3 株乳酸菌及其組合對發(fā)酵牦牛肉灌腸中肽變化的影響
2015-01-03 01:14:59白菊紅唐善虎李思寧盧付青閆利國
食品科學(xué) 2015年17期
白菊紅,唐善虎*,李思寧,王 柳,盧付青,閆利國
3 株乳酸菌及其組合對發(fā)酵牦牛肉灌腸中肽變化的影響
白菊紅,唐善虎*,李思寧,王 柳,盧付青,閆利國
(西南民族大學(xué)生命科學(xué)與技術(shù)學(xué)院,四川 成都 6 10041)
本實(shí)驗(yàn)探討了3 株乳酸菌發(fā)酵劑及其組合在發(fā)酵牦牛肉灌腸過程中對肽變化規(guī)律的影響。人工接種米酒乳桿菌(Lactobacillus sake,L. s)、植物乳桿菌(Lactobacillus plantarum,L. p)、戊糖片球菌(Pediococcus pentosaceus,P. p)及其組合菌種于牦牛肉灌腸,每隔24 h測定其pH值,分別在主發(fā)酵2 d(32 ℃),再自然風(fēng)干發(fā)酵10、20、30 d后取樣,采用反相高效液相色譜(reversed phase high performance liquid chromatography,RP-HPLC)對不同 發(fā)酵時(shí)期牦牛肉灌腸中肽的含量進(jìn)行檢測和分析。結(jié)果表明,在主發(fā)酵2 d,再自然風(fēng)干發(fā)酵10、20 d(除P. p與L. p+P. p外)和30 d后,不同乳酸菌發(fā)酵劑對牦牛肉灌腸中總肽含量的影響均顯著(P≤0.05)。按發(fā)酵時(shí)間由短及長的順序,在4 個(gè)不 同發(fā)酵時(shí)期檢出最大總肽含量的發(fā)酵菌種依次是L. s(1.73×105mAU·s)、P. p(1.62×105mAU·s)、對照組(1.81×105mAU·s)和L. p(1.83×105mAU·s)。隨著發(fā)酵時(shí)間的延長,發(fā)酵牦牛肉灌腸中的總肽含量發(fā)生了明顯的變化,相對極性較弱的肽減少了,存在的主要是相對極性較大的肽;主發(fā)酵結(jié)束時(shí),L. s發(fā)酵的處理組檢出最大總肽含量,且L. s與其參與發(fā)酵L. s+P. p和L. s+L. p+P. p組合中檢出總肽含量的變化呈現(xiàn)出相似的趨勢,L. s是牦牛肉灌腸中快速生成肽的良好發(fā)酵劑;在4 個(gè)發(fā)酵時(shí)期,L. p+L. s發(fā)酵的處理組檢出肽的含量均小于其單一菌種的發(fā)酵,L. p+L. s對牦牛肉灌腸中肽的生成具有一定的拮抗作用;L. s與P. p作用不明顯,但能快速降低pH值,保證產(chǎn)品的安全性。本實(shí)驗(yàn)結(jié)果為進(jìn)一步生產(chǎn)優(yōu)質(zhì)牦牛肉灌腸奠定了良好的基礎(chǔ)。
乳酸菌;發(fā)酵;牦牛肉灌腸;肽類;反相高效液相色譜
牦牛是我國特有的牛品種,世界上90%以上的牦牛都集中在我國境內(nèi),牦牛肉中含有豐富的蛋白質(zhì)、必需氨基酸、礦物質(zhì)(Fe、Zn、Se)和VB,是優(yōu)質(zhì)的動物源食材[1]。目前,市場上牦牛肉產(chǎn)品基本上是采用傳統(tǒng)加工工藝加工而成,特別是風(fēng)干牦牛肉,因歷史悠久、風(fēng)味獨(dú)特在牦牛肉制品市場占有主導(dǎo)地位。但是,傳統(tǒng)加工方式多以自然風(fēng)干為主,時(shí)間周期長,容易導(dǎo)致牦牛肉中的脂肪氧化變質(zhì),不僅會產(chǎn)生難以接受的哈喇味,而且也會產(chǎn)生有毒有害的物質(zhì)[2]。
近年來研究發(fā)現(xiàn),將乳酸菌應(yīng)用于食品中可以縮短產(chǎn)品的加工周期,降低亞硝酸鹽含量,提高產(chǎn)品品質(zhì)[3-4]。因此,應(yīng)用乳酸菌開發(fā)新型的牦牛肉制品,對豐富牦牛肉制品市場有著重要的意義。
