白樺子代家系幼林期生長表現及適應性分析
2015-01-11 06:33:08徐煥文李志新彭儒勝尚福強鄒建軍滕文華劉桂豐
浙江農林大學學報 2015年6期
關鍵詞:生長
劉 宇,徐煥文,李志新,姜 靜,彭儒勝,尚福強,鄒建軍,滕文華,劉桂豐
(1.東北林業大學 林木遺傳育種國家重點實驗室,黑龍江 哈爾濱 150040;2.遼寧省楊樹研究所,遼寧 蓋州 115200;3.遼寧省森林經營研究所,遼寧 丹東 118002;4.吉林省林業科學研究院,吉林 長春130033)
白樺子代家系幼林期生長表現及適應性分析
劉 宇1,徐煥文1,李志新1,姜 靜1,彭儒勝2,尚福強3,鄒建軍4,滕文華1,劉桂豐1
(1.東北林業大學 林木遺傳育種國家重點實驗室,黑龍江 哈爾濱 150040;2.遼寧省楊樹研究所,遼寧 蓋州 115200;3.遼寧省森林經營研究所,遼寧 丹東 118002;4.吉林省林業科學研究院,吉林 長春130033)
林木子代家系的多點試驗是研究其適應性及穩定性的重要環節。以6個白樺Betula platyphylla三倍體家系、4個二倍體家系為材料,分別在尚志、輝南、凌海、丹東等4個試驗點營造子代測定林,對3年生樹高和地徑等性狀進行多地點聯合分析。結果表明:樹高在各試驗點均達到差異顯著水平(P<0.01);地徑僅在丹東、凌海試驗點各家系達到顯著水平(P<0.05)。尚志試驗點最優三倍體家系為314和319;輝南試驗點為318和320;凌海試驗點為314,316和320;丹東試驗點為316,318和319。采用生產力指數法及基因型分組法將參試家系分為3種類型,即高產非穩產型家系,包括202,314和320;低產穩產型家系,包括201,318和319;低產非穩產型家系,包括203,204和315。各試驗點入選的優良家系分別屬于高產非穩產型家系及低產穩產型家系。研究結果不僅為白樺三倍體家系的早期推廣提供參考,而且也為高世代三倍體制種園的改建提供重要依據。圖3表6參24
林木育種學;白樺四倍體子代;多點試驗;家系選擇;遺傳穩定性
植物多倍體是自然界中的普遍現象。自然界中大約35%的被子植物和70%的禾本科Gramineae植物均屬于多倍體[1],植物染色體的多倍體化,有效推動了植物的進化,是新物種形成的重要途徑之一[2]。植物的多倍體化往往也帶來器官的巨大性,由于林木主要被利用的是營養器官,且多年生習性又保證品種一旦育成后就可以長期持續利用。因此,與其他植物相比,林木多倍體育種的潛力更大、作用更為突出。有人利用歐洲山楊Populus tremula四倍體與美洲山楊Populus tremuloides二倍體雜交,選育出了樹冠狹窄、適應性強、生長快、抗銹病能力強的人工雜種三倍體新品種 ‘Astria’[3]。北京林業大學采用化學誘導技術獲得2n花粉,再通過雜交育種的方法選育出了三倍體毛白楊Populus tomentosa優良新品種,其材積生長不僅較二倍體快[4],同時也表現出對不同環境高強的適應能力以及抗病能力[5]。