PEF作用下亞致死損傷釀酒酵母修復條件的初步研究
2014-12-16 08:09:38楊瑞金顧艷潔
食品工業科技 2014年21期
關鍵詞:體系
陳 晨,趙 偉,楊瑞金,顧艷潔
(1.江南大學食品學院,江蘇無錫214122;2.江南大學食品科學與技術國家重點實驗室,江蘇無錫214122)
高壓脈沖電場(PEF)技術能夠有效地殺滅食品中腐敗菌和致病菌,同時能夠較好保持食品固有的營養成分[1]。國內外的學者在研究大量PEF不同體系的殺菌效果同時,已開始對PEF的殺菌機理展開進一步的研究。有研究表明,PEF等非熱殺菌方式除殺滅部分微生物外,還會形成一定比例的損傷亞致死細胞,亞致死微生物在特定的環境下可以進行自我修復[2-4],恢復其自身繁殖能力,這使得直接對食品安全和貨架期造成不良的影響。因此,研究亞致死微生物的修復條件,從而進一步滅活微生物和驗證PEF殺菌食品的安全性是非常必要。陳曉嬋等人[5]對比研究了高壓脈沖電場對草莓汁中大腸桿菌和釀酒酵母損傷亞致死細胞產生的影響,同時還探究發現了PEF處理后冷臧或溫和熱處理都可以抑制亞致死釀酒酵母修復或使其進一步死亡。Somolinos[6]等選用不同 pH培養基研究了 PEF處理下下亞損傷釀酒酵母細胞的自我修復情況。Pernic[7]等人以大腸桿菌和沙門氏菌作為目標微生物,提出PEF亞致死損傷是一個累積的過程。
釀酒酵母是液體食品(乳制品、果蔬汁、非酒精性飲料等)中的常見污染菌種,對各種防腐劑、電離輻射照射、冷凍等的抵抗性強[8]。草莓營養豐富,常規熱殺菌會造成熱敏性活性成分流失,降低了草莓汁的營養價值,因此國內外學者開始傾向于利用PEF等非熱加工技術對草莓汁等果蔬飲品進行殺菌研究,以便于應用于實際生產。在釀酒酵母生長過程中,除需碳源、氮源和生物素外,還需要K+、Ca2+、Mg2+等無機金屬離子作為生長因子,培養環境中金屬離子的含量影響著釀酒酵母的生理代謝活動[9-10]。
本實驗以釀酒酵母作為研究模型,以培養體系和金屬離子作為研究平臺,選用模擬體系和實際體系(草莓汁體系),初步研究了培養體系和金屬離子對PEF作用下對亞致死損傷釀酒酵母的影響。為PEF殺菌機理提供進一步的研究提供了理論依據。
1 材料與方法
1.1 材料與儀器
釀酒酵母菌 江南大學食品酶學實驗室提供,菌種編號C-03;酵母浸出粉LP0021 英國OXOID公司;蛋白胨F403、葡萄糖、氯化鈉、無水氯化鈣、硫酸鎂、磷酸氫二鉀 均為分析純,購自國藥集團化學試劑有限公司。
OSU-4L型實驗室規模PEF連續處理設備 美國俄亥俄州立大學;DRP-9082型電熱恒溫培養箱 上海森信實驗儀器有限公司;LDZX-50KB立式壓力蒸汽滅菌器 上海申安醫療器械廠;XSW-CJ-2A標準型凈化工作臺 吳江市綠葉空調凈化有限公司;QYC2102-C型恒溫培養搖床 上海新苗醫療器械有限公司。
1.2 實驗方法
1.2.1 微生物的活化培養與接種 釀酒酵母(C-03)由酵母浸出粉胨葡萄糖(YPD)瓊脂斜面培養基移至YPD液體培養基,30℃搖床200r/min培養16~20h至對數生長期。菌液在4℃條件下冷凍離心(8000r/min,6min)得到菌液沉淀,用pH7.2滅菌的磷酸鹽緩沖液(PBS)反復洗2次后重懸,菌懸液濃度達到107~108CFU/mL,電導率約為2000μs/cm。