許慧卿等[5]研究了米酒乳桿菌對風(fēng)鴨肌肉蛋白質(zhì)降解的影響,發(fā)現(xiàn)米酒乳桿菌能促進(jìn)肌漿蛋白與肌原纖維蛋白的降解,能夠顯著增加游離氨基酸的總量。徐幸蓮等[6]研究了植物乳桿菌6003對肌肉蛋白的分解能力,結(jié)果表明植物乳桿菌6003的活細(xì)胞和細(xì)胞裂解液增加肌漿蛋白提取液中肽、游離氨基酸含量,活細(xì)胞和細(xì)胞裂解液兩者混合后的效果更為顯著。Rizzello等[7]對乳酸菌發(fā)酵谷類食物生成抗癌肽Lunasin進(jìn)行了研究,他們發(fā)現(xiàn)乳酸菌發(fā)酵谷類食品比對照產(chǎn)品中檢出的Lunasin含量增加了2~4 倍。但是,目前應(yīng)用乳酸菌及其復(fù)配發(fā)酵劑加工發(fā)酵牦牛肉灌腸,并研究其中肽變化規(guī)律尚未見報(bào)道。
肽是由20 種L-氨基酸中部分或全部種類以不同的排列方式和組成比例構(gòu)成的簡單二、三肽或復(fù)雜線性或環(huán)形結(jié)構(gòu)不同肽類的總稱,是由蛋白質(zhì)降解或生物體內(nèi)直接合成的多功能化合物[8]。國內(nèi)外大量研究報(bào)道認(rèn)為:肽在人體中可以像氨基酸一樣被吸收并直接進(jìn)入血液循環(huán),具有增強(qiáng)免疫、抗菌、抗病毒、調(diào)節(jié)激素、抗氧化、抗衰老,降血脂,降膽固醇等功能[9-11]。由于肽在微量狀態(tài)下具有強(qiáng)大的生物活性,并且具有食用安全性,肽類研究已成為當(dāng)前極具發(fā)展前景的功能因子和食品界最熱門的研究課題之一[12]。
本實(shí)驗(yàn)將米酒乳桿菌、植物乳桿菌、戊糖片球菌及其組合菌種接種于牦牛肉灌腸中,采用反相高效液相色譜技術(shù)(reversed phase high performance liquidchromatography,RP-HPLC)對不同發(fā)酵時(shí)期的牦牛肉灌腸中的肽進(jìn)行檢測,旨在探索3 株乳酸菌發(fā)酵劑及其組合對發(fā)酵牦牛肉灌腸中肽變化規(guī)律的影響,篩選出具有潛在市場價(jià)值的牦牛肉灌腸乳酸菌發(fā)酵劑,以此為牦牛肉新產(chǎn)品的開發(fā)提供參考。
1 材料與方法
1.1 材料、菌種、培養(yǎng)基與試劑
牦牛肉、豬肥膘、食鹽、蔗糖、五香粉、亞硝酸鈉(食品級)、味精、天然豬腸衣,均購于成都高升橋好又多超市。
米酒乳桿菌(Lactobacillus sake,L. s)、戊糖片球菌(Pediococcus pentosaceus,P. p)、植物乳桿菌(Lactobacillus plantarum,L. p),購于中國工業(yè)微生物菌種保藏管理中心。
MRS增菌培養(yǎng)基、MRS培養(yǎng)基購于杭州微生物試劑有限公司。
NaCl、NaH2PO4、Na2HPO4(分析純)、三氟乙酸、乙腈(色譜純) 成都市科龍化工試劑廠;氮?dú)?成都泰芋有限責(zé)任公司。
1.2 儀器與設(shè)備
Agilent 1100型高效液相色譜儀(配自動進(jìn)樣器和紫外檢測器) 美國Agilent公司;pH-Star儀 德國Matthaus公司;Centrifuge 5804離心機(jī) 德國Eppendorf公司;絞肉灌腸一體機(jī) 永康市卓越塑膠有限公司;PL303電子天平 瑞士Mettler Toledo公司;BCD-221TMBA冷凍冰箱 青島海爾股份有限公司;FSH-2A高速勻漿機(jī) 江蘇金壇市醫(yī)療儀器廠;YGC-12型氮吹儀 成都雅源科技有限公司;DHP-9052電熱恒溫培養(yǎng)箱 上海齊欣科學(xué)儀器有限公司;SW-CJ-1F超凈工作臺蘇凈集團(tuán)安泰公司。
1.3 方法
1.3.1 發(fā)酵劑制備
