非酵母屬酵母的接種順序對混合發酵菠蘿酒香氣成分的影響
2015-01-03 01:14:55賈言言劉四新李卓婷王艷梅易錦豪丁俊權李從發
食品科學 2015年17期
賈言言,劉四新,李卓婷,王艷梅,易錦豪,丁俊權,李從發*
非酵母屬酵母的接種順序對混合發酵菠蘿酒香氣成分的影響
賈言言,劉四新,李卓婷,王艷梅,易錦豪,丁俊權,李從發*
(海南大學食品學院,海南 海口 5702 28)
采用非酵母屬的梅奇酵母P3-3與酵母屬的釀酒酵母D254以不同接種順序混合發酵菠蘿酒。結果表明:頂空固相微萃取-氣質聯用法檢測菠蘿原果汁中共有20種香氣成分,而先接種P3-3培養7 d后再接種D254(SI-Ⅰ組)、先接種D254培養2 d再接種P3-3(SI-Ⅱ組)和二者同時接種(CI組)的菠蘿酒中則分別檢出53、44、41種香氣成分,SI-Ⅰ組所釀菠蘿酒風味物質種類最多;SI-Ⅰ組中羧酸酯類相對含量為15.594%,而CI組和SI-Ⅱ組分別為13.042%和6.655%,表明SI-Ⅰ組所合成的羧酸酯類物質較多;菠蘿特征香氣成分的菠蘿甲酯、菠蘿乙酯、丁內酯等的含量在SI-Ⅰ組中為1.337%,CI組為0.844%,SI-Ⅱ組為0.819%,說明SI-Ⅰ組能更好地保留菠蘿原果香;最后,SI-Ⅰ組感官品質最好。可見,采用先接種非酵母屬酵母P3-3使其生長達到旺盛生長期,再接種發酵 力強的酵母屬酵母D254的混合發酵方式,能更好地保留果酒中原果香的典型特征,增加菠蘿酒香氣成分的種類,并有效改善菠蘿酒的品質、風味。
非酵母屬酵母;梅奇酵母;混合發酵;菠蘿酒;香氣成分
菠蘿(Ananas comosus(L.)Merr.)又名鳳梨,鳳梨科果子蔓屬,單子葉多年生草本植物,是世界熱帶、亞熱帶特產水果之一,其中海南、廣東、廣西是我國菠蘿生產主要產區[1]。成熟的菠蘿果實中含有大量的糖、酸和芳香性物質,因此有大量的研究報道將菠蘿釀制成果酒。但實踐證明,經過發酵以后菠蘿的原果香損失較大、香味組成發生了變化,導致菠蘿酒的菠蘿風味特征不典型、不豐滿,酒香也單薄。因此,如何保香、增香是菠蘿酒釀制研中的難度、重點。
果酒釀造中由于要得到一定酒精體積分數的產品,一般都使用發酵力強的酵母屬酵母,如釀酒酵母(Saccharomyces cerevisiae)、貝酵母(Saccharomyces bayanus)等,但其在產香和香氣形成方面一般比較單薄,風味欠佳。因此近年有不少研究采用非酵母屬酵母進行發酵,它 們通常是一些篩自各種水果及其土壤環境、產香能力較好的非酵母屬酵母,如梅氏酵母(Metschnikowia)、假絲酵母(Candida)、有孢圓酵母(Torulaspora)、Lachancea、克魯維酵母(Kluyveromyces)、漢遜酵母(Hanseniaspora)和結合酵母(Zygosaccharomyces)等[2-4]。雖然使用非酵母屬酵母具有很多對果酒品質有利的影響,但也存在一些缺陷,比如耐酒精能力弱、發酵力低、醋酸產量高等。因此,采用酵母屬酵母與非酵母屬酵母進行混合發酵[5-6],是當前的一個研究熱點。