由于林木三倍體優良新品種表現出速生性、抗逆性以及高度不育性的特點而越來越被重視,成為當今研究的熱點之一。目前,已經在楊樹Populus,橡膠樹Hevea brasiliensis,漆樹Toxicodendron vernicifluum,桑樹Morus alba等多個樹種中均選育出了優良三倍體新品種,并在生產上大量推廣應用[6-13]。中國白樺Betula platyphylla倍性育種研究始于2004年,由本研究團隊采用秋水仙素誘導白樺種子創制了白樺四倍體[14],并營建了初級白樺三倍體制種園。根據種子活力、生長性狀初步選擇了部分優良白樺四倍體母樹[15-17]。在前期研究的基礎上,針對營建的多點子代測定林進行穩定性和適應性分析,是白樺四倍體子代家系推廣的重要環節。因此,研究團隊2011年在東北三省的4個試驗點開展了白樺三倍體家系的區域化試驗,通過多地點聯合分析確定各家系的穩定性和適應性,為今后白樺三倍體家系的推廣提供依據,也為制種園的建園母樹的改良提供參考。
1 材料與方法
1.1 試驗材料與試驗設計

表1 白樺雜交雙親及子代代碼Table1 Birch hybrid parents and progeny codes
2009年在東北林業大學白樺強化種子園中,以定植于白樺強化種子園內的四倍體白樺Q65為母本, 6株二倍體雜種白樺(B.platyphylla×B.pendula)1-11,2-3,2-4,2-9,3-8和3-10等為父本,通過控制授粉獲共獲得6個四倍體家系的種子(314,315,316,318,319和320),同時采集園中4個二倍體家系(201,202,203和204)自由授粉的種子(表1)。2010年 4月末于塑料大棚中育苗,2011年早春分別在黑龍江省尚志市帽兒山實驗林場、吉林省輝南縣石道河林場、遼寧省凌海縣金城良種場、丹東市五龍背新建村等4個試驗點開展造林試驗(表2),試驗林按完全隨機區組設計,20株小區,4次重復,株行距2 m×2 m。2012年秋季進行全林樹高、地徑及保存率調查。

表2 各試驗點的地理氣候條件Table2 Soil types and climatic factors of the test sites
1.2 數據處理
1.2.1 方差分析 運用Microsoft Excel進行數據處理,用SPSS.v16.0軟件進行方差分析及多重比較。多點聯合方差分析的線性模型為:xijk=μ+υj+ρjk+τi+(τυ)ij+εijk。其中:xijk第i個家系在第j個地點第k區組的平均觀測值;μ為群體的平均值;υj為地點j的效應值;ρjk為地點內的區組效應(隨機效應);τi為家系i的效應值;(τυ)ij為家系×地點的互作效應;εijk為隨機誤差。
各試驗地點內采用雙因素方差分析,線性模型為:xijk=μ+τi+ρk+εijk。其中:xijk為第i個家系在第k區組的平均觀測值;ρk為區組效應(隨機效應);εijk為隨機誤差。
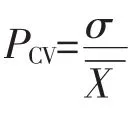