參考蔣云升[13]和王永霞[14]等的方法并修改進(jìn)行。將L. s、P. p、L. p分別按V(菌種)∶V(培養(yǎng)基)=1∶20接種于MRS增菌培養(yǎng)基,30 ℃條件下恒溫培養(yǎng)24 h左右,重復(fù)此操作兩次,當(dāng)培養(yǎng)基呈明顯混濁時(shí),活化完成,保存于4 ℃冰箱。應(yīng)用MRS培養(yǎng)基對活化好的菌種進(jìn)行平板活菌計(jì)數(shù)。
1.3.2 牦牛肉灌腸制作工藝
工藝流程:原料肉→切塊→混勻→腌制→絞肉→接種→灌腸→結(jié)扎、排氣→主發(fā)酵→自然風(fēng)干發(fā)酵→成熟→成品。
操作要點(diǎn):1)配方:主料:牦牛肉80%、豬肥膘20%;輔料:以主料100%計(jì),食鹽3.5%、蔗糖5.6%、亞硝酸鈉0.05%、五香粉0.45%、味精0.4%;2)切塊:分別將清洗并修整除去筋膜、筋腱的牦牛肉和豬肥膘切成約2 cm×2 cm×2 cm大小的塊狀,并瀝干表面水分;3)混勻與腌制:將肉塊與輔料混合均勻后置于4 ℃冰箱腌制約24 h,當(dāng)肉呈現(xiàn)出內(nèi)外一致的鮮紅色、不綿軟、有汁液滲出,且用手觸摸時(shí)有堅(jiān)實(shí)感、滑膩感時(shí),完成腌制;4)絞肉:用絞肉機(jī)將腌制好的肉塊絞為均勻的肉餡。5)接種:按表1菌種組合及配比,以1×106CFU/g接種量分別接種于絞好的肉餡。6)灌裝:灌裝時(shí)要掌握腸體的飽滿程度,不能松軟,也不宜過飽。7)結(jié)扎、排氣:灌裝好的腸體每隔10~20 cm用線繩結(jié)扎成一節(jié),用排氣針有規(guī)律地扎眼,排除腸體中的空氣。8)主發(fā)酵:用清水漂洗干凈灌裝好的腸體,32 ℃恒溫發(fā)酵2 d,當(dāng)接種乳酸菌發(fā)酵劑的灌腸pH值降至5.3~5.0時(shí),主發(fā)酵結(jié)束。9)自然風(fēng)干發(fā)酵:主發(fā)酵結(jié)束后,將灌腸懸掛于陰涼通風(fēng)處,于自然條件(13~22 ℃)下繼續(xù)發(fā)酵30 d。

表1 菌種組合及配比Table1 Bacterial strains and their combinations
1.3.3 測定方法
在整個(gè)發(fā)酵過程中,每24 h測定一次pH值,至第15天。分別在主發(fā)酵(2 d)、繼續(xù)自然發(fā)酵10、20、30 d結(jié)束后取樣,均保存于-18 ℃冰箱,用于蛋白肽檢測。
1.3.3.1 pH值測定
用pH Star儀測定牦牛肉灌腸的pH值,每個(gè)樣本重復(fù)3 次,取平均值。
1.3.3.2 肽含量的測定方法
[15-17]略做修改進(jìn)行。
樣品前處理和制備:準(zhǔn)確稱取3 g待測樣品,加入30 mL 0.2 mol/L磷酸鹽緩沖液(phosphate buffered saline,PBS)(NaH2PO4、Na2HPO4,pH 6.5),高速勻漿5 min(6 000 r/min),然后高速離心20 min(10 000×g)。取上清液2.5 mL,加入5 mL乙腈,高速離心20 min(15 000×g),取上清液5 mL,用氮吹儀吹去溶劑,用400 μL體積分?jǐn)?shù)為0. 05%三氟乙酸溶解殘留物,過0.22 μm濾膜后進(jìn)行HPLC測定,每個(gè)樣本重復(fù)3 次。
色譜條件:色譜柱:Waters Symmetry C18色譜柱(4.6 mm×250 mm,5 μm);流動相:A相(甲醇)、B相(乙腈-水-三氟乙酸(60∶40∶0.04,V/V));洗脫程序:A相加1%洗脫液B相洗5 min,然后B相由1%逐步上升至100%,洗脫25 min;流速:0.9 mL/min;進(jìn)樣量:20 μL;柱溫:40 ℃;檢測波長:214 nm。
樣品測定:利用配自動進(jìn)樣器和紫外檢測器的Agilent 1100型高效液相色譜儀對樣品肽含量進(jìn)行測定。
1.4 數(shù)據(jù)處理