1.2.2 模擬體系和實際體系配制 模擬體系一:主要是以酵母浸出粉、蛋白胨 F403和葡萄糖配制0.1%酵母浸出粉緩沖溶液、0.1%蛋白胨緩沖溶液、0.1%葡萄糖緩沖溶液作為目標體系,可利用PBS調節電導率至3000μs/cm左右。模擬體系二:在以pH7.2滅菌的PBS菌懸液中,用無水氯化鈣、硫酸鎂、磷酸氫二鉀在緩沖體系添加不同濃度的金屬離子。實際體系:選用新鮮草莓汁,將市售新鮮草莓洗凈后,按料液比1∶6,用果汁機榨汁,用離心機離心(3000r/min,15min)獲取上清液,抽濾后草莓汁電導率約為3000μs/cm,用無水氯化鈣、硫酸鎂、磷酸氫二鉀在草莓汁中添加不同濃度的金屬離子。以上均將10mL培養液1.2.1接種至90mL不同處理體系中。
1.2.3 高壓脈沖電場殺菌 采用實驗室規模連續PEF處理設備,脈沖電場為雙極方形波脈沖電場。實驗室選擇6個連續處理腔,選用了電場強度20kV/cm,處理循環時間為400μs,脈沖寬度 2μs,脈沖頻率200Hz,循環式冷卻水浴的溫度為15℃。PEF殺菌處理后的不同樣品在室溫下放置 0、5、30、50、70、120 和150min進行修復研究。
1.2.4 選擇性培養基和非選擇性培養基 采用選擇性平板法測定亞致死損傷微生物,計數參照Perni等的方法[7]。受損失的酵母細胞在正常培養基上可以進行生長,但是在高滲透環境下無法維持滲透平衡,故不能生長;而未受損傷的酵母細胞可以在適度的高滲透環境下生長。故選用兩種培養基進行亞致死損傷微生物的測定:一種是正常的非選擇性培養基,另一種是選擇性高滲透培養基,以顧艷潔等人[11]確定的4%NaCl濃度為釀酒酵母細胞的臨界滲透壓,即常規培養基補加4%NaCl。
1.2.5 微生物計數 根據GB/T4789.2-2003方法,對PEF處理前后的菌液采用菌落平板計數法進行微生物計數。通過將菌液用0.85%生理鹽水以10倍稀釋法進行逐級稀釋,選擇合適的稀釋度,吸取1.0mL稀釋樣于滅菌平皿中與培養基混合搖勻,于(30±0.1)℃條件下培養24h并計數。
1.2.6 cFDA標記 羧基熒光素乙酰乙酸(cFDA)是一種親脂性、無熒光的物質,能夠自由穿越細胞膜,可用來評價細胞內酶活力。cFDA進入細胞后會被細胞內非特異性酯酶水解生成極性、不能穿過細胞膜的熒光物質—羧基熒光素(cF)。cF標記細胞經488nm激發后可在530nm發射綠色熒光,熒光越強表明細胞非特異性內酯酶活越強。取PEF處理前后的釀酒酵母菌懸液1.0mL用cFDA染液(終濃度為5μg/mL)37℃避光染色30min,PBS反復沖洗兩次去除多余的染料。
1.2.7 釀酒酵母非特異性內酯酶活力的測定 經cFDA染色后的酵母細胞用流式細胞分析儀檢測FSC、SSC及FL1熒光通道(綠色)的各項指標。每個樣品檢測20000個細胞。采用488nm激發光,cFDA標記細胞在530nm處發綠色熒光,通過FL1通道檢測,數據采集后用CellQuest Pro軟件進行分析。
2 結果與討論
2.1 不同緩沖體系中亞致死損傷釀酒酵母的修復