1.2.2 隸屬函數法 隸屬函數值R(X)計算公式:R(Xi)=(Xi-Xmin)/(Xmax-Xmin),反隸屬函數值R(X)計算公式:R(Xi)=1-(Xi-Xmin)/(Xmax-Xmin)。其中:Xi為指標測定值,Xmin,Xmax為所有參試材料某一指標的最小值和最大值。
1.2.3 遺傳穩定性分析 ①回歸分析。以各家系在各試驗點的平均地徑與每一個試驗點所有家系的地徑平均值(即環境指數)的線性回歸系數(bi)估計遺傳穩定性。當bi=1時,認為家系具有平均穩定性;當bi<1時,表示該家系高于平均穩定性;當bi>1時,表示家系低于平均穩定性[18-19]。計算公式為:
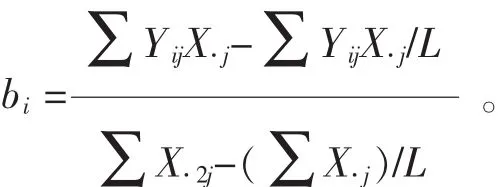
其中:bi表示第i家系回歸系數;Yij表示第i家系在j個參試點某性狀的平均值;X.j表示第j個參試點的環境指數;L表示各參試點個數。②Francis&Kannenberg基因型分組。以家系地徑平均值為縱坐標,變異系數為橫坐標繪制直方圖,并以所有參試家系地徑的總平均值和平均變異系數將其分為4組(圖1)。第Ⅰ組屬高產穩產型家系,第Ⅱ組是高產非穩產型家系,第Ⅲ組是低產穩產型家系,第Ⅳ組是低產非穩產型家系[20-21]。其中:家系地徑平均值(Yi)與變異系數(CV)計算如下:
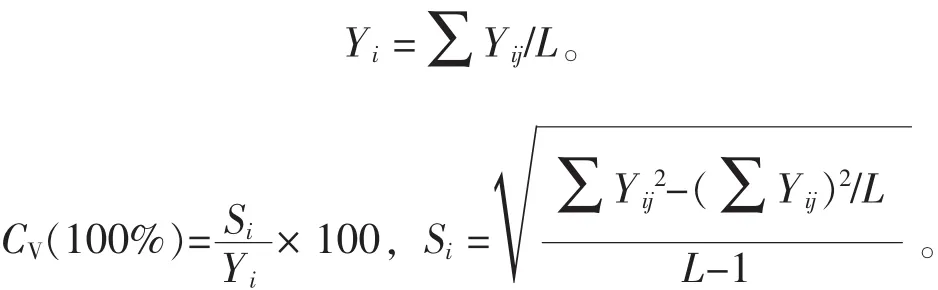
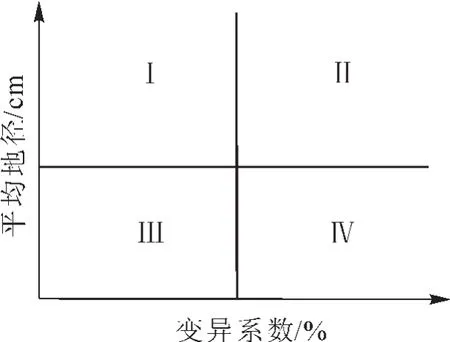
圖1 家系穩定性分組Figure 1 Family stability group
其中:Yij表示第i家系在j個參試點地徑的平均值;L表示各參試點個數。③生產力指數評價。生產力指數(IPQ)評價法:即生產力指數越大,表示該家系生長越快,反之越慢[22]。計算公式為:

其中:Xi-X…為家系效應,是所有試驗點i家系的地徑平均值與所有試驗點所有家系的平均值的離差;Σ∣X.j-X…∣為環境效應,是指各家系在j個參試點上的地徑平均值與家系總平均值離差的絕對值除以參試點數的商。
2 結果與分析
2.1 參試家系樹高和地徑及保存率的多點聯合方差分析
對4個試驗點10個參試家系的樹高、地徑以及保存率進行聯合方差分析表明(表3),在地點間各家系樹高、地徑、保存率差異均達到極顯著水平(P<0.01);在家系間的樹高、地徑、保存率也達到差異顯著(0.01<P<0.05)與極顯著水平(P<0.01);樹高與地徑在家系與地點的交互作用間達到顯著水平(0.01<P<0.05)。這一結果說明白樺家系在不同地點或不同的立地條件上生長表現與保存情況均不一致;基因型與環境之間存在互作。因此有必要開展區域性試驗,為白樺三倍體優良家系的推廣提供依據。
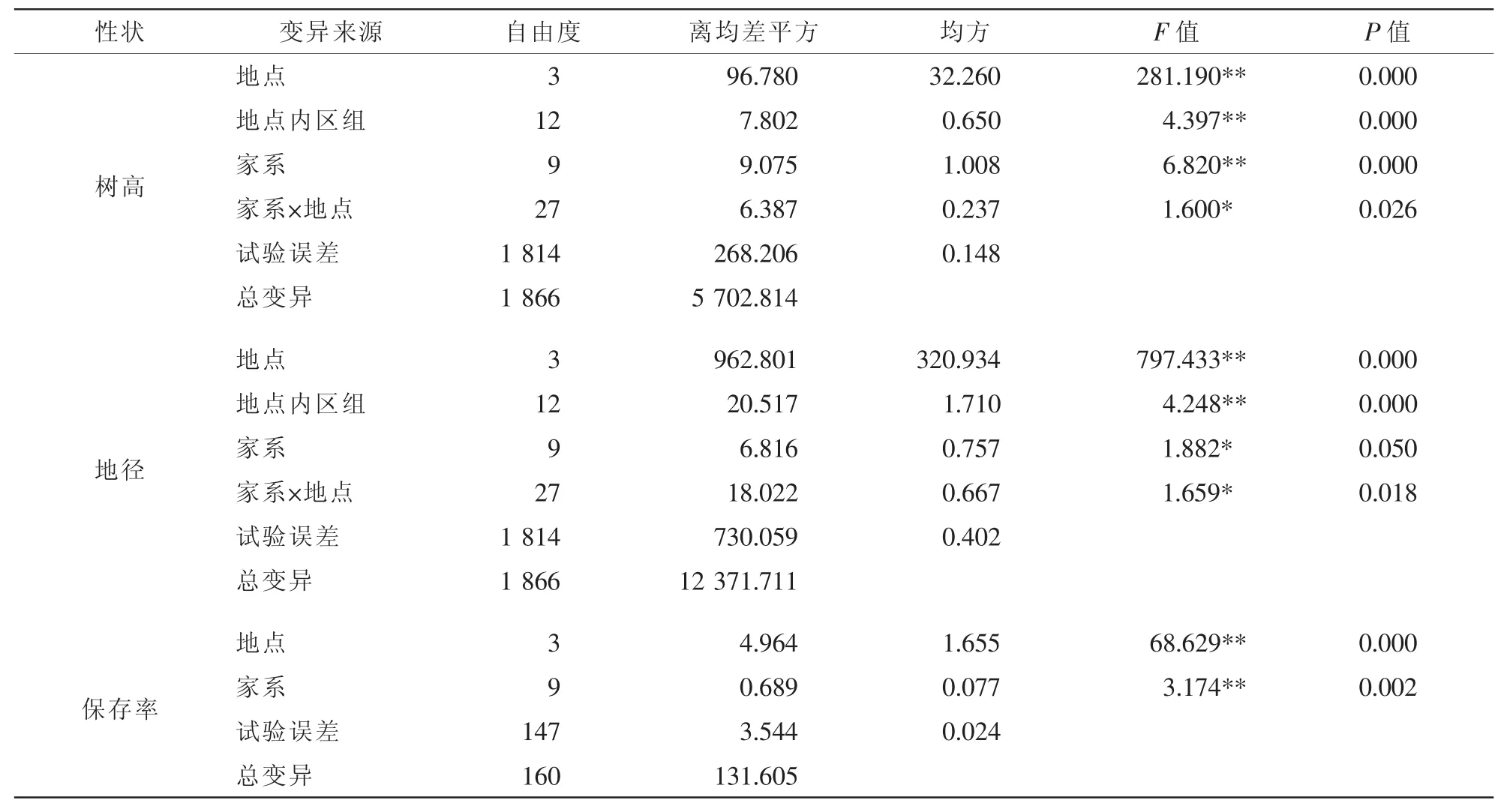
表3 參試家系生長性狀及保存率的多點聯合方差分析Table3 Analysis of variance among birch families for growth traits and preservation rate for the tested lines at different sites