數(shù)據(jù)采用SPSS 13.0統(tǒng)計(jì)軟件包進(jìn)行分析處理,并采用Duncan’s方法對各處理進(jìn)行均值的多重比較分析。
2 結(jié)果與分析
2.1 發(fā)酵牦牛肉灌腸過程中pH值的變化
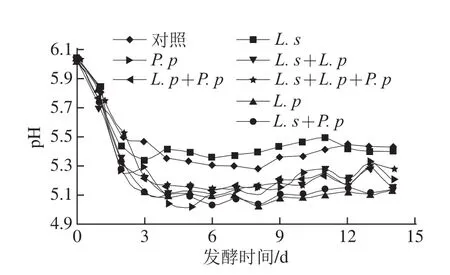
圖1 牦牛肉發(fā)酵灌腸在發(fā)酵過程中pH值變化Fig.1 Changes in pH value during the fermentation process of yak meat sausage
由圖1可知,在整個(gè)發(fā)酵過程中各個(gè)處理之間pH值均先在主發(fā)酵階段快速下降,此階段接種了乳酸菌灌腸的pH值下降速率大于對照組;其后在自然發(fā)酵過程中pH值均緩慢繼續(xù)下降到一定值后趨于穩(wěn)定,在此階段接種了L. s處理組的pH值穩(wěn)定于5.30~5.40,明顯高于其他處理;接種其他乳酸菌處理組的pH值均小于5.30,明顯小于對照組。乳酸菌能將牦牛肉灌腸中的碳水化合物快速降解為乳酸,致使處理組中的pH值快速下降;L. s含有大量的中性活性蛋白酶[18],將肉中蛋白快速分解生成堿性物質(zhì),使接種了L. s處理組的pH值明顯高于其他處理組。
2.2 發(fā)酵牦牛肉灌腸過程中總肽含量及其變化
國內(nèi)外多名學(xué)者[19-22]研究表明:乳酸菌促進(jìn)發(fā)酵肉制品中肽的生成主要有兩種途徑:1)通過乳酸菌發(fā)酵作用,產(chǎn)生乳酸降低pH值,激活肌肉本身組織蛋白酶的活性,加速肌肉蛋白的降解,促使牦牛肉灌腸產(chǎn)生肽類;2)乳酸菌分泌的細(xì)胞外蛋白酶降解肌肉蛋白產(chǎn)生肽類。但乳酸菌作用于蛋白分解成肽的過程中,不同的菌種會有差異。
2.2.1 不同發(fā)酵時(shí)期牦牛肉灌腸中肽的HPLC圖
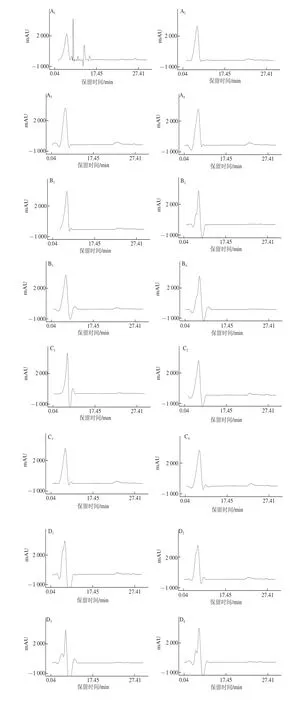
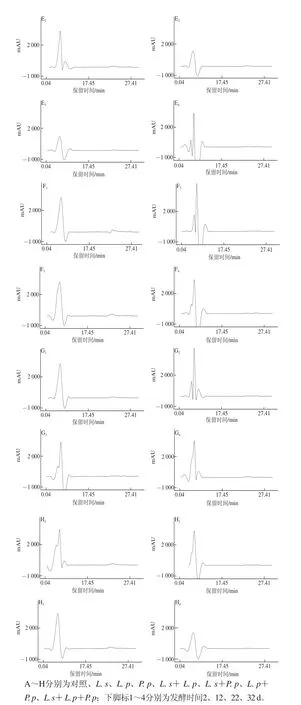
圖2 不同處理在不同發(fā)酵時(shí)間牦牛肉灌腸中肽的HPLC圖Fig.2 HPLC chromatography of peptides of yak meat sausage during different fermentation periods
由圖2可知,對照組在主發(fā)酵結(jié)束2 d時(shí)檢出了多條洗脫峰,其后在自然風(fēng)干發(fā)酵的3個(gè)時(shí)期檢出了峰形相似的洗脫峰,集中于保留時(shí)間在1~5 min,即在發(fā)酵過程中,保留時(shí)間在6~17 min,用大于0.6%乙腈參與洗脫出的相對極性較弱的肽明顯減少了,這與陸應(yīng)林等[19]在南京板鴨加工過程中發(fā)現(xiàn)非極性肽的減小相似。與對照組相比,此3 株乳酸菌及其組合在整個(gè)發(fā)酵過程中檢出肽的色譜圖與對照組在自然風(fēng)干發(fā)酵的極為相似,主要是保留時(shí)間在1~5 min,0.05%三氟乙酸洗脫出相對極性較強(qiáng)的肽,可能此3 株乳酸菌發(fā)酵加速了牦牛肉灌腸中相對極性較弱肽的降解,但機(jī)理尚不明確,有待于進(jìn)一步研究。
2.2.2 發(fā)酵牦牛肉灌腸中總肽含量及其變化
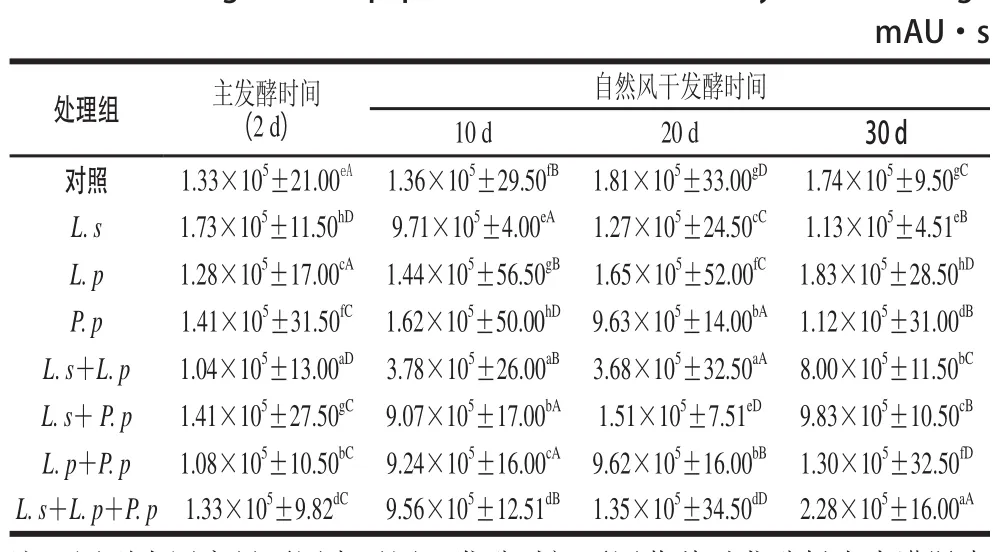
表2 發(fā)酵牦牛肉灌腸中的總肽含量及其變化Table2 Changes in total peptide content of fermented yak meat sausage
由表2可知,主發(fā)酵(32 ℃,2 d)結(jié)束時(shí),乳酸菌發(fā)酵劑對牦牛肉灌腸中總肽含量影響顯著(P≤0.05),肽含量從大到小依次為L. s>L. s+P. p>P. p>對照組>L. s+L. p+P. p>L. p>L. p+P. p>L. s+L. p。在此階段,接種了L. s的牦牛肉灌腸檢出肽的峰面積和為1.73×105mAU·s,顯著高于其他樣品,可能是因?yàn)長. s中含有較多與肌肉蛋白降解相關(guān)的中性活性酶類。
