蔬菜土傳病害生防木霉菌株資源的篩選及其防治效果評(píng)價(jià)
2015-01-04 00:57:26
中國(guó)蔬菜 2015年8期
(農(nóng)業(yè)部蔬菜遺傳改良重點(diǎn)開放實(shí)驗(yàn)室,中國(guó)農(nóng)業(yè)科學(xué)院蔬菜花卉研究所,北京100081)
蔬菜土傳病害生防木霉菌株資源的篩選及其防治效果評(píng)價(jià)
朱萍萍 凌 健 席亞東 陳國(guó)華 茆振川 楊宇紅*謝丙炎
(農(nóng)業(yè)部蔬菜遺傳改良重點(diǎn)開放實(shí)驗(yàn)室,中國(guó)農(nóng)業(yè)科學(xué)院蔬菜花卉研究所,北京100081)
采用平板對(duì)峙接種法和溫室盆栽試驗(yàn),測(cè)定18種木霉共175個(gè)菌株對(duì)黃瓜枯萎病菌、辣椒疫霉病菌和番茄青枯病菌3種不同類型的蔬菜土傳病害病原菌的防效,篩選生防木霉資源。試驗(yàn)結(jié)果表明:拮抗試驗(yàn)中,拮抗黃瓜枯萎病菌的木霉菌株有50株,抑制率達(dá)70.0%~86.3%;拮抗辣椒疫霉病菌的木霉菌株43株,抑制率達(dá)70.4%~88.7%。盆栽試驗(yàn)中,篩選出番茄青枯病的生防菌株32株,防效達(dá)50.0%~92.7%。進(jìn)一步對(duì)在拮抗試驗(yàn)中對(duì)黃瓜枯萎病菌抑制率達(dá)75%以上的其中13株木霉菌進(jìn)行盆栽防效評(píng)價(jià),獲得防效超過60%的高效菌株2株,分別為T52(深綠木霉)和32080(平菇木霉)。
木霉;黃瓜枯萎病菌;辣椒疫霉病菌;番茄青枯病菌;生物防治
木霉作為一類重要的生防菌,其生境廣、環(huán)境適應(yīng)能力強(qiáng),在幾乎所有溫度的自然土壤中均有存在,在朽木、樹皮及其他真菌中(Kubicek et al.,2008)、甚至在昆蟲體內(nèi)(Yoder et al.,2008)、海洋中(Gal-Hemed et al.,2011)均能分離到木霉菌株。木霉生長(zhǎng)繁殖速度快,種群資源豐富,對(duì)植物有良好的促生或生防作用,是目前研究、應(yīng)用最多的生防菌之一。