2.2 各家系在不同試驗地點的生長性狀及保存率變異情況
采用雙因素方差分析模型對各試驗點10個家系的樹高和地徑進行方差分析(表4),結果顯示:在樹高生長方面,4個試驗點的家系間差異均達到顯著水平(0.01<P<0.05)或極顯著水平(P<0.01);而地徑生長只有凌海、丹東試驗點的家系間差異達到極顯著水平(P<0.01)。
進一步分析發現,在年均溫較高的遼寧省丹東和凌海2個試驗點無論是樹高還是地徑生長均顯著高于其他試驗點,家系樹高均值分別為1.85 m和1.80 m,地徑均值分別為3.05 m和2.91 cm,而在黑龍江尚志市的帽兒山試驗點,年均溫僅有2.4℃,樹高和地徑均值分別只有1.10 m和1.31 cm。總之,參試家系的生長呈現從北至南隨著年均溫的增加其生長量也增加的規律。
各參試試驗點的保存率不盡相同(表4),遼寧省凌海試驗點保存率最高,達到70%以上,而黑龍江尚志市的帽兒山試驗點的保存率最低僅為31.13%,保存率在4個試驗點的家系間差異均未達到顯著水平(P>0.05),說明各家系在各地區保存情況基本一致。
10個家系在4個試驗點的生長變異情況見表4。從樹高來看,凌海試驗點的變異系數最大,達到27.42%;其次是尚志試驗點,其樹高變異系數為24.34%;丹東試驗點的變異系數最小,僅有17.79%。從地徑來看,丹東試驗點的地徑變異系數最大,達到36.37%;其次是凌海,為34.78%;尚志試驗點的地徑變異系數最小,為23.23%。
4 個試驗點的參試家系樹高、地徑多重比較表明:從整體來看,各試驗點樹高和地徑排在前列的均為二倍體家系,有些三倍體家系雖然在生長表現方面遜色于二倍體家系,但各試驗點表現較好的三倍體家系如318和319等與表現最好的二倍體家系差異尚未達到顯著水平(表5)。這說明參試的三倍體家系雖然目前生長表現不是最優,但其較優良二倍體家系相差不大,今后通過更多優良四倍體母本與更多二倍體父本雜交,有望篩選獲得優良三倍體家系以及適于生產優良三倍體子代的雜交雙親。
采用隸屬函數法(表6)分別對4個試驗點的參試家系生長優劣進行評價,若以隸屬函數值大于0.45為入選優良家系, 則尚志試驗點是203, 314, 319, 202和204家系; 輝南試驗點是201,202,204,320和318家系;凌海試驗點是202,320,314,203,204,201家系;丹東試驗點大于該值的是204,202,203,318和316等家系。分析結果表明:參試家系的樹高及地徑綜合評價在各個試驗點并不完全一致,也說明不同家系在不同試驗點的適應能力有所不同。

表4 各試驗點參試家系生長性狀及保存率比較Table4 Genetic parameters among birch families for growth traits and preservation rate for the tested lines at different sites
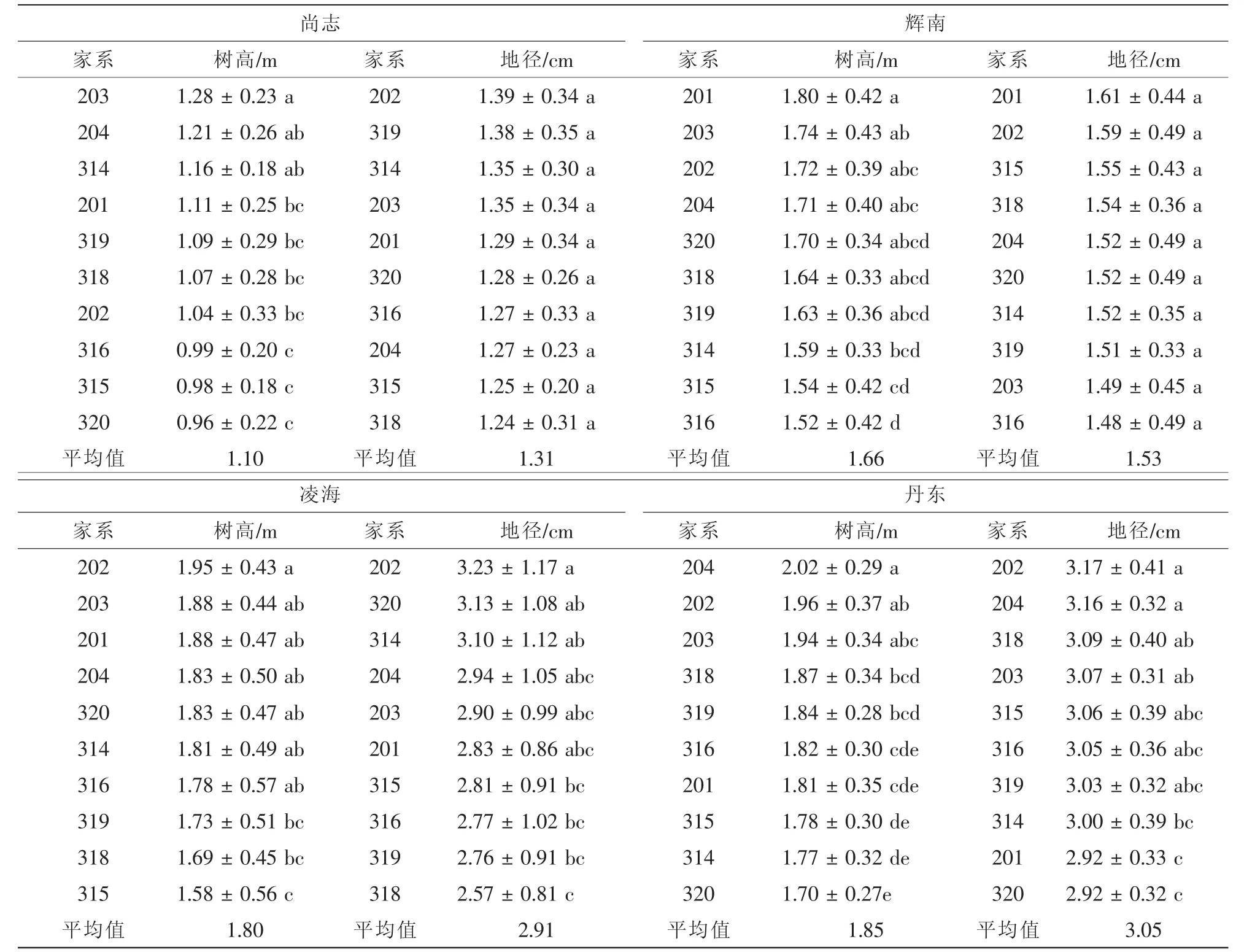