Panjai等[7]對比了戴爾凱氏有孢圓酵母(T. delbrueckii)和釀酒酵母(S. cerevisiae)混合發酵和單一發酵的菠蘿酒,結果表明,混合發酵的菠蘿酒具有更明顯的菠蘿風味特征。然而,當非酵母屬酵母與酵母屬酵母混合發酵時,往往因為其不耐酒精、對營養物質的低競爭力等原因[8-11],會在接種的3~4 d后迅速大量衰亡[12-13],而其過早衰亡對果酒香氣形成的貢獻就很有限,因此,本實驗為了延長非釀酒屬酵母的生長時間,在接種順序上進行了對比研究,將本課題組前期從番木瓜果皮上篩選到的一株產香良好的梅奇酵母屬(Metschnikowia agaves)P3-3菌株與酵母屬的釀酒酵母(Saccharomyces cerevisiae)D254菌株進行混合發酵釀制菠蘿酒,以期為高品質菠蘿酒的研發提供參考。
1 材料與方法
1.1材料、菌種、培養基與試劑
濃縮菠蘿原汁:巴厘種,由海南南派實業有限公司提供。
梅奇酵母(Metschnikowia agaves)P3-3,分離、鑒定自番木瓜果皮,保存于海南大學食品學院菌種保藏中心;釀酒酵母(Saccharomyces cerevisiae)D254購自上海杰兔工貿有限公司。
WL培養基與LY培養基 青島日水生物技術有限公司;YEPD培養基:酵母浸粉10 g、蛋白胨20 g、葡萄糖20 g、瓊脂20 g、蒸餾水1 000 mL,pH 6.0,115℃濕熱滅菌25 min。
果膠酶 煙臺曼森商貿有限公司;2-辛醇 阿拉丁試劑有限公司;其他試劑均為國產分析純。
1.2儀器與設備
HP6890/5973N型氣相色譜-質譜(gas chromatography mass spectrometry,GC-MS)聯用儀 美國Agilent公司;萃取頭(CAR/PDMS,75 μm) 美國Supelco公司;Astree2系統電子舌 法國Alpha M.O.S公司;Biofugestratos高速冷凍離心機 德國Thermo公司。
1.3方法
1.3.1菠蘿酒釀造工藝流程
濃縮菠蘿原汁→稀釋→果膠酶處理→離心→SO2處理→調整糖、酸度→接種酵母菌→主發酵→后發酵→貯存陳釀→分析、鑒評
1.3.2常規理化成分分析
總糖含量測定:3,5-二硝基水楊酸(3,5-dinitrosalicylic acid,DNS)比色法[14];可溶性固形物含量測定:手持糖度計法;總酸含量、揮發酸含量、酒精體積分數測定:參照GB/T 15038—2006《葡萄酒、果酒通用分析方法》進行測定。
1.3.3電子舌分析
電子舌系統:法國Alpha M.O.S公司生產的Astree2電子舌系統,此電子舌系統包含7個化學傳感器陣列代號分別為ZZ、BB、CA、GA、HA、JB、JE。這7個傳感器對5種基本味覺:酸、甜、苦、咸與鮮都有不同的響應。
電子舌實驗方法:取樣,每種樣品量取80 mL,靜置15 min。每次采樣時間120 s,每秒采樣一次,電子舌系統軟件每秒鐘自動記錄一次數據。每個樣品重復4次。
樣品的統計學分析:利用電子舌自帶的Alphasoft 12.3統計分析軟件,主成分分析(principal component analysis,PCA)和判別因子分析(discriminatory factor analysis,DFA)進行數據處理,對樣品進行定性判別。
