好相比,聯系性學習可能對多食性棉鈴蟲的蜜源寄主選擇影響更大。
本研究表明,無花香氣味接觸經歷的棉鈴蟲對苯乙醛和乙酸苯甲酯兩種關鍵花香氣味的選擇頻次均無顯著差異,且單純的花香氣味接觸經歷也不能誘導棉鈴蟲氣味偏好性的顯著改變,唯有花香氣味接觸經歷伴隨蔗糖溶液進行強化時,才能誘導氣味偏好性的顯著改變,其中棉鈴蟲更容易將苯乙醛與蔗糖溶液進行聯系性學習,而不容易將乙酸苯甲酯與蔗糖溶液進行聯系性學習,這種差別的原因可能與兩種氣味物質在植物中的存在狀態有關。植物花蜜中最重要的營養物質是蔗糖、葡萄糖和果糖[23],而苯乙醛是莽草酸代謝途徑的產物,在花中主要存在形式是糖苷,當植物開花時糖苷水解被大量釋放出來[24],故苯乙醛能更可靠地指示花中可溶性糖類的存在,這對于成蟲為虹吸式口器、具有補充營養習性的棉鈴蟲來說尤為重要。而乙酸苯甲酯呈典型的茉莉花香氣味,可能是莽草酸代謝途徑產生的苯甲醇和乙酰輔酶A的反應產物[25],其釋放節律與植物花中可溶性糖的形成并不同步,其指示可溶性糖類存在的作用相對較弱。此外,從雌雄成蟲聯系性學習的差別看,雌蟲擔負著繁殖后代的重任,取食補充營養可以延長壽命和提高產卵量[26],因此,雌蟲的聯系性學習行為比雄蟲更加明顯。
研究植食性昆蟲對花香氣味的學習行為可以為活性化合物的篩選和引誘劑的應用提供有益的啟示。已有的研究表明,取食可溶性糖類是許多蛾類成蟲趨向花的最終目的[14],本文選用的兩種花香氣味物質能夠引誘多種蛾類成蟲,但只有苯乙醛與食物資源伴隨訓練時,棉鈴蟲才有聯系性學習行為。因此,在大量篩選花香類誘蛾活性成分時,可以從生物合成和次生代謝的角度,首先分析哪些揮發物成分的釋放與可溶性糖類的大量產生緊密伴隨,從而排除一些關聯性不強的成分,以便縮小待測揮發物的范圍。在應用花香引誘劑誘捕蛾類害蟲時,考慮到許多蛾類飛翔能力較強,與優勢蜜源植物接觸的概率大,學習行為發生的機會多,應首先明確特定時空區域的優勢蜜源植物及其分布情況,并根據不同世代所面臨的蜜源寄主格局,適時調整引誘劑的配方,以克服背景氣味的干擾,增強引誘劑的誘捕率。當然,影響棉鈴蟲等蛾類聯系性學習行為的環境因素可能非常復雜,深入研究兩種關聯刺激的濃度、刺激順序、刺激時間差、空間距離、伴隨性出現的概率、學習訓練次數、單次學習持續期等對學習效果的影響也非常必要。此外,糖醋液是一種常用的食物源引誘劑,可用于小地老虎、黏蟲、甜菜夜蛾、甘藍夜蛾、斜紋夜蛾、銀紋夜蛾等蛾類成蟲以及金龜甲、實蠅的大量誘捕防治,但對棉鈴蟲的誘捕效果并不理想,若將花香引誘劑誘捕盆中的清水變為糖醋液,是否可以發揮相輔相成的作用,進而提高對棉鈴蟲的誘捕效果,也有待田間試驗予以證實。
[1] Dukas R. Evolutionary biology of insect learning. Annual Review of Entomology, 2008, 53(1): 145- 160.
[2] 王國紅, 劉勇, 戈峰, 吳開拓. 粉蝶盤絨繭蜂中國和荷蘭種群學習行為及EAG反應的比較. 生態學報, 2012, 32(2): 351- 360.
[3] 李月紅, 劉樹生. 植食性昆蟲的學習行為. 昆蟲學報, 2004, 47(1): 106- 116.
[4] Kandori I, Yamaki T, Okuyama S, Sakamoto N, Yokoi T. Interspecific and intersexual learning rate differences in four butterfly species. The Journal of Experimental Biology, 2009, 212(23): 3810- 3816.