表5 各試驗地點參試家系樹高、地徑多重比較Table5 Multiple comparisons of birch height and base diameter for the tested lines at different sites
2.3 生長穩定性的多點聯合分析
由于地徑性狀在地點與家系間的交互作用達到了差異顯著的水平(0.01<P<0.05),說明不同地點、不同家系存在較大差異,有必要開展生長穩定性分析。因此,分別采用生產力指數法以及基因型分組法對10個家系的遺傳穩定性進行測定(圖2和圖3)。結果表明:除316家系外,上述2種方法區劃結果基本一致。

表6 各試驗地點參試家系隸屬函數值Table6 Subordinate function values for the tested lines at different sites
據此,將參試家系分為3種類型,即基因型分組法的第Ⅱ類高產非穩產型家系,包括202,314和320家系,這些家系綜合表現為具有較高的指標值,但遺傳穩定性較差,隨環境條件的變化而變化;第Ⅲ類為低產穩產型家系,包括201,318和319家系,這種類型的家系變異較小,具有良好的遺傳穩定性,可以適應較為惡劣的環境,但其生產力水平較低;第Ⅳ類為低產非穩產型家系,包括203,204和315家系,這類家系突出表現為家系內變異較大,遺傳穩定性較低,并且生產力也不高。上述2種方法分類結果基本一致,只有316家系被歸為不同的類型,對該家系暫不進行生長穩定性的劃分。

圖2 基因型分組法對家系穩定性劃分Figure 2 Gene grouping method for family stability group
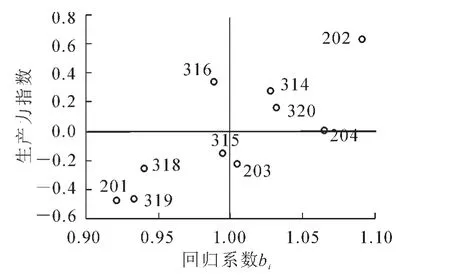
圖3 生產力指數與回歸系數法對家系穩定性劃分Figure 3 Productivity index and the regression coefficient for family stability group
3 結論與討論
自開展白樺倍性育種研究以來,對白樺四倍體及其子代的研究已有諸多報道,但這些研究還僅僅限于種子性狀及1年生苗期生長等方面[23,15-17]。本研究對早期獲得的三倍體家系于2010年在東北三省的4個地點營建子代測定林,3年生時調查其樹高、地徑以及保存率,進行了遺傳變異分析。結果表明:參試的10個家系(包括6個白樺三倍體全同胞家系、4個二倍體半同胞家系)樹高、地徑及保存率在地點間、家系間的差異均達到顯著或極顯著水平。丹東試驗點的樹高、地徑生長最好,家系樹高均值為1.85 m,地徑均值為3.05 cm。丹東造林點年均溫較高,平均年降水量大,適宜白樺的生長發育。