繼續(xù)自然風(fēng)干發(fā)酵10 d,即發(fā)酵12 d后,發(fā)酵劑對牦牛肉灌腸中總肽含量的影響仍顯著(P≤0.05),P. p發(fā)酵的處理組檢出最大,峰面積和為1.62×105mAU·s,其含量是此階段檢出最低總肽含量L. s+L. p處理組的42.8 倍。產(chǎn)生這一結(jié)果的可能原因是P. p快速降解牦牛肉灌腸中的碳水化合物產(chǎn)酸,降低樣品pH值,進(jìn)而激活牦牛肉灌腸中的組織蛋白酶,生成了大量的肽類[20]。
發(fā)酵22 d后,除P. p與L. p+P. p發(fā)酵處理組檢出差異不顯著(P>0.05)外,其余樣差異均顯著(P≤0.05)。牦牛肉灌腸中檢出的總肽含量的大小依次為對照組>L. p>L. s+P. p>L. s>L. s+L. p>P. p>L. p+P. p>L. s+ L. p。對照組的峰面積和(1.81×105mAU·s)是L. s+ L. p組合發(fā)酵處理組的5 倍;L. s+P. p檢出峰面積和為1.51×105mAU·s,是復(fù)配菌種在本實(shí)驗(yàn)中檢出的最大值。
發(fā)酵32 d后,乳酸菌發(fā)酵牦牛肉灌腸中檢出總肽含量依次為L. p>對照組>L. p+P. p>L. s>P. p>L. s+P. p>L. s+L. p>L. s+L. p+P. p。L. p檢測出總肽含量最多,峰面積和為1.83×105mAU·s;L. s+L. p+P. p檢測出肽含量最少,峰面積和為2.28×105mAU·s。在此階段,接種了L. p的牦牛肉灌腸中的肽含量高于其他處理。
在整個(gè)發(fā)酵過程中,不同乳酸菌發(fā)酵牦牛肉灌腸中,檢測出的總肽含量從高到低依次為L. p(1.83×105mAU·s)、對照組(1.81×105mAU·s)、L. s(1.73×105mAU·s)、P. p(1.62×105mAU·s)、L. s+P. p(1.51×105mAU·s)、L. s+L. p+P. p(1.35×105mAU·s)、L. p+P. p(1.30×105mAU·s)、L. s+L. p(1.04×105mAU·s)。
由表2可知,同一乳酸菌菌種在發(fā)酵牦牛肉灌腸的4 個(gè)不同時(shí)期,檢出發(fā)酵時(shí)間對灌腸中總肽含量的影響差異均顯著,但因發(fā)酵菌種的不同,總肽含量的變化趨勢有差異。
單一乳酸菌L. p發(fā)酵的處理組在整個(gè)發(fā)酵過程中檢出總肽含量基本呈現(xiàn)出上升的趨勢,與L. p極強(qiáng)的環(huán)境適應(yīng)能力和蛋白酶分泌能力有關(guān)[23]。在4 個(gè)發(fā)酵時(shí)期,L. p+L. s組合發(fā)酵的處理組和L. p+P. p組合發(fā)酵的處理組檢出總肽含量均低于L. p單一菌種的發(fā)酵,其中L. s+L. p檢出值均低于L. p+P. p。說明發(fā)酵牦牛肉灌腸過程中L. p與L. s、P. p在肽的生成作用中可能具有一定的拮抗作用,且與L. s的拮抗性強(qiáng)于P. p。
隨發(fā)酵時(shí)間的延長,對照組、L. s、L. s+P. p、L. s+L. p+P. p檢出總肽含量呈現(xiàn)出相似的趨勢,即先下降后上升再略有下降。其中對照組的變化不大,與自然微生物較強(qiáng)的環(huán)境適應(yīng)能力有關(guān),從12~22 d總肽含量略有升高是因?yàn)殛笈H夤嗄c利用環(huán)境與自身攜帶的微生物經(jīng)過12 d的發(fā)酵,分解碳水化合物產(chǎn)酸,降低pH值,激活肌肉內(nèi)源性蛋白酶,使其分解蛋白產(chǎn)生肽[19]。