1.3.4 GC-MS分析
萃取方法:將固相微萃取(s o l i d-p h a s e microextraction,SPME)萃取頭在GC-MS儀的進樣口老化,老化溫度250℃,老化時間30 min,載氣流速1.0 mL/min。準確稱取10.000 g樣品于15 mL的頂空瓶中,10 ?L 2-辛醇(250 mL/L)為內標,密封,水浴溫度45℃,插入經過老化處理的固相微萃取頭,使萃取頭處于樣品之上2 cm,萃取30 min后取出,迅速插入GC-MS聯用儀的進樣口,于250℃條件下解吸5 min后進行GC-MS分析。
GC-MS分析條件:色譜條件:毛細管柱為DB-5。氦氣為載氣,恒定流速1.0 mL/min,進樣口溫度250℃。程序升溫:40℃保持6 min,以3℃/min升至120℃,再以4℃/min升至230℃。
質譜檢測條件:電子轟擊離子源(EI),電子能量70 eV,質量掃描范圍m/z30~450,采集方式為全掃描scan,溶劑延遲3 min。
定性定量分析:分析結果運用計算機譜庫(NIST/ WILEY)進行初步檢索及資料分析,并結合已有的相關文獻進行定性分析。樣品定量可根據已知濃度的內標物2-辛醇進行定量分析,各物質的含量采用峰面積法計算出,即半定量的方法。
1.3.5感官評定
對不同處理的菠蘿酒,經1個月陳釀后,請8名評審員參照GB/T 15038—2006進行感官鑒評,并測定酒精體積分數。
1.3.6菠蘿酒的混合發酵
菌種活化:非酵母屬的P3-3:將斜面接入盛有5 mL豆芽汁的試管中,25℃振蕩培養10 h,然后轉入盛有菠蘿汁的三 角瓶中繼續擴大培養10 h; 酵母屬的D254(活性干酵母):稱取1 g加入到含5%葡萄糖的10 mL水溶液中,40℃活化30 min。為便于后續接種時計算菌體濃度,先接入YEPD斜面培養,然后同P3-3一樣進行活化培養。
發酵操作:濃縮菠蘿原果汁加水稀釋至20 °Brix,40 ℃條件下果膠酶處理40 min,10 000×g離心10 min,取上清液以檸檬酸調整pH值為3.5,加200 mg/L偏重亞硫酸鈉滅菌6 h備用。然后以不同順序接種P3-3與D254進行菠蘿酒的混合發酵,25 ℃主發酵20 d,15 ℃后發酵1 個月,然后陳釀1 個月。
接種方式:將兩種酵母按兩種順序依次接種,并與同時接種進行對照。
將P3-3與D254分別以1×107、1×105CFU/mL同時接入處理過的菠蘿汁中,進行混合發酵(同時接種組,co-inoculating,CI),作為對照;先接入濃度為1×107CFU/mL的P3-3培養7 d,以達 到其旺盛生長期,后接入濃度為1×105CFU/mL的D254進行混合發酵(依次接種Ⅰ組,successively-inoculatingⅠ,SI-Ⅰ)。先接入濃度為1×105CFU/mL的D254培養2 d,以達到其旺盛生長期,后接入濃度為1×107CFU/mL的P3-3進行混合發酵(依次接種Ⅱ組,SI-Ⅱ)。
2 結果與分析
2.1不同接種方式的發酵體系中酵母菌細胞濃度的變化