有研究指出,多倍體植物其發育時間要較同種二倍體植物遲緩[24]。本試驗由于參試對象為3年生的林分,尚處于發育初期,所以可能導致部分試驗點的三倍體家系間樹高、地徑性狀差異不顯著。而位于遼寧省的2個試驗點生長差異性顯著,這與試驗地點生長期長短及年積溫等條件相關。地處遼寧省的2個試驗點由于年均溫較高、生長期長、年降雨量大的氣候特點,白樺的生長量較大,6個三倍體家系間的生長特性表現較充分;而地處吉林省、黑龍江省的試驗點由于生長期短、年積溫較低,白樺生長量較小,所以親緣關系較近的6個三倍體家系間的特性差異尚未充分表達。
雖然參試家系在高生長和徑向生長上表現最優的均是二倍體家系,三倍體家系表現稍遜于二倍體家系,但各試驗點表現最好的三倍體家系均與該試驗點表現最好的二倍體家系差異未達到顯著水平,這說明白樺三倍體家系的選擇是具有潛力的。研究結果顯示,各試驗點同一母本不同父本的三倍體子代家系間存在較大差異,這說明在母本相同的情況下,子代雜種的差異是由二倍體父本不同決定的。這與研究團隊前期試驗結果一致,即白樺三倍體的生長既依賴于染色體加倍效應,又與親本的配合力水平有關[15]。由于本試驗僅有1株白樺四倍體母樹(Q65)達到了開花結實的年齡,無法進行多個母本與多個父本的雜交選配,因此,單一母本的遺傳基礎限制,可能也是三倍體家系生長表現略遜于二倍體家系的原因。
實現林木優良家系在一定區域范圍內的推廣,開展生長穩定性的多點聯合分析是重要環節之一。生產力高,產量穩定的家系是推廣的首選家系。對參試家系分別采用生產力指數與基因型分組法分類,10個家系在2種區劃方法中分布基本一致有3種家系,316家系劃分的不一致,建議隨著林齡的增長再對該家系進行研究劃分。
將各試驗點選擇的三倍體優良家系與生長穩定性分類結果進行比較發現:各試驗點的三倍體優良家系其穩定性均屬于第Ⅱ類和第Ⅲ類中,例如,屬于高產非穩產型的314和320家系,也在尚志、輝南、凌海試驗點入選的優良家系中;屬于低產穩產型的318和319家系,也在尚志、輝南、丹東試驗點入選的優良家系中。目前,由于生產單位對于白樺這一樹種利用方式的多樣化,我們的選育目標也隨之具有多個方向。對于高產非穩產型家系的選育主要側重于其在生長適宜地區能夠體現出產量增長迅速,成材所需時間短,木材產量高等方面的優勢;而對于低產穩產型家系的選育則側重于其生長適應能力強,可克服環境不良的影響,生長所需時間長則其木材材質勢必高于速生類型,因此可以滿足對材質要求較高的需求。我們建議對高產非穩產型的314和320家系在參試造林點及環境條件相近的毗鄰地區進行推廣。由于試驗分析的林齡只有3年生,對營建的子代測定林的材積分析也是我們的后續研究。隨著白樺制種園中四倍體母樹的陸續開花結實,研究團隊在后續的白樺四倍體育種中正在加大對親本的選配,采用交配設計方法進一步深入白樺四倍體子代的遺傳研究。
[1] 郭啟高,宋明,梁國魯.植物多倍體誘導育種研究進展[J].生物學通報,2000,35(2):8-10.
GUO Qigao,SONG Ming,LIANG Guolu.Plant induced polyploidy breeding research progress[J].Bull Biol,2000, 35(2):8-10.
[2] 孫敏紅,張蜀寧.多倍體育種在園藝作物中的應用[J].江蘇農業科學,2004,32(1):68-72.