參考 Garc?á,Somolinos 等人[6]對 PEF 處理后大腸桿菌及釀酒酵母修復情況考察時的培養液選取原則,選取相同電導率的0.1%酵母浸出粉緩沖溶液、0.1%蛋白胨緩沖溶液、0.1%葡萄糖緩沖溶液作為釀酒酵母的培養修復環境,采用選擇性培養基與非選擇性培養基進行菌落計數,結果如表1~表3所示。
通過相同條件PEF處理后,三種緩沖體系中釀酒酵母的存活情況及亞致死損傷釀酒酵母的修復均存在差異。在0.1%酵母浸出粉緩沖體系中,PEF處理后亞損傷酵母細胞約為2個對數,在0.1%蛋白胨緩沖體系中,PEF處理后亞損傷酵母細胞約為1個對數,而在0.1%葡萄糖緩沖體系中,PEF處理后亞損傷酵母細胞約為0.8個對數。亞致死損傷酵母的修復速度在成分相對豐富的酵母浸出粉緩沖體系與蛋白胨緩沖體系中遠快于成分較為單一的葡萄糖緩沖體系,在酵母浸出粉緩沖體系與蛋白胨緩沖液中處理后30min內損傷細胞即基本修復完畢,在葡萄糖緩沖液中處理后120min左右損傷細胞才修復完全。

表1 0.1%酵母浸出粉緩沖體系中PEF處理后釀酒酵母的修復Table 1 The recovery of PEF treated S.cerevisiae in 0.1%Yeast extract solution

表2 0.1%蛋白胨緩沖體系中PEF處理后釀酒酵母的修復Table 2 The recovery of PEF treated S.cerevisiae in 0.1%Peptone solution

圖1 不同添加量的金屬離子Ca2+對釀酒酵母存活量的影響Fig.1 Effect of different addition of metal ions Ca2+on the survival of S.cerevisiae cells
2.2 金屬離子對模擬體系中亞致死損傷釀酒酵母的修復
無機鹽離子是釀酒酵母生長過程中必不可少的營養物質,其中 Na、K、Ca、Mg等金屬陽離子對釀酒酵母的正常生理機能有著重要的影響。本實驗初步選取K、Ca、Mg三種均對酶有一定激活作用的金屬離子考察其對PEF處理后亞致死釀酒酵母細胞的修復作用。
參照釀酒酵母生長過程中三種離子的最適添加量以及蘋果、草莓等常見水果中三種金屬離子的一般含量,分別向釀酒酵母緩沖溶液中添加不同濃度的金屬離子,同時保證微生物初始濃度在一個對數級。PEF處理前后進行菌落計數,得到釀酒酵母的存活情況如圖1~圖3所示。
在未添加特定金屬離子的對照組,PEF處理后釀酒酵母存活對數約為2.3~3.0個對數,存活細胞中亞致死損傷釀酒酵母的所占比例約為40%左右,PEF處理后亞致死酵母細胞存活對數約為1個對數,證明PEF處理后有亞致死酵母細胞的存在。
從圖1可見,當Ca2+濃度升至50mg/L后,對酵母細胞存活有顯著影響(p<0.05),可見Ca2+存在會影響酵母的存活。而當Ca2+濃度升至200mg/L,非選擇性培養基和選擇性培養基分別檢測酵母存活對數為4.31和4.36個對數,說明修復完全。從圖2可見,當Mg2+濃度升至30mg/L后,酵母存活對數顯著提高至約4.7個對數,而隨著離子濃度的增加,存活不同大寫或小寫字母代表不同培養基的差異性(p<0.05)。
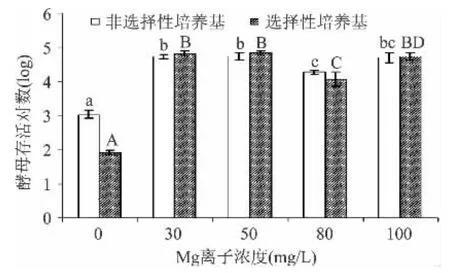