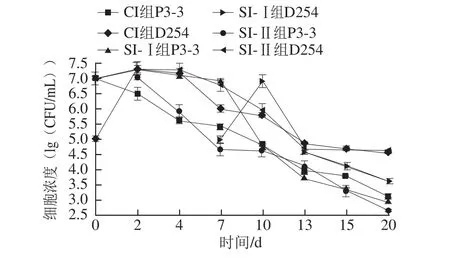
圖1 菠蘿酒混合發酵中酵母菌細胞濃度的變化Fig.1 Effect of fermentation time on cell concentrations of yeasts in pineapple wine
根據前期單菌株發酵的實驗,梅奇酵母P3-3需要達到107~108CFU/mL數量級才啟動發酵,以及為防止D254的旺盛繁殖帶走過多香氣成分,故分別選擇了1×107CFU/mL和1×105CFU/mL兩種接種濃度。計數結果(圖1)可看出,同時接種的CI組與先接種D254的SI-Ⅱ組的P3-3與D254的生長規律相似,二者中D254細胞都在前2 d迅速增殖,而P3-3都呈現活細胞數迅速下降的趨勢,這與Soden[15]和Toro[16]等研究結論相似,說明在強發酵力D254的環境中,非酵母屬酵母的生長勢確實很弱;而先接種P3-3的SI-Ⅰ組中,P3-3能在第2天細胞濃度達到峰值,之后較穩定生長,稍有下降,直到第7天接入D254時P3-3細胞迅速衰亡。這可能與混合發酵體系中營養物質的競爭效應、有毒物質(如乙醇)的形成濃度、以及細胞的群體感應等因素有關,這從生物量的角度說明混合發酵時以非酵母屬酵母先接種、培養一段時間后再接入發酵力強的酵母屬酵母的方式是非常必要的。
2.2常規理化分析

圖2 菠蘿酒發酵過程中各常規成分的變化Fig.2 Effect of fermentation time on chemical indicators of pineapple wine
常規理化成分(圖2)分析結果顯示,SI-Ⅱ組與CI組的變化相似,而SI-Ⅰ組中可溶性固形物、總糖含量下降緩慢,這說明P3-3作為野生酵母,其代謝能力顯然低于商業酵母D254;產酸方面,發酵初期,SI-Ⅰ組總酸下降較慢,說明其代謝菠蘿果汁中的有機酸能力也較弱,加之一般非酵母屬酵母的產醋酸能力較強[17],因此發酵終了時SI-Ⅰ組總酸含量高于CI、SI-Ⅱ組。但值得關注的是,其揮發酸含量在SI-Ⅰ組卻最低,這可能與先接種P3-3、使其生長時間有效延長后,能在后期混合發酵和陳釀過程中將部分乙酸轉化為具有芳香味的低級醇有關[18]。
2.3電子舌分析
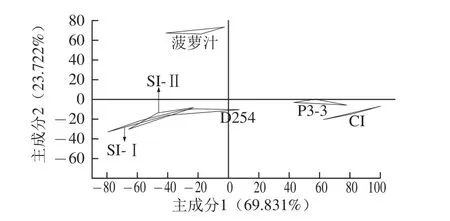
圖3 菠蘿汁和菠蘿酒的主成分分析Fig.3 Principal component analysis of pineapple juice and wine
為了比較不同接種方式的混合發酵的菠蘿酒之間的口味差異,進行了電子舌分析(圖3),并引入菠蘿汁和純種接種的單一發酵的菠蘿酒做對照。同一樣品在主成分圖上由3個點組成,得到前2個主成分的貢獻率分別為69.831%、23.722%,累計貢獻率93.553%,說明足以收集特征性信息[19]。菠蘿汁、單一接種P3-3組與CI組的酒樣都能夠很好地落在各自的區域里沒有重疊,說明它們存在的差異能給區分開來。而SI-Ⅰ與SI-Ⅱ、SI-Ⅱ與單一接種D254組的響應數據圖有交叉,說明兩兩之間口味差異不大,但結合感官評定情結果,表明電子舌沒能很好地區分它們的差異。然而另一方面,SI-Ⅰ組、SI-Ⅱ組與單一接種D254組三者間有一部分重疊,表示D254產生的風味在SI-Ⅰ組、SI-Ⅱ組都有所體現。因此為了將CI組、SI-Ⅰ組、SI-Ⅱ組的口味差異區分出來,需對其進行進一步的判別因子分析。