特定種類的木霉不僅能顯著促進(jìn)植物的生長(zhǎng)發(fā) 育(Chang et al.,1986;Yedidia et al.,2001;Vinale et al.,2008),還能直接拮抗植物病原微生物,或通過誘導(dǎo)植物產(chǎn)生系統(tǒng)抗性及局部抗性等發(fā)揮生防作用(Hermosa et al.,2012),如朱廷恒等(2004)研究發(fā)現(xiàn)木霉菌株T97可通過競(jìng)爭(zhēng)和重寄生對(duì)多種植物病原真菌產(chǎn)生較強(qiáng)的拮抗作用;Shoresh等(2005)的研究結(jié)果顯示棘孢木霉菌株T203參與誘導(dǎo)黃瓜產(chǎn)生系統(tǒng)抗性的茉莉酸或水楊酸途徑;Moran-Diez等(2009)研究表明哈茨木霉菌株T34能通過分泌多聚半乳糖醛酸內(nèi)切酶誘導(dǎo)擬南芥(Arabidopsis)產(chǎn)生類似系統(tǒng)抗性的反應(yīng);Mastouri等(2012)報(bào)道哈茨木霉菌株T22處理番茄種子,可通過誘導(dǎo)葉綠體內(nèi)超氧化物歧化酶基因表達(dá),整株植物體內(nèi)該酶活性顯著提升,增強(qiáng)幼苗活力,減輕冷、熱及鹽等脅迫對(duì)植株的影響,從而促進(jìn)番茄生長(zhǎng)。因此,采用木霉生防植物病原菌,不僅可以顯著減少化學(xué)農(nóng)藥的使用,而且能通過促進(jìn)植物生長(zhǎng)等增強(qiáng)植物應(yīng)對(duì)生物、非生物因素脅迫的能力(Moran-Diez et al.,2009;Hermosa et al.,2012),進(jìn)而解決化學(xué)肥料、農(nóng)藥等帶來的一系列生態(tài)環(huán)境問題。雖然木霉資源豐富,但已知高效、穩(wěn)定的生防菌株資源仍然十分匱乏。為尋找能有效控制蔬菜主要土傳病害的木霉資源,本文以前期工作獲得的175株木霉菌株為原始材料,采用平板對(duì)峙及溫室盆栽防治的方法篩選對(duì)黃瓜枯萎病菌、辣椒疫霉病菌及番茄青枯病菌3種土傳病害病原菌具有拮抗或防治活性的木霉菌株,旨在為蔬菜土傳病害的生物防治提供新的菌株資源和科學(xué)依據(jù)。
1 材料與方法
1.1 供試菌株