L. s與其參與發(fā)酵L. s+P. p和L. s+L. p+P. p呈現(xiàn)出相似的變化,在發(fā)酵第2天、第22天均檢出了較高的總肽含量,與L. s中含有大量的肽酶有直接的關(guān)系[24]。在發(fā)酵第12天、第32天均檢出較低的總肽含量,可能原因主要有3 個(gè):1)由L. s從適宜的主發(fā)酵環(huán)境到自然環(huán)境的不適應(yīng)引起;2)與環(huán)境中其他腐敗微生物對肽的降解有關(guān);3)低pH值加速了水分的蒸發(fā),快速降低了樣品的水分活度,造成了L. s的不良生存環(huán)境[25]。但偏重程度不同,12 d的偏于前一個(gè)原因,32 d的主要由后兩個(gè)原因引起。L. s+P. p從發(fā)酵第2天到第12天降低幅度小于L. s,從12~22 d范圍增加幅度大于L. s,并在22 d檢出的總肽含量顯著高于由單一乳酸菌發(fā)酵L. s和P. p發(fā)酵的處理組,32 d檢出值略小于由兩個(gè)單一菌種發(fā)酵的處理組,L. s與P. p同時(shí)作用于發(fā)酵牦牛肉灌腸過程中對肽的生成可能具有一定的協(xié)同作用。
在發(fā)酵牦牛肉灌腸中,檢出總肽含量先上升后下降再略有所上升的是P. p,從主發(fā)酵到自然風(fēng)干發(fā)酵12 d含量上升,說明P. p具有一定的環(huán)境適應(yīng)能力和蛋白的分解能力[26]。隨發(fā)酵時(shí)間的延長,牦牛肉灌腸水分活度的降低,可能是P. p適應(yīng)低水分活度的環(huán)境能力較差,加之,肽類自身的易消耗性,導(dǎo)致了從12~22 d急劇的下降。
在3 株乳酸菌發(fā)酵牦牛肉灌腸過程中,發(fā)酵菌種和發(fā)酵時(shí)間均是影響灌腸中總肽含量變化的顯著性因素,主要通過分泌蛋白酶或降低pH值激活組織蛋白酶使肌肉蛋白降解成肽。Fadda等[27]采用超濾、RP-HPLC、基質(zhì)輔助激光解吸電離飛行時(shí)間質(zhì)譜測序等技術(shù),對市場品質(zhì)優(yōu)良的5 種阿根廷干發(fā)酵腸中的水解酶體系進(jìn)行了研究,結(jié)果發(fā)現(xiàn),酶解后的氨基酸組成極其復(fù)雜,間接反映出了水解體系的復(fù)雜性,所以乳酸菌酶系對發(fā)酵肉制品蛋白降解的具體作用機(jī)制尚不明確,有待于進(jìn)一步的研究。
3 結(jié) 論
乳酸菌發(fā)酵劑菌種、發(fā)酵時(shí)間對發(fā)酵牦牛肉灌腸過程中肽的變化規(guī)律均有顯著影響。隨著發(fā)酵時(shí)間的延長,發(fā)酵牦牛肉灌腸中的肽發(fā)生了明顯的變化,相對極性較弱的肽減小了,相對極性較強(qiáng)的肽集中在保留時(shí)間為1~5 min。在32 ℃條件下發(fā)酵2 d后,L. s發(fā)酵處理組中檢出了最大的總肽含量,且其參與發(fā)酵的L. s+P. p和L. s+L. p+P. p組合中檢出總肽含量的變化趨勢與單一L. s發(fā)酵處理組相似,L. s是牦牛肉灌腸中快速生成肽的良好發(fā)酵劑;在4 個(gè)發(fā)酵時(shí)期,L. p+L. s組合發(fā)酵處理組檢出的總肽含量均小于單一L. p與L. s發(fā)酵處理組,L. p與L. s在同時(shí)作用于發(fā)酵牦牛肉灌腸中肽的生成作用中,兩者之間具有一定的拮抗作用;L. s與P. p組合在降解牦牛肉蛋白生成肽的整個(gè)發(fā)酵過程中,產(chǎn)生較多的肽,也能迅速降低發(fā)酵牦牛肉灌腸pH值,保證產(chǎn)品的安全性,為開發(fā)生產(chǎn)高營養(yǎng)、安全性能好等優(yōu)質(zhì)牦牛肉制品提供了新技術(shù)方案。
參考文獻(xiàn):
[1] 萬紅玲, 雒林通, 吳建平. 牦牛肉品質(zhì)特性研究進(jìn)展[J]. 畜牧獸醫(yī)雜志, 2012, 31(1): 36-40.