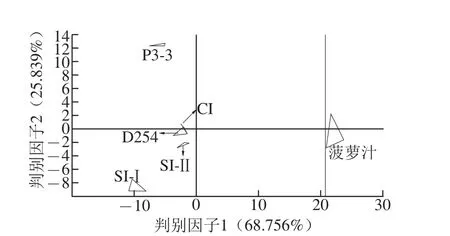
圖4 菠蘿汁和菠蘿酒的判別因子分析Fig.4 Discriminant factor analysis of pineapple juice and wine
將電子舌采集得到的數據進行判別因子分析(圖4)可知,判別因子分 析對樣品的累積區分指數達到了94.595%,同一樣品組內的離散度也比主成分分析(圖3)的小,不同樣品組間的區分更明顯,能夠對樣品間差異進行區分。而在主成分分析中不能明顯區分的SI-Ⅰ組、CI組、SI-Ⅱ組,判別因子分析能將這3種菠蘿酒很好地區分出來,說明三者的口味之間有顯著差異,此差異可能源自非酵母屬酵母P3-3與商業酵母D254的不同接種順序。
2.4香氣成分分析

表1 不同接種順序發酵的菠蘿酒中揮發性化合物的組成及含量Table1 Concentrations of the volatile compounds in pineapple wine fermented with mixed and sequential cultures of M. agaves and S. cerevisiae

續表1
菠蘿酒香氣成分中有些成分是菠蘿汁中本身存在的,有些則在發酵中產生或者后酵中形成的,這與發酵醪中微生物各種酶的活力及其代謝中間物種類等有關[20]。由氣質聯用檢測結果顯示,菠蘿汁中檢測出了20種香氣成分,而SI-Ⅰ組、CI組與SI-Ⅱ組則分別檢出53、44、41種。由此可知,混合發酵的菠蘿酒香氣成分的種類增加了[21-22]。而且,SI-Ⅰ組中香氣成分種類最多,說明先接種P3-3的SI-Ⅰ組,由于顯著延長了其生長時間,對混合發酵釀制菠蘿酒的香氣成分的復雜多樣性產生了有益效應。
酯類物 質往往是果酒中水果味與花香味的重要貢獻者。事實上果酒中的酯類一方面來自釀酒所用水果原料,另一方面來自發酵過程中的各種酯化反應。菠蘿甲酯、菠蘿乙酯、丁內酯等是菠蘿特征香氣的重要組成部分,經測定,酯類在菠蘿汁中的相對含量為0.869%,而在SI-Ⅰ組為1.337%,在CI組為0.844%,SI-Ⅱ組為0.819%,可見,SI-Ⅰ組更能保留菠蘿本身的原果香。此外,SI-Ⅰ組中的羧酸酯類相對含量為15.594%,而CI組(13.042%)與SI-Ⅱ組(6.655%)則明顯較低。這也表明先接種P3-3組的菠蘿酒通過合成重要羧酸酯類尤其是乙酸乙酯(10.480%)、乙酸苯乙酯(3.595%)等增加了酒體中的果香味與其他風味物質的復雜性。羧酸酯類一般來自于乙醇乙酰轉移酶催化CoA與醇類反應而生成[23],關于這點,Fukuda等[24]曾證明羧酸酯類含量的變化與菌體細胞內酯酶與乙醇乙酰轉移酶的合成和分解間的平衡有關。