圖2 不同添加量的金屬離子Mg2+對釀酒酵母存活量的影響Fig.2 Effect of different addition of metal ions Mg2+on the survival of S.cerevisiae cells
不同大寫或小寫字母代表不同培養基的差異性(p<0.05)。對數略有所下降,當離子濃度到達100mg/L,存活對不同大寫或小寫字母代表不同培養基的差異性(p<0.05)。數變化不顯著(p>0.05)。說明在 Mg2+濃度升至30mg/L后,已修復完全。從圖3可見,當K+濃度升至0.8g/L后,酵母存活對數顯著提高至4.4個對數,已修復完全。但隨著離子濃度的提高,會對酵母菌的存活產生一定的副作用。

圖3 不同添加量的金屬離子K+對釀酒酵母存活量的影響Fig.3 Effect of different addition of metal ions K+on the survival of S.cerevisiae cells
考慮到K、Ca、Mg三種金屬離子均對酶有一定激活作用,初步以釀酒酵母內酯酶為研究對象,應用cFDA對酵母進行熒光標記能夠反應酵母的酯酶和蛋白的水平,從而反應酵母的活性。
選取約20000個細胞,以cFDA標記釀酒酵母細胞內酯酶,FAM使光越強表明細胞非特異性內酯酶活越強,如圖4所示,三種金屬離子的添加對釀酒酵母的非特異性內酯酶活均產生了不同程度的影響。Ca2+的添加使得釀酒酵母細胞內酯酶活呈現先降后升的趨勢;Mg2+的添加使得釀酒酵母細胞內酯酶活呈現負增長的趨勢;而K+的添加使得釀酒酵母細胞內酯酶活呈現增加的趨勢。說明三種金屬離子添加對亞致死損傷釀酒酵母細胞的修復也部分體現在于對非特異性內酯酶的修復。金屬離子的添加對PEF處理下酵母細胞內酯酶的修復是個較為復雜的過程,目前研究未能闡述其影響機理。
2.3 金屬離子對食品體系中亞致死損傷釀酒酵母的修復
為了更全面的反映三種金屬離子對亞致死損傷酵母細胞的修復作用,選取鮮榨草莓汁作為實際食品體系,三種離子的添加量與緩沖體系中保持一致,進行菌落計數,得到釀酒酵母的存活情況如圖5~圖7所示。
從圖5可見,在未添加特定金屬離子的對照組中,PEF處理后產生亞致死酵母細胞約為0.5個對數。金屬陽離子的添加對鮮榨草莓汁中亞致死損傷酵母細胞的修復產生了一定的促進效果。當Ca2+濃度升至50mg/L后,Ca2+添加對酵母細胞存活有顯著影響(p<0.05),酵母存活對數約達至3個對數,且修復完全。Ca2+主要以離子狀態存在于細胞中以控制細胞的生理狀態,如調節膜的透性、激活酶等。馮玉娟等[12]曾指出釀酒酵母的生長代謝速度隨著Ca2+的添加量而加快,而當Ca2+濃度超過一定量之后釀酒酵母生長會受到抑制。Yamada等人在對 Ca2+-ATPase活力的測定中已經得知PEF處理會顯著增加釀酒酵母中Ca2+-ATPase的活力,因此可以推測當緩沖體系中額外添加Ca2+后,釀酒酵母依賴于高爾基體上活躍的Ca2+-ATPase將胞外Ca2+運輸到胞內,從而加快損傷釀酒酵母的修復速度[13]。
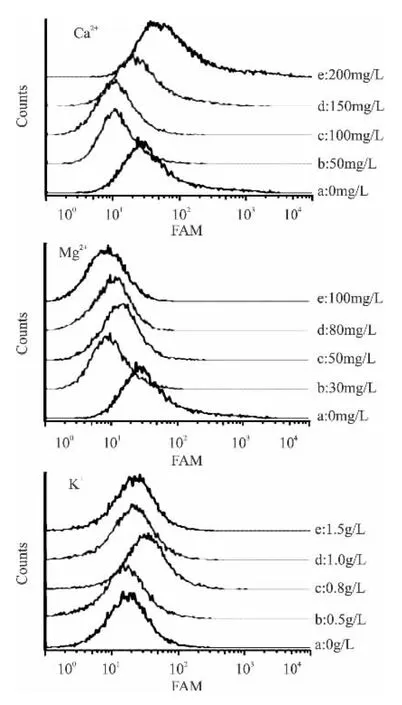