SUN Minhong,ZHANG Shuning.Polyploid breeding in application of horticultural crops[J].Jiangsu Agric Sci,2004, 32(1):68-72.
[3] WEISGERBER H,RAU H M,GARTNER E J,et al.25 years of forest tree breeding in Hesse[J].Allg Forstztg,1980, 26:665-712.
[4] 康向陽.三倍體毛白楊新品種選育[J].北京林業大學學報,2004,26(3):40,63.
KANG Xiangyang.Triploid Populus tomentosa new species breeding[J].J Beijing For Univ,2004,26(3):40,63.
[5] 高克姝.三倍體毛白楊新品種對比試驗[J].山西林業科技,1999(1):5-9.
GAO Keshu.Contrast test of triploid Populus tomentosa new species[J].J Shanxi For Sci Technol,1999(1):5-9.
[6] EINSPAHR D W.Production and utilization of triploid hybrid aspen[J].Iowa State J Res,1984,58(4):401-409.
[7] ZHANG Zhenghai,KANG Xiangyang,ZHANG Pingdong,et al.Incidence and molecular markers of 2n pollen in Populus tomentosa Carr.[J].Euphytica,2007,154(1/2):145-152.
[8] ZHANG Zhenghai,KANG Xiangyang.Cytological characteristics of numerically unreduced pollen production in Populus tomentosa Carr.[J].Euphytica,2010,173(2):151-159.
[9] 李云,馮大領.木本植物多倍體育種研究進展[J].植物學通報,2005,22(3):375-382.
LI Yun,FENG Daling.Advances in research into polyploidy breeding of woody plants[J].Chin Bull Bot,2005,22(3):375-382.
[10] 康向陽.林木多倍體育種研究進展[J].北京林業大學學報,2003,25(4):70-74.
KANG Xiangyang.Advances in researches on polyploid breeding of forest trees[J].J Beijing For Univ,2003,25(4):70-74.
[11] 賴杭桂,陳霞,徐洪偉,等.橡膠樹三倍體種質創制及生物學鑒定[J].熱帶作物學報,2013,34(6):1001-1006.
LAI Hanggui,CHEN Xia,XU Hongwei,et al.Creation and biological identification of triploid germplasm in rubbertree[J].Chin J Trop Crop,2013,34(6):1001-1006.
[12] 尚宗燕,張繼祖,劉謙虎,等.漆樹染色體觀察及三倍體漆樹的發現[J].西北植物學報,1985,5(3):187-191.
SHANG Zongyan,ZHANG Jizu,LIU Qianhu,et al.The observation on chromosome of Rhus verniciflua stokes and discovery of triploid lacquer tree[J].Acta Bot Boreali-Occident Sin,1985,5(3):187-191.
[13] 楊今后,楊新華.桑樹人工三倍體育種的研究[J].蠶業科學,1989,15(2):65-70.
YANG Jinhou,YANG Xinhua.Breeding of artificial triploid in mulberry[J].Ser Sin,1989,15(2):65-70.
[14] 劉福妹,穆懷志,劉子嘉,等.秋水仙素處理不同家系白樺種子誘導四倍體的研究[J].北京林業大學學報, 2013,35(3):87-92.
LIU Fumei,MU Huaizhi,LIU Zijia,et al.Inducing tetraploid of Betula platyphylla with different generations of seeds by colchicine[J].J Beijing For Univ,2013,35(3):87-92.
[15] 林琳,穆懷志,姚啟超,等.白樺不同雜交組合三倍體子代當年生苗木生長性狀分析[J].北京林業大學學報, 2012,34(5):1-5.
LIN Lin,MU Huaizhi,YAO Qichao,et al.Growth traits of triploid progenies in one year old from different crosses of Betula platyphylla Suk.[J].J Beijing For Univ,2012,34(5):1-5.
[16] 徐煥文,劉宇,李雅婧,等.白樺三倍體制種園中各家系種子活力比較[J].西南林業大學學報,2013,33(5):34-39.
XU Huanwen,LIU Yu,LI Yajing,et al.Comparative analysis on seed vitality of different families produced in Betula pendula triploid seed orchard[J].J Southwest For Univ,2013.33(5):34-39.
[17] 劉宇,徐煥文,姜靜,等.基于種子活力及苗期生長性狀的白樺四倍體半同胞家系初選[J].北京林業大學學報, 2014,36(2):74-80.
LIU Yu,XU Huanwen,JIANG Jing,et al.Family selection of birch tetraploid half-sibling based on seed vigor and seedling growth traits[J].J Beijing For Univ,2014,36(2):74-80.
[18] 楊傳平,楊書文,劉桂豐,等.長白落葉松生長性狀的穩定性分析[J].東北林業大學學報,1991,19(育種專刊):32-37.