[2] 李平蘭, 王成濤. 發(fā)酵食品安全生產(chǎn)與品質(zhì)控制[M]. 北京: 化學(xué)工業(yè)出版社, 2005: 50-55.
[3] 劉曉蓉, 鄧毛程, 連曉蔚. 腌肉中乳酸菌的分離選育[J]. 中國釀造, 2009, 28(1): 46-48.
[4] 何煜波. 乳桿菌屬發(fā)酵過程中變化的研究[J]. 肉類工業(yè), 2002(5): 23-28.
[5] 許慧卿, 蔣云升, 王畏畏, 等. 米酒乳桿菌對風(fēng)鴨肌肉蛋白質(zhì)降解的影響[J]. 食品與發(fā)酵工業(yè), 2008, 34(2): 145-148.
[6] 徐幸蓮, 徐為民, 周光宏, 等. 植物乳桿菌6003對肌肉蛋白質(zhì)的分解能力(Ⅰ)[J]. 食品科學(xué), 2003, 24(2): 56-61.
[7] RIZZELLO C G, NIONELLI L, CODA R, et al. Synthesis of the cancer preventive peptide lunasin by lactic acid bacteria during sourdough fermentation[J]. Nutrition & Cancer, 2012, 64(1)∶ 111-120.
[8] 吳少輝, 張成桂, 劉光明. 高效液相色譜法在蛋白質(zhì)分離檢測中的應(yīng)用[J]. 畜牧與飼料科學(xué), 2011, 32(8): 63-65.
[9] HERN?NDEZ-LEDESMA B, HSIEH C C, de LUMEN B O. Lunasin, a novel seed peptide for cancer prevention[J]. Peptides, 2009, 30(2)∶426-430.
[10] CAVAZOS A, de MEJIA E G. Identification of bioactive peptides from cereal Storage proteins and their potential role in prevention of chronic diseases[J]. Comprehensive Reviews in Food Science and Food Safety, 2013, 12(4)∶ 364-380.
[11] PINS J J, KEENAN J M. Effects of whey peptides on cardiovascular disease risk factors[J]. The Journal of Clinical Hypertension, 2006, 8(11)∶ 775-782.
[12] GIESSEN T W, MARAHIEL M A. Ribosome-independent biosynthesis of biologically active peptides∶ application of synthetic biology to generate structural diversity[J]. FEBS Letters, 2012, 586(15)∶ 2065-2075.
[13] 蔣云升, 汪志君, 于海, 等. 如式香腸用發(fā)酵劑多菌種混培養(yǎng)技術(shù)的研究[J]. 食品科學(xué), 2008, 29(8): 410-413.
[14] 王永霞, 牛天貴. 肉品混合發(fā)酵劑的篩選及應(yīng)用研究[J]. 食品科技, 2004, 29(8): 34-38.
[15] SILVINA F, YOLANDA S, GRACIELA V, et al. Hydrolysis of pork muscle sarcoplasmic proteins by lactobacillus curvatus and lactobacillus sake[J]. Applied and Environmental Microbiology, 1999, 65(2): 578-584.
[16] MOYA V J, FLORES M, ARISTOY M C, et al. Evolution of hydrophobic polypeptides during the ageing of exudative and nonexudative pork meat[J]. Meat Science, 2001, 57(4): 395-401.
[17] FADDA S, SANZ Y, VIGNOLO G, et al. Characterization of muscle sarcoplasmic and myofibrillar protein hydrolysis caused by Lactobacillus plantarum[J]. Applied and Environmental Microbiology, 1999, 65(8): 3540-3546.
[18] 陳倩, 韓齊, 孔保華, 等. 乳酸菌對發(fā)酵肉制品中肌肉蛋白降解作用的研究進(jìn)展[J]. 食品科學(xué), 2014, 35(9): 279-284. doi: 10.7506/ spkx1002-6630-201409055.
[19] 陸應(yīng)林, 周光宏, 徐幸蓮, 等. 南京板鴨加工過程中肌肉蛋白降解的研究[J]. 食品科學(xué), 2006, 27(7): 44-47.
[20] 陸應(yīng)林. 南京板鴨加工過程中蛋白降解及風(fēng)味物質(zhì)的研究[D]. 南京: 南京農(nóng)業(yè)大學(xué), 2012.
[21] 白風(fēng)翎, 張柏林, 趙宏飛. 乳酸菌蛋白代謝研究進(jìn)展[J]. 食品科學(xué), 2010, 31(19): 381-384.
[22] SAVIJOKI K, INGMER H, VARMANEN P. Proteolytic systems of lactic acid bacteria[J]. Applied Microbiology and Biotechnology, 2006, 71(4)∶ 394-406.