圖4 不同添加量的金屬離子對釀酒酵母內酯酶活的影響Fig.4 Effect of different addition of metal ions on the lactone enzyme activity of S.cerevisiae cells

圖5 不同添加量的金屬離子Ca2+對草莓汁中釀酒酵母存活量的影響Fig.5 Effect of different addition of metal ions Ca2+on the sublethally injured percentage of S.cerevisiae in Strawberry juice
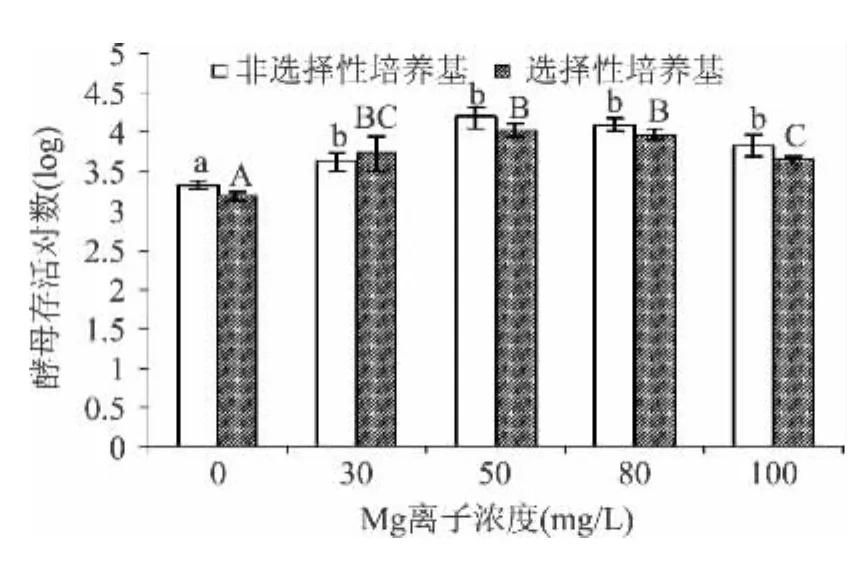
圖6 不同添加量的金屬離子Mg2+對草莓汁中釀酒酵母存活量的影響Fig.6 Effect of different addition of metal ions Mg2+on the sublethally injured percentage of S.cerevisiae in Strawberry juice

圖7 不同添加量的金屬離子K+對草莓汁中釀酒酵母存活量的影響Fig.7 Effect of different addition of metal ions K+on the sublethally injured percentage of S.cerevisiae in Strawberry juice
從圖6可見,當Mg2+濃度升至30mg/L,Mg2+的添加對酵母存活對數影響顯著(p<0.05),當Mg2+濃度升至50mg/L和80mg/L,Mg2+濃度的添加對酵母存活數影響不顯著(p<0.05),隨著濃度提高,會對酵母菌的存活產生一定的副作用,表現在釀酒酵母存活對數的下降。
從圖7可見,在未添加特定金屬離子的對照組中,PEF處理后亞致死酵母細胞約為0.8個對數,當K+濃度升至0.8g/L后,K+的添加對酵母細胞存活有顯著影響(p<0.05),酵母存活對數約達至4個對數。隨著濃度提高,酵母存活對數下降,殺菌效果反而提高。可能是由于釀酒酵母在受到PEF處理刺激時會導致細胞膜通透性、滲透壓及膜電位改變,從而可能引起K+外流。當緩沖體系中額外加入K+后,胞外K+滲透壓高于胞內,阻止了酵母細胞內K+外流現象,同時刺激了酵母內酯酶活,促進亞致死損傷釀酒酵母細胞的修復。
3 結論
在單一的葡萄糖緩沖體系存在的亞損傷酵母細胞較復雜成分的酵母浸出粉、蛋白胨溶液少。亞致死酵母細胞在以YPD培養基為主要成分的酵母浸出粉、蛋白胨和葡萄糖緩沖溶液于30~70min內完成修復。
在模擬體系中,未添加特定金屬離子的對照組均存在亞損傷酵母細胞。當Ca2+濃度升至50mg/L,Mg2+濃度升至30mg/L,當K+濃度升至0.8g/L后,亞致死損傷細胞均已修復。
在真實草莓汁體系中,當Ca2+濃度升至50mg/L,K+濃度升至0.8g/L后,亞致死損傷細胞均已修復。Mg2+離子的添加對PEF作用下亞致死酵母修復影響不顯著,但隨著離子濃度的增加,PEF對酵母細胞殺菌效果先降后升。
[1]曾新安,陳勇.脈沖電場非熱滅菌技術[M].北京:中國輕工業出版社,2004,3-7.