供試木霉菌株共175株,包括18個(gè)種,主要為哈茨木霉(表1),由湖南農(nóng)業(yè)大學(xué)、湖南農(nóng)業(yè)科學(xué)院、河北科技師范學(xué)院、山東農(nóng)業(yè)科學(xué)院、中國(guó)農(nóng)業(yè)微生物菌種保藏中心、中國(guó)農(nóng)業(yè)科學(xué)院植物保護(hù)研究所和中國(guó)農(nóng)業(yè)科學(xué)院蔬菜花卉研究所7個(gè)單位提供。
供試致病菌株分別為Fuw-BN-7(黃瓜枯萎病菌,F(xiàn)usarium oxysporum f. sp. cucumebrium Owen)、Phy-01(辣椒疫霉病菌,Phytophthora capsici)、RS-60(番茄青枯病菌,Ralstonia solanacearum),由本所病害課題組分離純化保存。
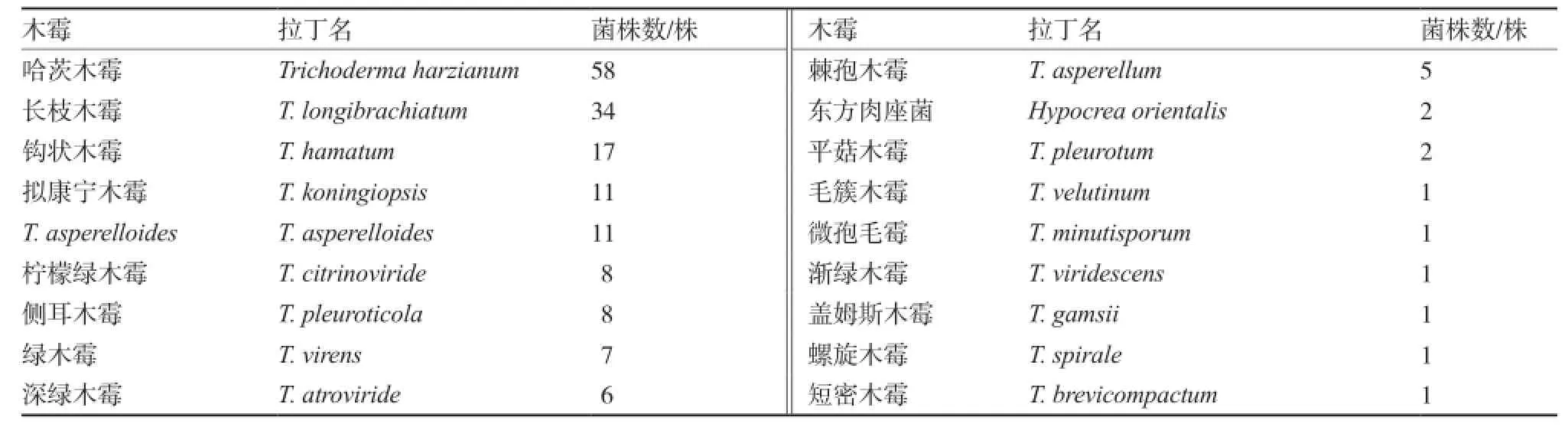
表1 供試木霉菌及其拉丁名
1.2 供試作物
供試黃瓜品種為中農(nóng)6號(hào),番茄品種為麗春,均購自中蔬種業(yè)科技(北京)有限公司。
1.3 拮抗效果測(cè)定
2014年4月,參考肖淑芹等(2011)的平板對(duì)峙接種方法,測(cè)定175株木霉對(duì)黃瓜枯萎病菌、辣椒疫霉病菌的拮抗效果。每處理3次重復(fù),每重復(fù)3個(gè)平板。28 ℃恒溫黑暗培養(yǎng)5 d后,分別測(cè)量木霉處理及對(duì)照種內(nèi)病原菌的菌落直徑,記錄菌落生長(zhǎng)狀況,計(jì)算抑制率。
抑制率(%)=(對(duì)照病原菌菌落直徑-處理病原菌菌落直徑)/(對(duì)照病原菌菌落直徑)×100%
1.4 溫室盆栽防效測(cè)定
1.4.1 木霉菌對(duì)番茄青枯病盆栽防效測(cè)定 將在PDA平板上28 ℃恒溫黑暗培養(yǎng)7 d的木霉菌用攪拌機(jī)攪碎,每皿加水400 mL,制成木霉菌液,于2014年4月番茄幼苗3~4葉期澆灌于植株根部,每株灌菌液30 mL;待木霉菌在土中定殖7 d后采用傷根灌注法接種番茄青枯病菌,接種濃度為1×107cfu·mL-1,接種時(shí)用刀片劃傷根部后每株用20 mL菌液灌根。以澆灌清水和硫酸鏈霉素800倍液分別作陰性對(duì)照(CK-)和陽性對(duì)照(CK+)。