YANG Chuanping,YANG Shuwen,LIU Guifeng,et al.The stability analysis of the growth character of Larix olgensis[J].J Northeast For Univ,1991,19(Spiss):32-37.
[19] 王秋玉,楊書文,劉桂豐,等.紅皮云杉遺傳穩定性的研究及最佳種源選擇[J].東北林業大學學報,1993,21(1):5-12.
WANG Qiuyu,YANG Shuwen,LIU Guifeng,et al.Astudy on the genetic stability of Picea korraiesis and the optimal provenance selection[J].J Northeast For Univ,1993,21(1):5-12.
[20] 吳幼媚,王鵬良,韋穎文,等.桉樹無性系區域試驗[J].林業科技開發,2008,22(2):64-67.
WU Youmei,WANG Pengliang,WEI Yingwen,et al.Regional tests of clones of Eucalyptus[J].China For Sci Technol,2008,22(2):64-67.
[21] 李培,華勁松,王清,等.幾種產量分析法在蕓豆多點試驗中的應用[J].西昌學院學報:自然科學版,2011,25(3):9-12.
LI Pei,HUA Jinsong,WANG Qing,et al.Application of several yield analytical methods on kidney bean’s multilocation test[J].J Xichang Coll Nat Sci Ed,2011,25(3):9-12.
[22] 劉桂豐,楊書文,楊傳平,等.樟子松種源試驗的研究——遺傳穩定性測定及最佳種源選擇[J].東北林業大學學報,1991,19(2):17-23.
LIU Guifeng,YANG Shuwen,YANG Chuanping.et al.Studies on the provenances test of mongolian scots pine:the genetic stability determination and the optimal provenance selection[J].J Northeast For Univ,1991,19(2):17-23.
[23] MU Hhuaizhi,JIANG Jing,LI Huiyu,et al.Seed vigor,photosynthesis and early growth of saplings of different triploid Betula families[J].Dendrobiology,2012,68(2):11-20.
[24] 楊俊寶,彭正松.多倍體植物的表觀遺傳現象[J].遺傳,2005,27(2):335-342.
YANG Junbao,PENG Zhengsong.Epigenetic phenomena of plant polyploids[J].Hereditas,2005,27(2):335-342.
Growth performance and adaptability of Betula platyphylla offspring in the period of young forest
LIU Yu1,XU Huanwen1,LI Zhixin1,JIANG Jing1,PENG Rusheng2,SHANG Fuqiang3,ZOU Jianjun4, TENG Wenhua1,LIU Guifeng1
(1.State Key Laboratory of Tree Genetics and Breeding,Northeast Forestry University,Harbin 150040,Heilongjiang, China;2.Poplar Institute of Liaoning Province,Gaizhou 115200,Liaoning,China;3.Liaoning Institute of Forest Management,Dandong 118002,Liaoning,China;4.Jilin Provincial Academy of Forestry Science,Changchun 130033, Jilin,China)
Multi-point testing of tree progeny,an important link in the study of tree suitability and stability,was used to determine the height and ground diameter of Betula platyphylla (birch)for six triploid families and four diploid families planted at four sites:Shangzhi,Huinan,Linghai,and Dandong.3 years ago a multi-point conjoint analysis was used for differences in height between sites.Results of the multi-point conjoint analysis of height and ground diameter showed that differences in height between the sites were highly significant(P<0.01).Also,ground diameter for the Dandong and Linghai sites was significantly different(P<0.05).Families 314 and 319 were excellent triploid families at the Shangzhi site;at Huinan they were 318 and 320;at Linghai:314,316 and 320;and at Dandong:316,318 and 319.Then,tested families were divided into three types according to their productivity index and genotype grouping methods.The first type included high yieldinstability families 202,314 and 320.201,318 and 319 belonged to the low yield-stability type.The low yieldinstability type contained families 203,204 and 315.Excellent families selected from all sites belonged to thefirst and second type.This study not only provided a reference for early popularization of birch triploid families, but also formed an important basis for alterations of high generation triploid seed orchards.[Ch,3 fig.6 tab.24 ref.]
forest tree breeding;birch(Betula platyphylla)tetraploid progeny;multi-point test;family selection;genetic stability
S722.3
A
2095-0756(2015)06-0853-08
浙 江 農 林 大 學 學 報,2015,32(6):853-860
Journal of Zhejiang A&F University
10.11833/j.issn.2095-0756.2015.06.005
2015-01-23;
2015-04-01
“十二五”國家科技支撐計劃項目(2012BAD01B05)
劉宇,從事林木遺傳育種研究。E-mail:nefuly@126.com。通信作者:劉桂豐,教授,博士生導師,從事林木遺傳育種研究。E-mail:liuguifeng@163.com
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