[5] Sim?es P, Ott S R, Niven J E. Associative olfactory learning in the desert locust,Schistocercagregaria. The Journal of Experimental Biology, 2011, 214(15): 2495- 2503.
[6] Skiri H T, Stranden M, Sandoz J C, Menzel R, Mustaparta H. Associative learning of plant odorants activating the same or different receptor neurones in the mothHeliothisvirescens. The Journal of Experimental Biology, 2005, 208(4): 787- 796.
[7] 呂建華, 李月紅, 劉樹生. 植食性昆蟲學習行為與害蟲治理的關系. 昆蟲知識, 2008, 45(4): 663- 667.
[8] 仵均祥, 李照會, 原國輝. 農業昆蟲學 (第二版). 北京: 中國農業出版社, 2011: 214- 218.
[9] Gregg P C, del Socorro A P, Henderson G S. Development of a synthetic plant volatile-based attracticide for female noctuid moths. II. Bioassays of synthetic plant volatiles as attraction for adults of cotton bollworm,Helicoverpaarmigera(Hübner) (Lepidoptera: Noctuidae). Australian Journal of Entomology, 2010, 49(1): 21- 30
[10] Meagher R L, Landolt P J. Binary floral lure attractive to velvetbean caterpillar adults (Lepidoptera: noctuidae). Florida Entomologist, 2010, 93(1): 73- 79.
[11] Landolt P J, Adams T, Zack R S. Field response of alfalfa looper and cabbage looper moths (Lepidoptera: Noctuidae, Plusiinae) to single and binary blends of floral odorants. Environmental Entomology, 2006, 35(2): 276- 281.
[12] Meagher R L Jr. Trapping noctuid moths with synthetic floral volatile lures. Entomologia Experimentalis et Applicata, 2002, 103(3): 219- 226.
[13] Landolt P J, Smithhisle C L. Characterization of the floral scents of oregongrape: possible feeding attractants for moths. Northwest Science, 2003, 77(1): 81- 86.
[14] Gregg P C, Del Socorro A P, Henderson G S. Development of a synthetic plant volatile-based attracticide for female noctuid moths. II. Bioassays of synthetic plant volatile as attractant for adults of cotton bollworm,Helicoverpaarmigera(Hübner) (Lepidoptera: Noctuidae). Australian Journal of Entomology, 2010, 49(1): 21- 30.
[15] Tóth M, Szarukán I, Dorogi B, Gulyás A, Nagy P, Rozgonyi Z. Male and female noctuid moths attracted to synthetic lures in Europe. Journal of Chemical Ecology, 2010, 36(6): 592- 598.
[16] Hartlieb E, Rembold H. Behavioral response of femaleHelicoverpa(Heliothis)armigeraHB. (Lepidoptera: Noctuidae) moths to synthetic pigeonpea (CajanuscajanL.) kairomone. Journal of Chemical Ecology, 1996, 22(4): 821- 837.
[17] Cunningham J P, West S A, Wright D J. Learning in the nectar foraging behaviour ofHelicoverpaarmigera. Ecological Entomology, 1998, 23(4): 363- 369.
[18] Cunningham J P, Moore C J, Zalucki1 M P, West S A. Learning, odour preference and flower foraging in moths. The Journal of Experimental Biology, 2004, 207(1): 87- 94.
[19] Cunningham J P, West S A. How host plant variability influences the advantages to learning: a theoretical model for oviposition behaviour in Lepidoptera. Journal of Theoretical Biology, 2008, 251(3): 404- 410.
[20] Cunningham J P, West S A, Zalucki M P. Host selection in phytophagous insects: a new explanation for learning in adults. Oikos, 2001, 95(3): 537- 543.
[21] Knudsen J T, Tollsten L. Trends in floral scent chemistry in pollination syndromes: floral scent composition in moth-pollinated taxa. Botanical Journal of the Linnean Society, 1993, 113(3): 263- 284.
[22] 徐廣, 郭予元, 吳孔明. 棉鈴蟲成蟲攜帶花粉的分析. 中國農業科學, 1999, 32(6): 63- 68.
[23] Corbet S A. Nectar sugar content: estimating standing crop and secretion rate in the field. Apidologie, 2003, 34(1): 1- 10.
[24] Tzin V, Galili G. New insights into the shikimate and aromatic amino acids biosynthesis pathways in plants. Molecular Plant, 2010, 3(6): 956- 972.