酸類也是果酒中重要的組成部分,本次實驗檢測出的酸類有乙酸、3-甲基丁酸、己酸等。CI組酸類的相對含量為11.270%、SI-Ⅰ組為1.696%、SI-Ⅱ組為14.750%,SI-Ⅰ組中酸類物質含量較少,這可能因為P3-3利用大量的乙酰CoA合成羧酸酯類,導致乙酰CoA減少,而不足以形成乙酸等酸類,還可能是P3-3將部分乙酸轉化為具有芳香味的低級醇[18]有關。除酯類和酸類外,醇類也是果酒揮發性成分的重要組成部分。SI-Ⅰ組、CI組、SI-Ⅱ組共有的主要醇類為苯乙醇、3-甲基丁醇、2-甲基丁醇等。SI-Ⅰ組中3-甲基丁醇、苯乙醇相對含量比CI組、SI-Ⅱ組中的高,苯乙醇具有愉快的果香,對菠蘿酒有積極影響。其他揮發性物質如醛類、酮類、酚類等,由于在SI-Ⅰ組、CI組、SI-Ⅱ組中總體含量都不高,故其相對含量差異不大,但這些物質與酯類、酸類、醇類相互協同配合,共同賦予菠蘿酒清醇香韻、酒香飽滿、果香典型圓潤的風格。
2.5酒精體積分數和感官評定結果
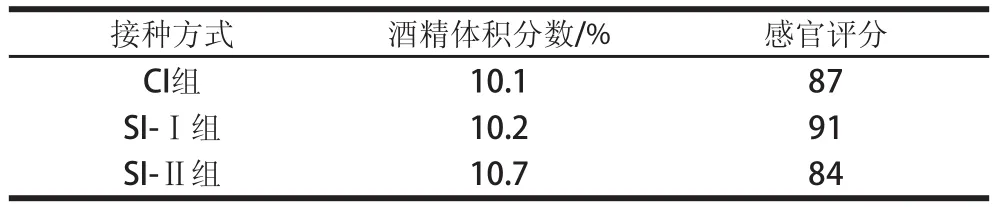
表2 菠蘿酒酒精體積分數和感官評分結果Table2 Alcohol concentrations and sensory evaluation of pineapple wines
對不同接種方式混合發酵釀制的菠蘿酒進行酒精體積分數測定和感官評價,結果見表2。CI組、SI-Ⅰ組、SI-Ⅱ組酒精體積分數差異不大,但感官綜合得分以先接種非酵母屬酵母的SI-Ⅰ組最高,其果酒色澤澄清透明、香氣圓潤、菠蘿香氣特征更突出、酒體豐滿,較好地體現了菠蘿酒的獨特風格。
3 結 論
本實驗研究了非酵母屬酵母在與酵母屬酵母混合發酵釀制菠蘿酒中二者的接種順序對果酒品質的影響,選擇使用了一株本實驗室篩選自番木瓜果皮的野生酵母Metschnikowia agavesP3-3與商業安琪酵母D254以不同接種方式進行混合發酵。SPME-GC/MS表明,菠蘿汁中檢測出20種香氣成分,而SI-Ⅰ組、CI組與SI-Ⅱ組則分別為53、44、41種,混合發酵的菠蘿酒中的香氣成分的種類增加了,且以SI-Ⅰ組中香氣成分種類最多;SI-Ⅰ組中乙酸乙酯、乙酸苯乙酯等羧酸酯類相對含量為15.594%,而CI組和SI-Ⅱ組都顯著 低于SI-Ⅰ組,分別為13.042%和6.655%,表明P3-3合成羧酸酯類的能力比較強;菠蘿特征香氣的重要組成成分菠蘿甲酯、菠蘿乙酯、丁內酯等脂類物質在菠蘿汁中相對含量為0.869%,而SI-Ⅰ組為1.337%,CI組為0.844%,SI-Ⅱ組為0.819%,說明SI-Ⅰ組能更好保留菠蘿原果香;電子舌分析表明SI-Ⅰ組、CI組、SI-Ⅱ組3種菠蘿酒口味之間有顯著差異;先接種P3-3發酵7 d后接入D254的SI-Ⅰ組由于有效延長了P3-3培養時間,使其產生了更豐富的代謝產物,結果所釀菠蘿酒風味物質多樣性最好、感官得分最高。可見,采用先接種非酵母屬酵母達到其對數期,再接種發酵力強的酵母進行混合發酵,能有效保留酒中原果香典型特征,豐富菠蘿酒香氣成分的多樣性,改善菠蘿酒的品質、風味。本實驗將梅奇酵母屬的M. agaves作為非酵母屬酵母運用于菠蘿酒的釀造中,取得良好效果,為高品質菠蘿酒的研發提供了有益借鑒。
[1] 龐社霞. 菠蘿果酒制作工藝初步探索[J]. 農產品加工∶ 學刊, 2014(8)∶ 43-45.