[2]Jaeger H,Schulz A,Karapettov N,et al.Protective effect of milk constituents and sublethal injuries limiting process effectiveness during PEF inactivation of Lb.rhamnosus[J].International Journal of Food Microbiology,2009,134:154-161.
[3]Simpson R K,Whittington R,Earnshaw R G,et al.Pulsed high electric field causes‘all or nothing’membrane damage in Listeria monocytogenes and Salmonella typhimurium,but membrane H+-ATPase is not a primary target[J].International Journal of Food Microbiology,1999,48:1-10.
[4]Russell N J,Colley M ,Simpson R K ,et al.Mechanism of action of pulsed high electric field(PHEF)on the membranes of food-poisoning bacteria is an ‘all-or-nothing’effect[J].International Journal of Food Microbiology.2000,55:133-136.
[5]陳曉嬋,趙偉,楊瑞金,等.高壓脈沖電場處理草莓汁中損傷亞致死大腸桿菌與釀酒酵母的檢測[J].食品工業科技,2013,34(16):79-83.
[6]Somolinos M ,Ma?as P ,Condón S,et al.Recovery of Saccharomyces cerevisiae sublethally injured cells after Pulsed Electric Fields[J].International Journal of Food Microbiology.2008,125:352-356.
[7]Perni S,Chalise P R,Shama G ,et al.Bacterial cells exposed to nanosecond pulsed electric fields show lethal and sublethal effects[J].International Journal of Food Microbiology,2007,120:311-314.
[8]周剛.酵母對食品的污染[J].環境衛生學雜志,1979,6:325-329.
[9]Pironcheva G L.The effect of magnesium ions during beer fermentation[J].Cytobios,1998,94:135-139.
[10]McConkey D J,Orrenius S.Breakthroughs and views:The role of calcium in the regulation of apoptosis[J].Biochemical and Biophysical Research Communications,1997,239:357-366.
[11]顧艷潔,趙偉,楊瑞金,等.高壓脈沖電場作用下亞致死酵母的存在與檢測[J].食品發酵與工業,2012,38(7):90-93.
[12]馮玉娟.釀酒酵母生長代謝影響因素的測定[J].濰坊學院學報,2010,10(6):110-112.
[13]Akiko Yamada,Takenori Yamamoto,Yuya Yoshimura,et al.Ca2+-induced permeability transition can be observed even in yeast mitochondria under optimized experimental conditions[J].Biochimica et Biophysica Acta,2009,1787:1486-1491.
猜你喜歡
商品與質量(2021年43期)2022-01-18 05:31:22
杭州(2020年23期)2021-01-11 00:54:42
新世紀智能(數學備考)(2020年11期)2021-01-04 00:38:16
中國外匯(2019年17期)2019-11-16 09:31:14
中國衛生(2015年12期)2015-11-10 05:13:40
現代企業(2015年1期)2015-02-28 18:43:18
汽車零部件(2014年5期)2014-11-11 12:24:28
新高考·高一物理(2014年1期)2014-09-18 01:26:07
浙江人大(2014年1期)2014-03-20 16:19:53
終身教育研究(2012年4期)2012-03-25 10:41:11