[25] Pichersky E, Gershenzon J. The formation and function of plant volatiles: perfumes for pollinator attraction and defense. Current Opinion in Plant Biology, 2002, 5(3): 237- 243.
[26] 侯茂林, 盛承發. 成蟲取食對棉鈴蟲雌蛾繁殖的影響. 生態學報, 2000, 20(4): 601- 605.
Associative learning ofHelicoverpaarmigeraadults to two key floral odours
LI Weizheng, WANG Qiong, LI Huiling, WANG Jue, LI Yangyang, GUO Xianru, YUAN Guohui*
PlantProtectionCollege,HenanAgriculturalUniversity,Zhengzhou450002,China
As an important polyphagous agricultural insect pest species, the adults ofHelicoverpaarmigera(Lepidoptera: Noctuidae) have the traits and habit of feeding various flower nectar as complementary nutrition. Two common floral volatiles, phenylacetaldehyde and benzyl acetate, which being released from a number of nectar host and oviposition host plant species of many moths and exhibited fairly strong attractiveness in previous field trapping experiments, are the key olfactory cues on guiding the flower-visiting behaviour of various moth species, includingH.armigera. To elucidate the associative learning ability ofH.armigeramoths to these floral volatiles, we tested the odour choice preference of the male and female moths after different training treatments (na?ve, exposure, and conditioning) in a dual choice bioassay, as well as the electroantennographic (EAG) responses of the male moths to these odours (use cis- 3-hexen- 1-ol as a reference) after corresponding treatments. The results of the odour preference bioassay manifested that, the choice frequencies of the na?ve group (having no contact experience with the floral odours) did not differ significantly between phenylacetaldehyde and benzyl acetate; moreover the exposure treatment (exposure to these odours without the association of food rewarding) could not also induce significant alteration of the odour preference; interestingly, the conditioning treatment (conditioning with the pairs of conditioned stimuli [floral odour] and the unconditioned stimulus [sucrose solution]) significantly altered the odour preference pattern, suggesting thatH.armigeramoths could associate the olfactory cues with the presence of food reward. Comparison of the learning performance ofH.armigeraconditioned with different floral odours showed that, phenylacetaldehyde was easier to be associated with sucrose solution than benzyl acetate, and the choice response percents of the males and females to phenylacetaldehyde correspondingly reached up to 70.00% and 67.50% after conditioned, respectively, but the individuals in the benzyl acetate-conditioned group only exhibited a weak discrimination response to the paired odour sources, suggesting that the preference ofH.armigerato benzyl acetate was an innate response rather than later learned response. The difference in associative learning between the two sexes was also studied. The results suggested that the female moths had stronger learning capacity of associating floral odours with sucrose solution than that of the male moths, which might be caused by the differential nectar-feeding requirements between sexes, since that three behaviors (feeding, mating, and oviposition) are involved in the life of female moths while oviposition is absent in the life of male moths. The results of EAG recording showed that the EAG responses of all the three male groups (na?ve, exposure, and conditioning) to the two floral odours and a green leaf volatile (cis- 3-hexen- 1-ol) did not differ significantly, suggesting that the associative learning of the males to phenylacetaldehyde occurred at the level of central nervous system rather than at the level of peripheral nervous system such as the olfactory receptor in the antennae. Taken together, the associative learning experience of floral odours paired with food resource could further improve the selection preference ofH.armigeramoths to the odour used as the conditioned stimulus. Our results provided a foundation for the further optimization of the formula design and field application of floral odour-based attractants on the basis of the associative learning.
Helicoverpaarmigera; floral odour; associative learning; selection preference
國家公益性行業(農業)專項資助項目(201203036); 河南省教育廳自然科學研究基礎項目(12A210012)
2013- 08- 14;
2014- 06- 12
10.5846/stxb201308142079
*通訊作者Corresponding author.E-mail: hnndygh@126.com
李為爭, 王 瓊, 李慧玲, 王 玨, 李洋洋, 郭線茹, 原國輝.棉鈴蟲成蟲對兩種關鍵花香氣味的聯系性學習.生態學報,2015,35(11):3534- 3540.
Li W Z, Wang Q, Li H L, Wang J, Li Y Y, Guo X R, Yuan G H.Associative learning ofHelicoverpaarmigeraadults to two key floral odours.Acta Ecologica Sinica,2015,35(11):3534- 3540.