1.4.2 木霉菌對(duì)黃瓜枯萎病盆栽防效測(cè)定 選擇對(duì)黃瓜枯萎病菌平板拮抗效果達(dá)75%以上的其中13株木霉菌,進(jìn)一步測(cè)定其溫室盆栽防治黃瓜枯萎病的效果。具體方法:2014年5月將在PDA平板上28 ℃恒溫黑暗培養(yǎng)7 d的木霉菌用攪拌機(jī)攪碎,每皿加水400 mL,制成木霉菌液,每盆育苗土接種2皿木霉菌攪碎后的菌液并混合均勻,置室溫下保濕培養(yǎng)7 d后,將黃瓜枯萎病菌采用胚根接種法接種(翁祖信 等,1985),接種后直接播于該育苗土上,以未經(jīng)木霉處理的育苗土作空白對(duì)照。
盆栽試驗(yàn)每處理3次重復(fù),每重復(fù)10株幼苗。待空白對(duì)照嚴(yán)重發(fā)病時(shí),調(diào)查各處理病株率或病級(jí)數(shù)(中華人民共和國(guó)農(nóng)業(yè)部,2010),并計(jì)算病情指數(shù)和防治效果。
病情指數(shù)=∑(各級(jí)病株數(shù)×相對(duì)應(yīng)級(jí)別數(shù))/(調(diào)查總株數(shù)×最高級(jí)值)×100
防治效果(%)=〔對(duì)照病情指數(shù)(或病株率)-處理病情指數(shù)(或病株率)〕/對(duì)照病情指數(shù)(或病株率)× 100%
2 結(jié)果與分析
2.1 木霉對(duì)黃瓜枯萎病菌的拮抗效果
供試的175株木霉菌對(duì)黃瓜枯萎病菌均有一定的拮抗作用,拮抗效果為39.1%~86.3%。其中,抑菌率達(dá)50%以上的木霉菌株有170株;抑菌率超過70%的木霉菌株50株(表2),包含13個(gè)種,主要為哈茨木霉、長(zhǎng)枝木霉及T. asperelloides;抑菌率達(dá)80%的木霉菌株4株,分別為長(zhǎng)枝木霉2株(T29、T60),綠木霉1株(T47),深綠木霉1株(T52)。木霉菌在PDA平板上對(duì)黃瓜枯萎病菌的拮抗效果主要表現(xiàn)為競(jìng)爭(zhēng)作用和重寄生作用(圖1),以競(jìng)爭(zhēng)作用為主,占供測(cè)菌株的87.1%,包括所有參試木霉種。
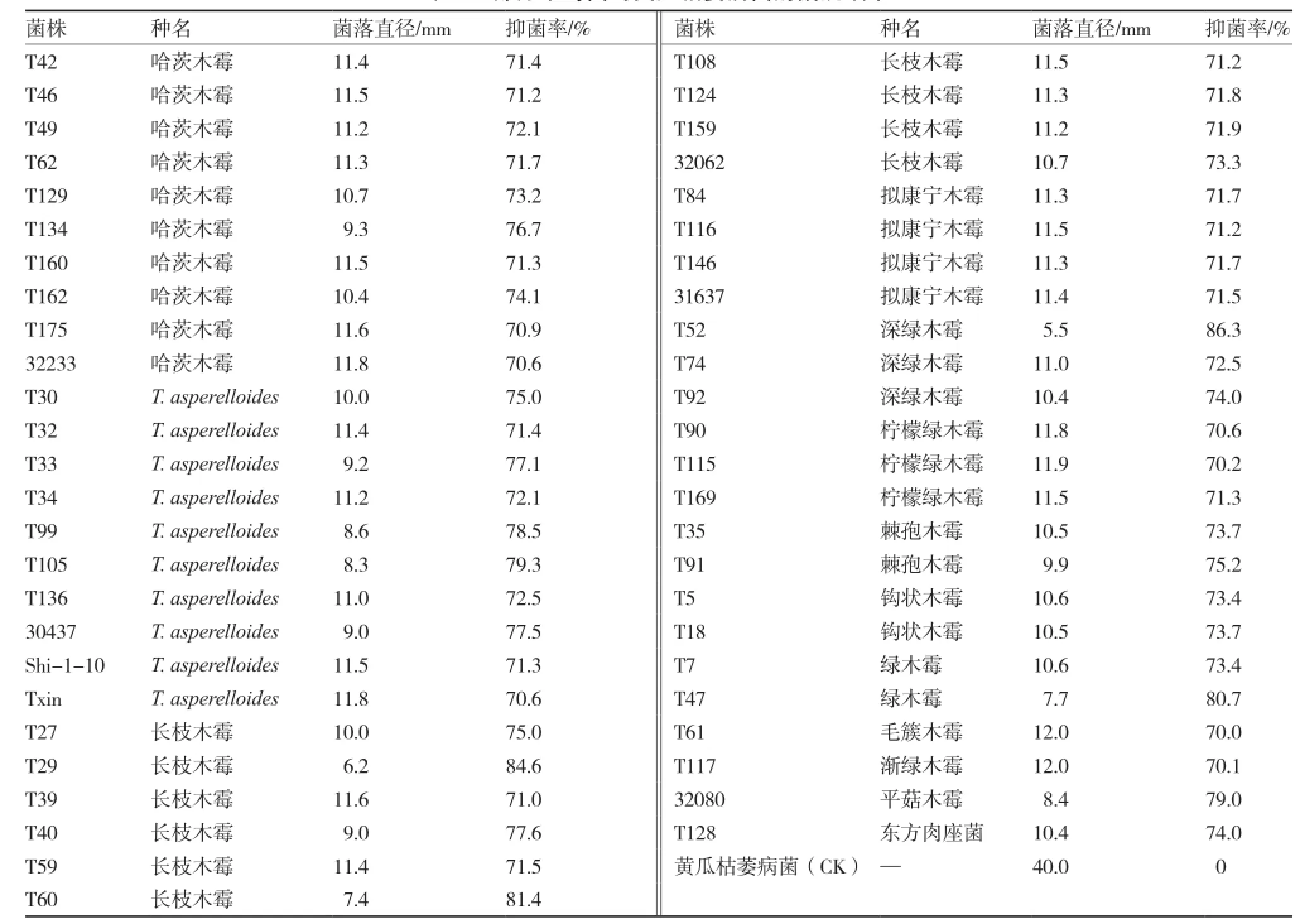
表2 部分木霉菌對(duì)黃瓜枯萎病菌的拮抗效果
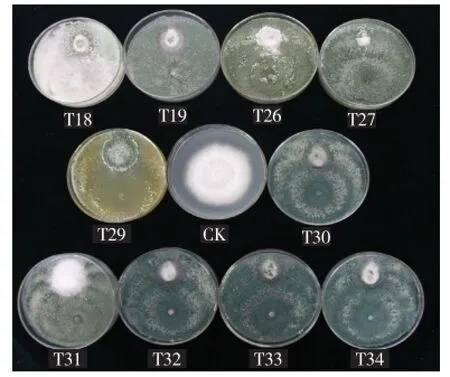