[2] CRAY J A, BELL A N W, BHAGANNA P, et al. The biology of habitat dominance; can microbes behave as weeds?[J]. Microbial Biotechnology, 2013, 6(5)∶ 453-492.
[3] JOLLY N P, VARELA C, PRETORIUS I S. Not your ordinary yeast∶non-Saccharomyces yeasts in wine production uncovered[J]. FEMS Yeast Research, 2014, 14(2)∶ 215-237.
[4] PRETORIUS I S. Tailoring wine yeast for the new millennium∶ novel approaches to the ancient art of winemaking[J]. Yeast, 2000, 16(8)∶675-729.
[5] GOBBI M, COMITINI F, DOMIZIO P, et al. Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential cofermentation∶ a strategy to enhance acidity and improve the overall qualityof wine[J]. Food Microbiology, 2013, 33(2)∶ 271-281.
[6] SUN Shuyang, GONG Hansheng, JIANG Xiaoman, et al. Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae on alcoholic fermentation behaviour and wine aroma of cherry wines[J]. Food Microbiology, 2014, 44∶ 15-23.
[7] PANJAI L, ONGTHIP K, CHOMSRI N. Complex fruit wine produced from dual culture fermentation of pineapple juice with Torulaspora delbrueckii and Saccharomyces cerevisiae[J]. Asian Journal of Food and Agro-Industry, 2009, 2(2)∶ 135-139.
[8] MARTINI A. Origin and domestication of wine yeast Saccharomyces cerevisiae[J]. Journal of Wine Research, 1993, 3(3)∶ 165-176.
[9] NISSEN P, ARNEBORG N. Characterization of early deaths of non-Saccharomyces yeasts in mixed cultures with Saccharomyces cerevisiae[J]. Archives of Microbiology, 2003, 180(4)∶ 257-263.
[10] COMITINI F, GOBBI M, DOMIZIO P, et al. Selected non-Saccharomyces wine yeasts in controlled multi starter fermentations with Saccharomyces cerevisiae[J]. Food Microbiology, 2011, 28(5)∶873-882.
[11] LEE P R, CHONG I S M, YU B, et al. Effects of sequentially inoculated Williopsis saturnus and Saccharomyces cerevisiae on volatile profiles of papaya wine[J]. Food Research International, 2012, 45(1)∶ 177-183.
[12] JEMEC K P, CADEZ N, ZAGORC T, et al. Yeast population dynamics in five spontaneous fermentations of Malvasia must[J]. Food Microbiology, 2001, 18(3)∶ 247-259.
[13] ANFANG N, BRAJKOVICH M, GODDARD M R. Co-fermentation with Pichia kluyveri increases varietal thiol concentrations in Savignon Blanc[J]. Australian Journal of Grape and Wine Research, 2009, 15(1)∶1-8.
[14] SUDHEER KUMAR Y, PRAKASAM R S, REDDY O V S. Optimisation of fermentation conditions for mango (Mangifera indica L.) wine production by employing response surface methodology[J]. International Journal of Food Science & Technology, 2009, 44(11)∶2320-2327.
[15] SODEN A, FRANCIS I L, OAKEY H, et al. Effects of cofermentation with Candida stellata and Saccharomyces cerevisiae on the aroma and composition of Chardonnay wine[J]. Australian Journal of Grape and Wine Research, 2000, 6(1)∶ 21-30.
[16] TORO M E, VAZQUEZ F. Fermentation behaviour of controlled mixed and sequential cultures of Candida cantarellii and Saccharomyces cerevisiae wine yeast[J]. World Journal of Microbiology and Biotechnology, 2002, 18(4)∶ 351-358.
[17] 楊雅利, 沈海亮, 闞建全. 紫色甘薯酒香氣成分分析和發酵規律[J].食品科學, 2012, 33(12)∶ 242-246.
[18] 呂慧威, 孫玉梅, 盧明春, 等. 自選酵母菌株草莓酒發酵特性比較[J].食品科學, 2010, 31(11)∶ 197-201.