[23] 陳臣, 任婧, 周方方, 等. 植物乳桿菌的比較基因組學(xué)研究[J]. 中國生物工程雜志, 2013, 33(12): 35-44.
[24] MOLLY K, DEMEYER D, JOHANSSON G, et al. The importance of meat enzymes in ripening and fl avour generation in dry fermented sausages. First results of a European project[J]. Food Chemistry, 1997, 59(4): 539-545.
[25] 謝愛英, 張富新, 陳穎. 發(fā)酵香腸的pH值、水分含量與水分活度(Aw)的關(guān)系及其對制品貯藏性的影響[J]. 食品與發(fā)酵工業(yè), 2004, 30(11): 143-146.
[26] 戴夢婕, 許艷順, 姜啟興, 等. 戊糖片球菌產(chǎn)蛋白酶對發(fā)酵 鰱魚魚糜凝膠性能的影響[J]. 食品與發(fā)酵工業(yè), 2014, 40(6): 17-21.
[27] FADDA S, LóPEZ C, VIGNOLO G. Role of lactic acid bacteria during meat conditioning and fermentation peptides generated as sensorial and hygienic biomarkers[J]. Meat Science, 2010, 86(1): 66-79.
Effects of Three Strains of Lactic Acid Bacteria and Their Combinations on the Peptide Changes in Fermented Yak Meat Sausage
BAI Juhong, TANG Shanhu*, LI Sining, WANG Liu, LU Fuqing, YAN Liguo
(College of Life Science and Technology, Southwest University for Nationalities, Chengdu 610041, China)
This study has explored the effects of different starter cultures including three strains of lactic acid bacteria and their combinations on the peptide changes of fermented yak meat sausage.Lactobacillus sake(L. s),Lactobacillus plantarum(L. p),Pediococcus pentosaceus(P. p) and their combination were artifi cially inoculated for the fermentation of yak meat sausage. pH values were determined at intervals of 24 hours. Samples were collected after 2 days of primary fermentation (32 ℃) and after 10, 20, and 30 days of natural air-dry fermentation. Peptide content of yak meat sausages was analyzed by reversed phase high performance liquid chromatography (RP-HPLC). The results showed that all different lactic acid bacteria and their combinations exceptP. pandL. p+P. phad signifi cant effects on the peptide contents of yak meat sausage after 2 days of primary fermentation and after 10, 20, and 30 days of natural air dry fermentation (P< 0.05). The highest peptide contents of 1.73 × 105, 1.62 × 105, 1.81 × 105and 1.83 × 105mAU·s in yak meat sausages fermented withL. s,P. p, control (no starter culture) andL. p,respectively, were detected at the above four fermentation times sequentially. With the extension of fermentation time, notable changes were observed in the peptide content of fermented yak meat sausage. The content of peptides with relatively weak polarity was reduced, whereas peptides with relatively strong polarity largely remained unchanged. At the end of primary fermentation, theL.sgroup showed the maximum total peptide content, and the changes in the total peptide contents of theL. s,L. s+P. p,L. s + L. p + P. pgroups presented similar trends.L. sfermentation was able to quickly generate peptides in yak meat sausage. At the four fermentation periods, the peptide content of theL. p+L. sgroup were lower than that of the single strain groups, andL. p + L. seven showed inhibition on the production of peptides in yak meat sausage.L. sandP. pdemonstrated no observable differences in peptide production, but could quickly reduce the pH value, ensuring the safety of the product. These results may provide useful information for the commercial production of yak meat sausage with high quality.
lactic acid bacteria; fermentation; yak meat sausage; peptides; reversed phase high perf ormance liquid chromatography (RP-HPLC)
TS251.1
A
10.7506/spkx1002-6630-201517035
2014-11-26
西南民族大學(xué)2014年研究生“創(chuàng)新型科研項(xiàng)目”碩士重點(diǎn)項(xiàng)目(CX2014SZ83);四川省科技支撐計(jì)劃項(xiàng)目(2014NZ0052)
白菊紅(1989—),女,碩士研究生,研究方向?yàn)樾螽a(chǎn)品加工與安全。E-mail:18215526804@163.com
*通信作者:唐善虎(1964—),男,教授,博士,研究方向?yàn)槭称房茖W(xué)與工程。E-mail:stang01@126.com

- 食品科學(xué)的其它文章
- “互聯(lián)網(wǎng)+”潮中百舸爭流,調(diào)味品業(yè)內(nèi)究竟何為?
——2015第十一屆調(diào)味品博覽會營銷峰會11月下旬將在上海召開 - 第二屆中國食品科技成果交流會暨第五屆中國食品產(chǎn)業(yè)產(chǎn)學(xué)研創(chuàng)新發(fā)展高峰論壇
- 我國水產(chǎn)品安全風(fēng)險(xiǎn)來源與風(fēng)險(xiǎn)評估研究進(jìn)展
- 食源性蛋白中免疫活性肽的研究進(jìn)展
- 轉(zhuǎn)基因組學(xué)分析技術(shù)研究進(jìn)展
- 多酚類化合物對糖脂代謝影響的研究進(jìn)展