圖1 部分木霉菌對(duì)黃瓜枯萎病菌(平板上方)的拮抗效果
2.2 木霉對(duì)辣椒疫霉病菌的拮抗效果
參試木霉菌株對(duì)辣椒疫霉病菌的抑菌率為36.0%~88.7%,均表現(xiàn)一定的拮抗作用,且91.8%的參試木霉菌株的抑菌率在50%~80%。其中,抑菌率在70%以上的菌株有43株 (表3),包含7個(gè)種,主要為哈茨木霉、長(zhǎng)枝木霉及T. asperelloides;抑菌率超過80%的菌株10株,分別為哈茨木霉4株(T152、T175、Shi-8-9、mc-2),長(zhǎng)枝木霉2株(T29、T127),T. asperelloides 2株(T33、30437),檸檬綠木霉1株(T169),平菇木霉1株(32080)。木霉菌株拮抗辣椒疫霉病菌的機(jī)制主要為競(jìng)爭(zhēng)和重寄生(圖2),其中具有競(jìng)爭(zhēng)作用的菌株94株,包括所有的參試木霉種。
2.3 木霉對(duì)黃瓜枯萎病的盆栽防效
對(duì)平板拮抗試驗(yàn)中對(duì)黃瓜枯萎病菌抑菌效果達(dá)75%以上的其中13株木霉菌株進(jìn)行盆栽防效測(cè)定,發(fā)現(xiàn)供測(cè)木霉對(duì)黃瓜種子的出苗率、植株生長(zhǎng)均無不利影響,但總體防效不高,絕大部分木霉菌株的盆栽防效在40%以下,遠(yuǎn)低于離體拮抗效果(圖3),僅篩選出防效超過60%的木霉菌株2株,即T52(深綠木霉)和32080(平菇木霉),防效分別為72.0%和60.1%。
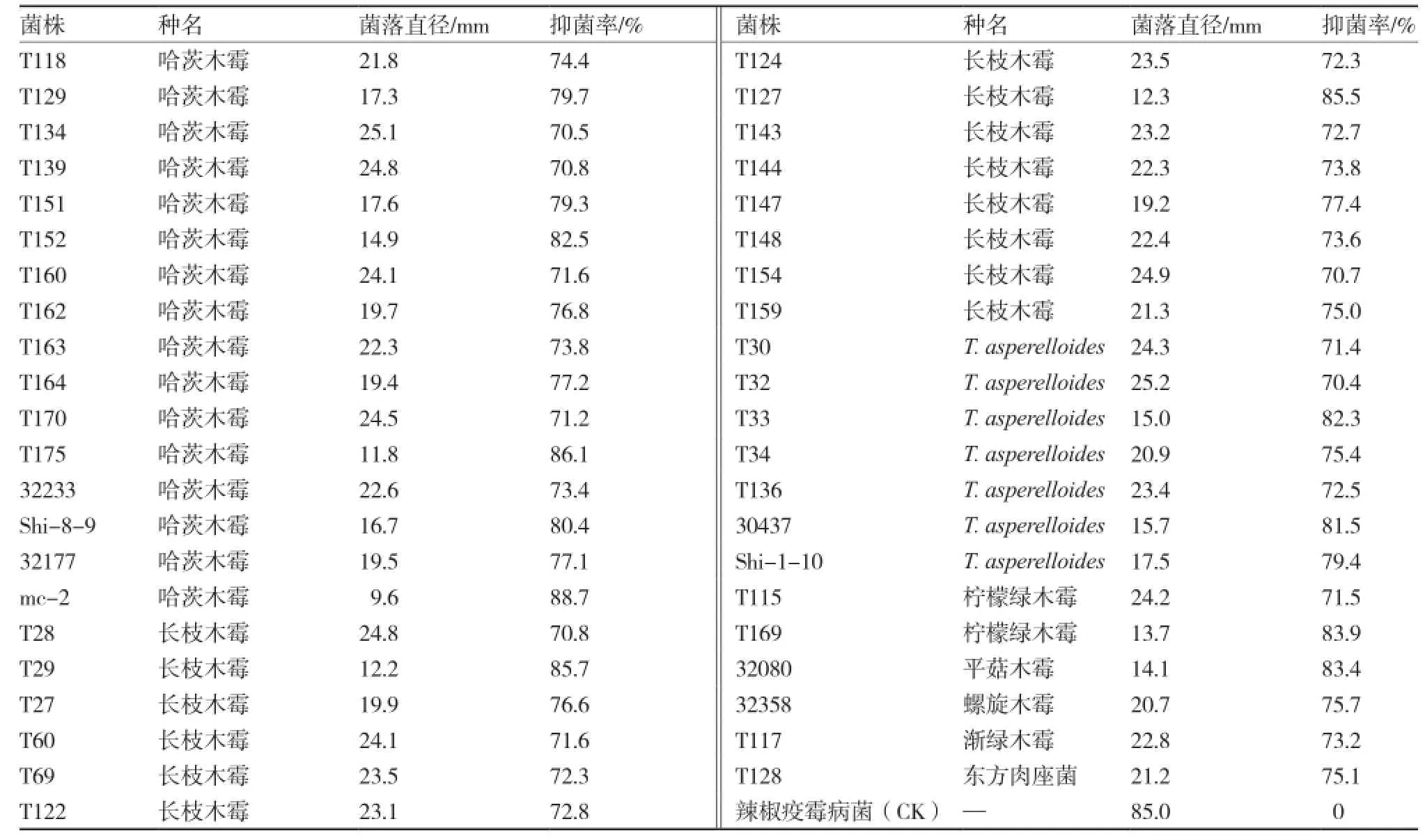
表3 部分木霉菌對(duì)辣椒疫霉病菌的拮抗效果
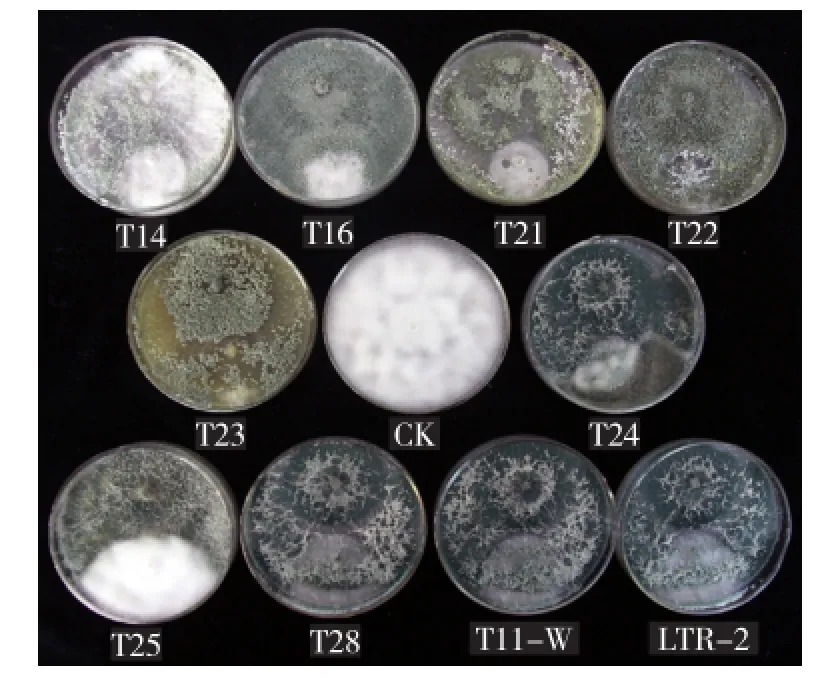
圖2 部分木霉菌對(duì)辣椒疫霉病菌(平板下方)的拮抗效果
2.4 木霉對(duì)番茄青枯病的盆栽防效
木霉盆栽防治番茄青枯病的測(cè)定結(jié)果表明,絕大多數(shù)木霉菌株對(duì)番茄青枯病盆栽防治效果不佳,一些木霉菌甚至加重青枯病的發(fā)生。在參試的175株木霉菌株中,防治青枯病效果達(dá)50%以上的木霉菌株有32株(表4),包含11個(gè)種,哈茨木霉菌株占46.9%;其中,防效接近或超過對(duì)照藥劑硫酸鏈霉素(69.2%)的木霉菌株20株,分別屬于9個(gè)種;防治效果達(dá)到80%以上的菌株4株,分別為T87(微孢木霉)、T173(哈茨木霉)、Shi-1-10(T. asperelloides)和18-8(棘孢木霉)。