[19] 張航. 電子舌傳感器快速檢測油茶籽油中摻雜棕櫚油[J]. 食品科學, 2013, 34(14)∶ 218-222. doi∶ 10.7506/spkx1002-6630-201314044.
[20] SUMBY K M, GRBIN P R, JIRANEK V. Microbial modulation of aromatic esters in wine∶ current kmowledge and future prospects[J]. South African Journal of Enology and Viticulture, 2010, 21∶ 97-129.
[21] MAURIZIO C, FRANCESCA C, MARIA M I, et al. Controlled mixed culture fermentation∶ a new perspective on the use of non-Saccharomyces yeasts in winemaking[J]. FEMS Yeast Research, 2010, 10(2)∶ 123-133.
[22] LEE P R, CHONG S M. Effect of precursors on volatile compoun ds in papaya wine fermented by mixed yeasts[J]. Food Technology and Biotechnology, 2013, 51(1)∶ 92-100.
[23] SAERENS S M G, DELVAUX F, VERSTREPEN K J, et al. Parameters affecting ethyl ester production by Saccharomyces cerevisiae during fermentation[J]. Applied and Environmental Microbiology, 2008, 74(2)∶ 454-461.
[24] FUKUDA K, YAMAMOTO N, KIYOKAWA Y, et al. Balance of activities of alcohol acetyltransferase and esterase in Saccharomyces cerevisiae is important for production of isoamyl acetate[J]. Applied and Environmental Microbiology, 1998, 64(10)∶4076-4153.
Effects of Successive Inoculation of Non-Saccharomyces Yeast on Aroma Components of Pineapple Wine by Mixed-Culture Fermentation
JIA Yanyan, LIU Sixin, LI Zhuoting, WANG Yanmei, YI Jinhao, DING Junquan, LI Congfa*
(College of Food Science and Technology, Hainan University, Haikou 570228, China)
Pineapple wines fermented with successive inoculation ofMetschnikowia agavesP3-3 andSaccharomyces cerevisiaeD254 were analyzed by gas chromatography-mass spectrometry (GC-MS) combined with head-space solid-phase microextraction (HS-SPME). The results showed that 53, 44, 41 and 20 aroma components were detected from group SI-I (inoculating D254 after 7 days of fermentation withP3-3), group CI (simultaneous inoculation of both P3-3 andD254), group SI-II (inoculating of P3-3 after 2 days of fermentation with D254) and the pineapple juice, respectiv ely. SI-I group dominated by P3-3 produced higher amounts of acetate ester (15.594%) than group CI (13.042%) and SI-II (6.655%). SI-I retained the highest concentration of the characteristic aroma components of pineapple including 3-(methylthio)propanoic acid methyl ester,3-(methylthio)propanoic acid ethyl ester and butyrolactone as compared to group CI (0.844%) and SI-II (0.819%). This study suggested that more characteristic aroma components of pineapple were retained and more ethyl esters were synthesized to increase the complexity of the wine in group SI-I. At last, with the extended survival ofP3-3, the highest sensory score was obtained from group SI-I. The successive inoculation of non-SaccharomycesandSaccharomycesyeasts may be a valuable method to manipulate yeast succession and to modulate the volatile profiles and organoleptic properties of pineapple wine.
non-Saccharomycesyeast;Metschnikowia agaves; mixed-culture fermentation;pineapple wine; aroma-active components
TS262.7
A
10.7506/spkx1002-6630-201517029
2015-05-03
“十一五”國家科技支撐計劃項目(200 7BAD76B01-2);海口市熱帶 農產品深加工重點實驗室項目(2013-45)
賈言言(1988—),女,碩士研究生,研究方向為應用微生物技術。E-mail:952262544@qq.co m
*通信作者:李從發(1967—),男,教授,博士,研究方向為食品生物技術。E-mail:congfa@vip.163.com