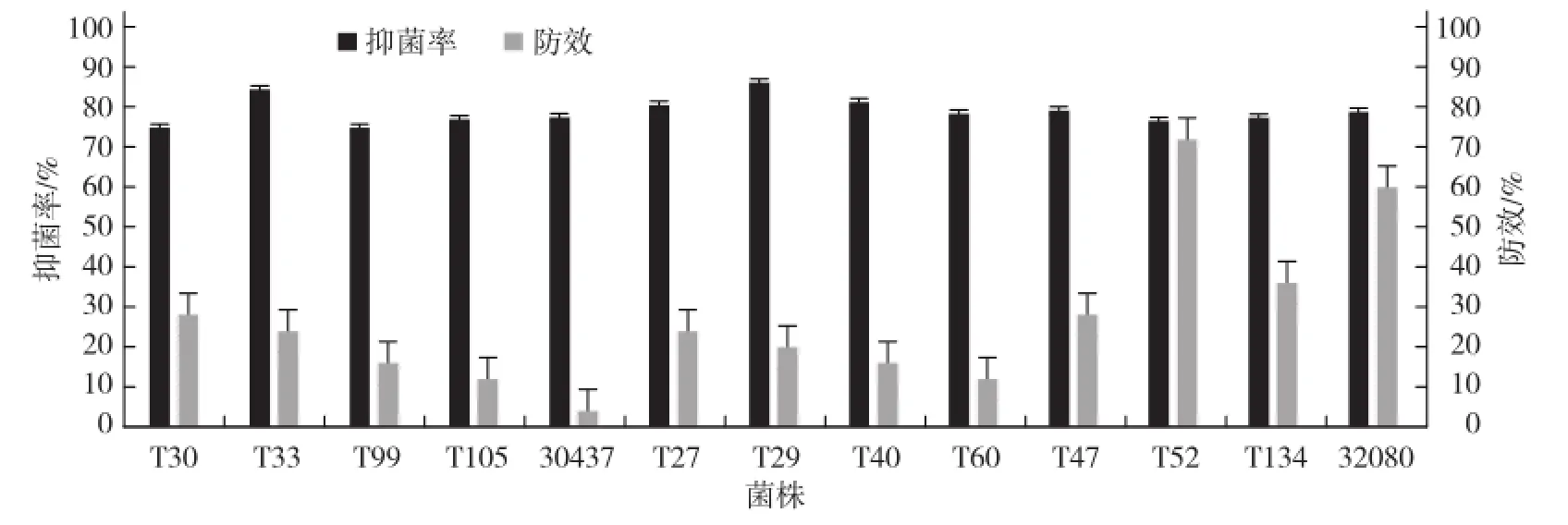
圖3 木霉菌對(duì)黃瓜枯萎病菌的平板拮抗作用和盆栽防治效果
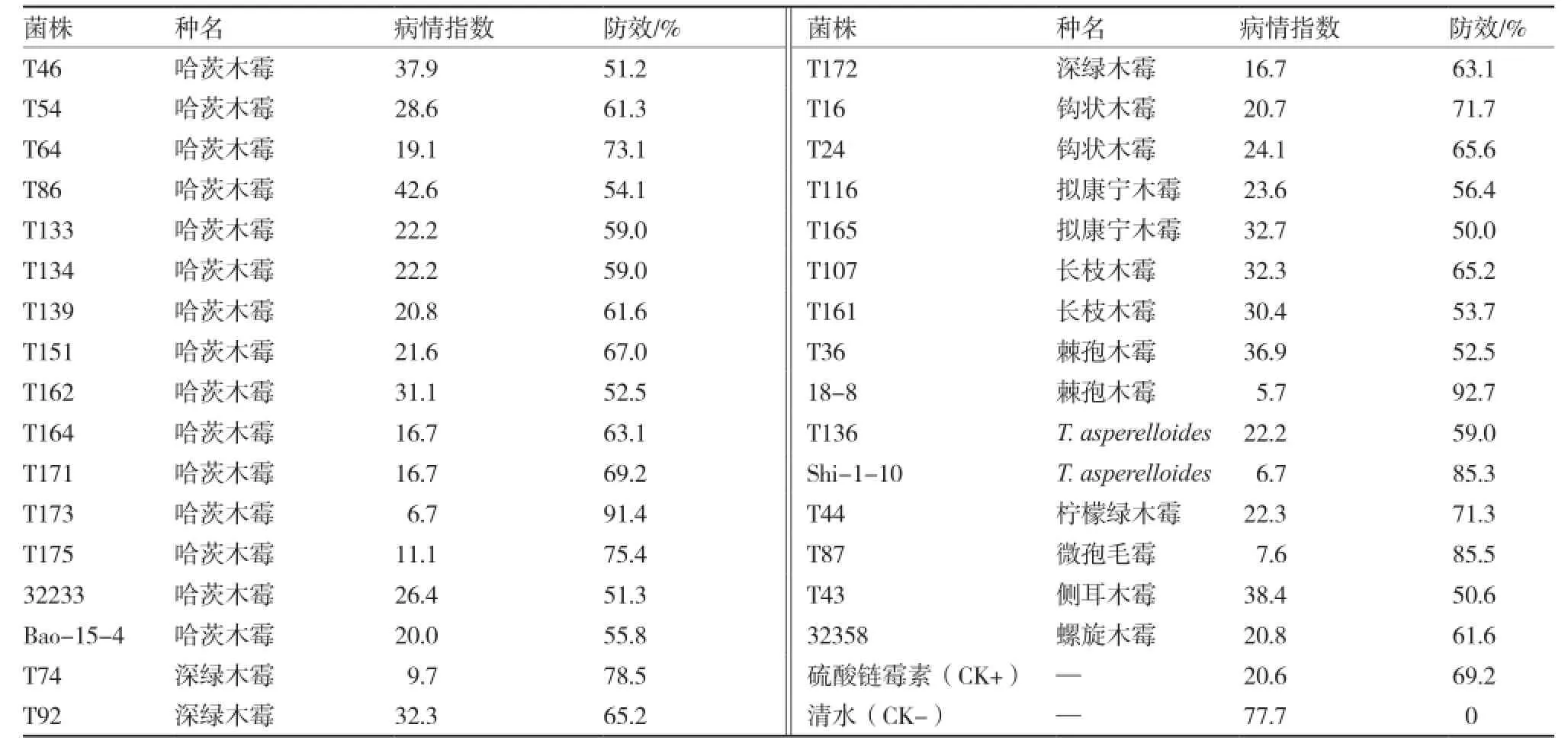
表4 部分木霉菌對(duì)番茄青枯病的盆栽防效
3 結(jié)論與討論
本試驗(yàn)中供試175株木霉菌株包含18個(gè)種,種質(zhì)資源豐富,且來源于全國(guó)多個(gè)省份農(nóng)作物種植區(qū)的土壤、腐朽的木頭及死亡線蟲蟲體等,已有研究表明部分菌株具有促進(jìn)植物生長(zhǎng)、生防蔬菜病原線蟲的潛力(未發(fā)表數(shù)據(jù))。相對(duì)于目前主要的生防木霉種哈茨木霉、長(zhǎng)枝木霉、鉤狀木霉和綠色木霉等,本試驗(yàn)擴(kuò)大了木霉種類的篩選范圍,發(fā)現(xiàn)一些研究甚少的木霉種類對(duì)蔬菜土傳病害具有較好的生防潛力。如T. asperelloides是近幾年才與棘孢木霉在種水平上區(qū)分開的一種新的木霉種,目前對(duì)其生防潛力的研究甚少。本試驗(yàn)通過對(duì)11株T. asperelloides生防3種主要蔬菜土傳病害病原菌的初步研究,篩選出針對(duì)不同病害有生防效果的菌株,為進(jìn)一步深入研究該種和挖掘新的生防木霉資源提供了科學(xué)依據(jù)。
哈茨木霉、長(zhǎng)枝木霉及T. asperelloides為平板對(duì)峙試驗(yàn)中篩選出生防菌株最多的3個(gè)種,且對(duì)黃瓜枯萎病菌的最高平板抑制率均超過75%,而對(duì)辣椒疫霉病菌則高達(dá)80%以上,為極具生防潛力的木霉種類,因此,在木霉生防菌株篩選時(shí)需要引起重視。
木霉菌株生防黃瓜枯萎病的平板對(duì)峙和溫室盆栽試驗(yàn)結(jié)果表明,木霉菌株對(duì)該病菌的拮抗作用并不與盆栽防效成正相關(guān)。這可能是因?yàn)槟久股L(zhǎng)速度快,因而在平板上生長(zhǎng)時(shí)具有明顯的競(jìng)爭(zhēng)優(yōu)勢(shì),而在盆栽試驗(yàn)中,木霉的生長(zhǎng)優(yōu)勢(shì)不一定能體現(xiàn)出來;除了共有的競(jìng)爭(zhēng)、抗生和重寄生等生防機(jī)制(Atanasova et al.,2013;劉增亮 等,2014;Abo-Elyousr et al.,2014)外,在溫室盆栽時(shí),木霉不僅需要與病原菌競(jìng)爭(zhēng)有限的空間和資源,還要與植物互作,通過與病原菌競(jìng)爭(zhēng)植物表面的生態(tài)位、誘導(dǎo)植物產(chǎn)生抗性等多種機(jī)制生防病原菌(Bae et al.,2009)。木霉菌的生防機(jī)制研究仍是以后的工作重點(diǎn)之一。
劉增亮,李梅,張玉杰,張林,楊曉燕,蔣細(xì)良.2014.哈茨木霉Thga1基因敲除突變株對(duì)幾種植物病原真菌的離體拮抗作用.中國(guó)生物防治學(xué)報(bào),30(3):370-375.
翁祖信,蔣興祥,肖小文.1985.黃瓜枯萎病抗病性鑒定方法研究——胚根接種法.中國(guó)蔬菜,(2):30-33.
肖淑芹,薛春生,曹遠(yuǎn)銀.2011.辣椒疫霉菌拮抗木霉的篩選及抑菌機(jī)制研究.北方園藝,(1):26-28.
中華人民共和國(guó)農(nóng)業(yè)部.2010.黃瓜主要病害抗病性鑒定技術(shù)規(guī)程第3部分:黃瓜抗枯萎病鑒定技術(shù)規(guī)程//中華人民共和國(guó)農(nóng)業(yè)行業(yè)標(biāo)準(zhǔn)NY/T 1857.3—2010.
朱廷恒,邢小平,孫順娣.2004.木霉T97菌株對(duì)幾種植物病原真菌的拮抗作用機(jī)制和溫室防治試驗(yàn).植物保護(hù)學(xué)報(bào),31(2):139-143.
Abo-Elyousr K A M,Abdel-Hafez S I I,Abdel-Rahim I R.2014.Isolation of Trichoderma and evaluation of their antagonistic potentialagainst Alternaria porri.Journal of Phytopathology,162(9):567-574.
Atanasova L,Le C S,Gruber S,Coulpier F,Seidl-Seiboth V,Kubicek C P,Druzhinina I S.2013.Comparative transcriptomics reveals different strategies of Trichoderma mycoparasitism.BMC Genomics,14:121-135.
Bae H,Sicher R C,Kim M S,Kim S H,Strem M D,Melnick R L,Bailey B A.2009.The beneficial endophyte Trichoderma hamatum isolate DIS 219b promotes growth and delays the onset of the drought response in theobroma cacao.J Exp Bot,60(11):3279-3295.
Chang Y C,Chang Y C,Baker R.1986.Increased growth of plants in the presence of the biological control agent Trichoderma harzianum.Plant Disease,70:145-148.
Gal-Hemed I,Atanasova L,Komon-Zelazowska M,Druzhinina I S,Viterbo A,Yarden O.2011.Marine isolates of Trichoderma spp. as potential halotolerant agents of biological control for arid-zone agriculture .Appl Environ Microbiol,77:5100-5109.
Hermosa R,Viterbo A,Chet I,Monte E.2012.Plant-beneficial effects of Trichoderma and of its genes.Microbiology,158:17-25.
Kubicek C P,Komon Z M,Druzhinina I S.2008.Fungal genus Hypocrea/Trichoderma:from barcodes to biodiversity.Journal of Zhejiang University Science B,9:753-763.
Mastouri F,Bj?rkman T,Harman G E.2012.Trichoderma harzianum enhances antioxidant defense of tomato seedlings and resistance to water deficit.Molecular Plant-microbe Interactions,25(9):1264-1271.
Moran-Diez E,Hermosa R,AmbrosinoP,Cardoza R E,Gutierrez S,Lorito M,Monte E.2009.The ThPG1 endopolygalacturonase is required for the Trichoderma harzianum-plant beneficial interaction.Molecular Plant-microbe Interactions,22(8):1021-1031.
Shoresh M,Yedidia I,Chet I.2005.Involvement of jasmonic acid/ ethylene signaling pathway in the systemic resistance induced in the systemic resistance induced in cucumber by Trichoderma asperellum T203.Phytopathology,95(1):76-84.
Vinale F,Sivasithamparam K,Ghisalberti E L,Marra R,Woo S L,Lorito M.2008.Trichoderma-plant-pathogen interactions. Soil Biology and Biochemistry,40:1-10.
Yedidia I,Srivastva A K,Kapulnik Y C.2001.Effect of Trichoderma harzianum on microelement concentrations and increased growth of cucumber plants.Plant and Soil,235:235-242.
Yoder J A,Glenn B D,Benoit J B,Zettler L W.2008.The giant Madagascar hissing-cockroach (Gromphadorhina portentosa)as a source of antagonistic moulds:concerns arising from its use in a public setting.Mycoses,51:95-98.
Screening and Bio-control Effect Assessment of Trichoderma Strains against 3 Soilborn Vegetable Pathogens
ZHU Ping-ping,LING Jian,XI Ya-dong,CHEN Guo-hua,MAO Zhen-chuan,YANG Yu-hong*,XIE Bing-yan
(Vegetable Genetic Improvement,Ministry of Agriculture Key Laboratory,Institute of Vegetables and Flowers,Chinese Academy of Agricultural Sciences, Beijing 100081,China)
This paper had screened the bio-control resources of 18 Trichoderma species,including 175 strains,against 3 kinds of soil-born vegetable pathogens as Fusarium oxysporum f. sp. cucumebrium Owen,Phytophthora capsici and Ralstonia solanacearum through confront culture experiments and green house potted plant experiments. The results indicated that through confront culture experiments,we have screened out 50 Trichoderma strains with inhibitation rate as 70.0%-86.3% against Fusarium oxysporum f. sp. cucumebrium Owen,and 43 Trichoderma strains with inhibitation rate as 70.4%-88.7% against Phytophthora capsici.And the potted plant experiments have screened out 32 Trichoderma strains with bio-control efficacy of 50.0%-92.7% against Ralstonia solanacearum.We further assessed the bio-control efficacy of 13 Trichoderma strains with antagonistic inhibitation rate higher than75% by potted plant experiment,and found 2 Trichoderma strains T52 (Trichoderma atroviride) and 32080 (Trichoderma pleurotum) with high bio-control efficacy over 60%.
Trichoderma spp.;Fusarium oxysporum f. sp. cucumebrium Owen;Phytophthora capsici;Ralstonia solanacearum;Bio-control
朱萍萍,女,碩士研究生,專業(yè)方向:分子植物病理學(xué),E-mail:ping574992751@126.com
*通訊作者(Corresponding author):楊宇紅,研究員,專業(yè)方向:植物病理學(xué),E-mail:yangyuhong@caas.cn
2015-03-09;接受日期:2015-04-21
國(guó)家公益性行業(yè)(農(nóng)業(yè))科研專項(xiàng)(201303014),國(guó)家大宗蔬菜產(chǎn)業(yè)技術(shù)體系建設(shè)項(xiàng)目(CARS-25-B-01),農(nóng)業(yè)部園藝作物遺傳改良重點(diǎn)開放實(shí)驗(yàn)室項(xiàng)目
